
Prohibitin 1 Regulates Inflammatory Mediators and Reactive Oxygen Species in Retinal Endothelial Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
3. Results
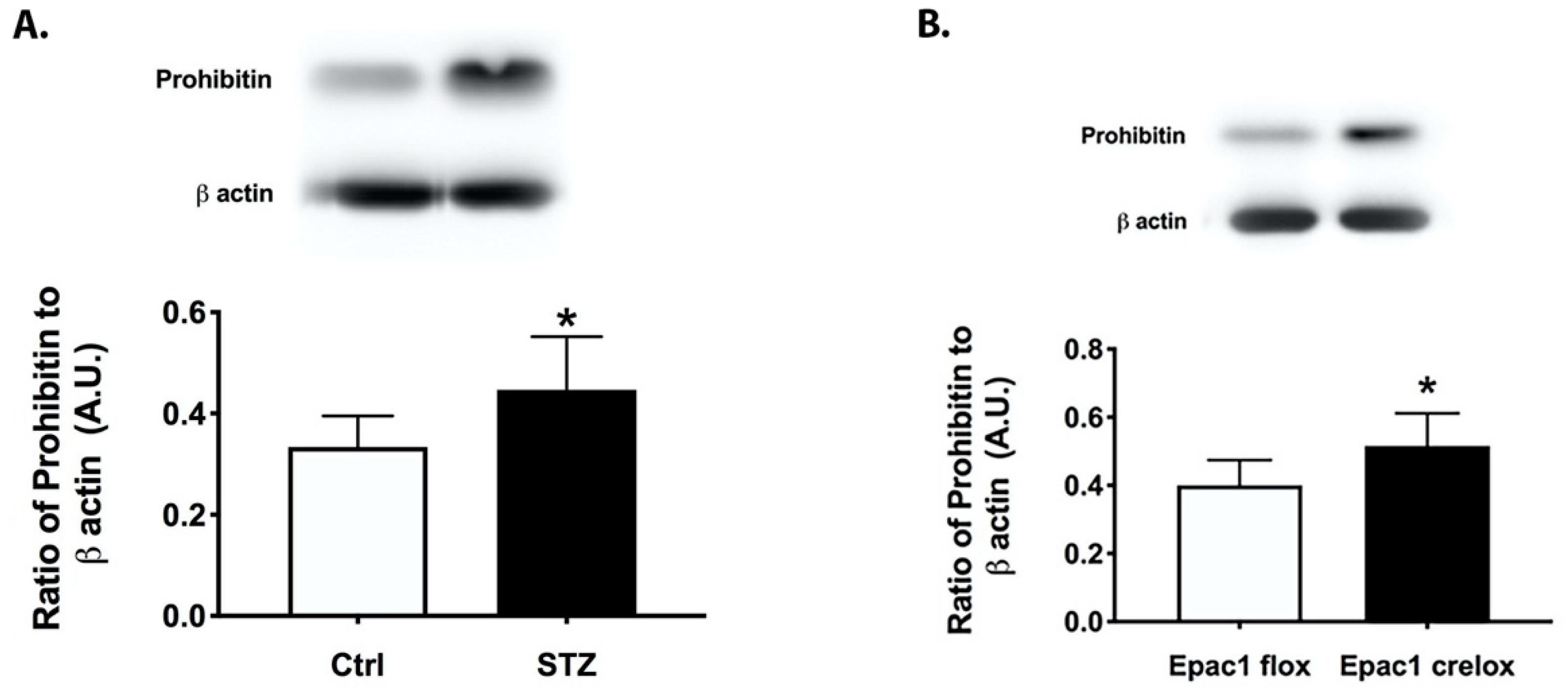
3.1. Prohibitin 1 Is Increased in the Diabetic Retina and in Retinal Lysates from cdh5Cre-Epac1 Mice
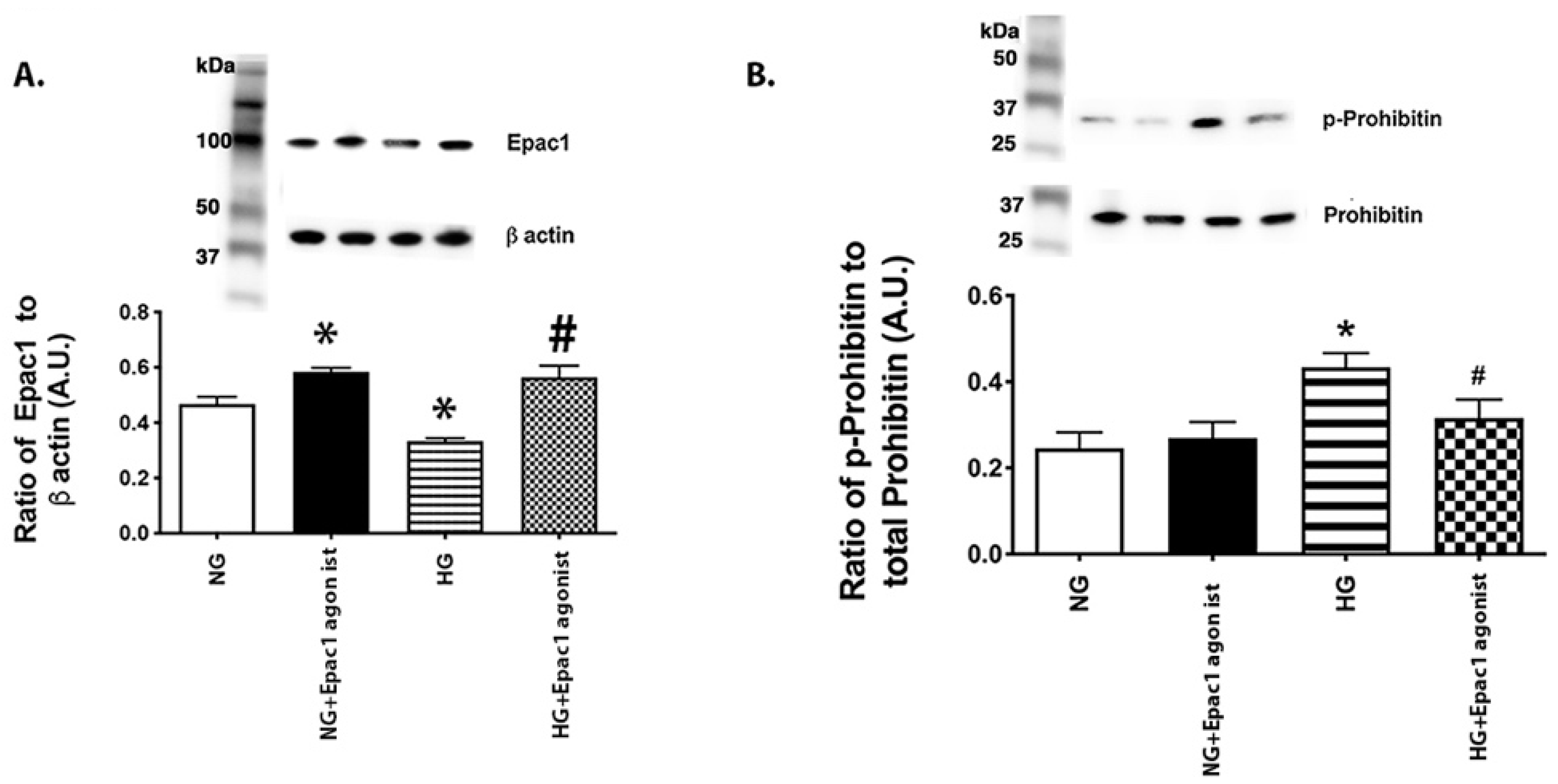
3.2. An Epac1 Agonist Can Decrease Phosphorylation of Prohibitin 1 in REC Grown in High Glucose
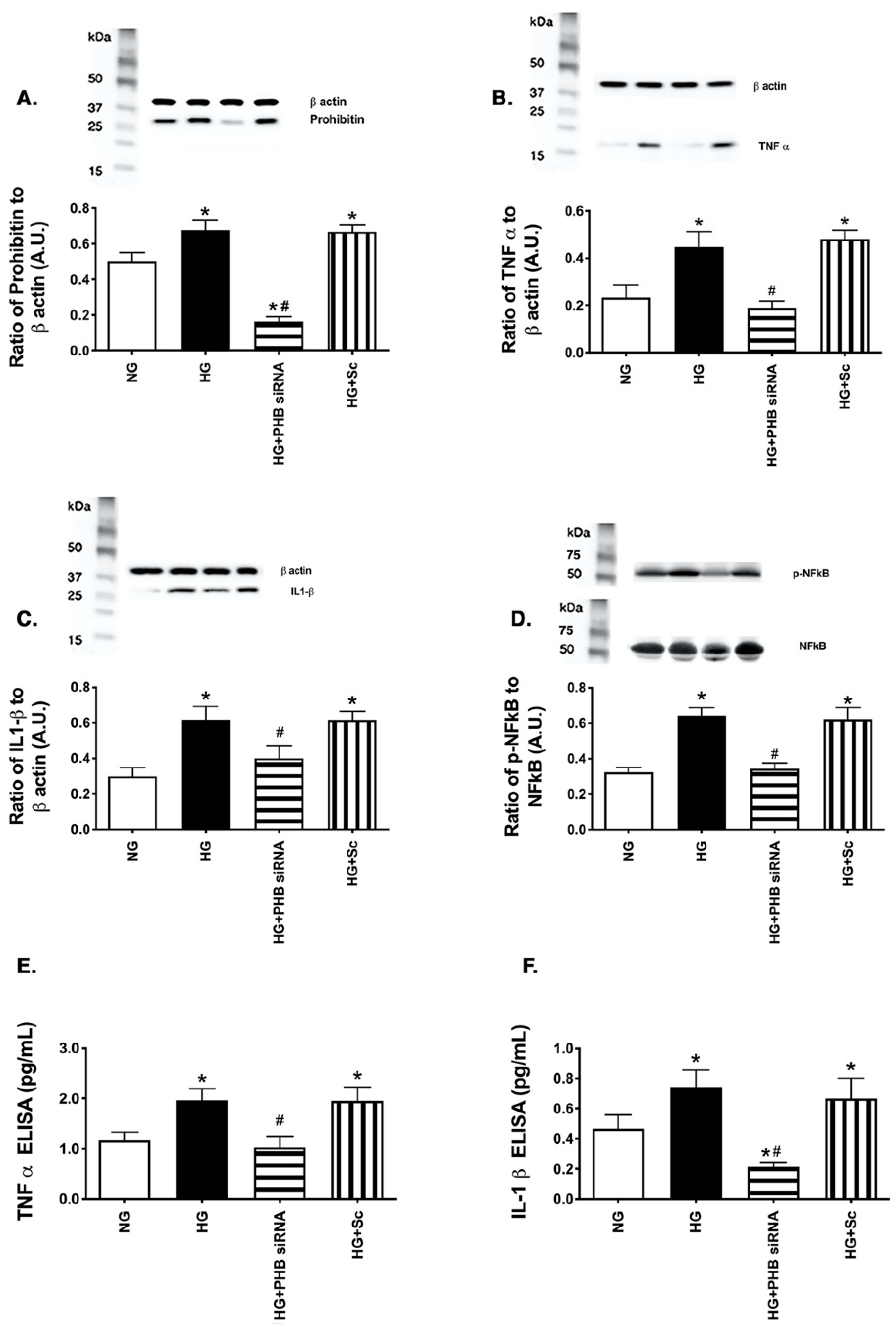
3.3. Reduction in Prohibitin 1 Levels Inhibited Inflammatory Mediator Levels in REC Grow in High Glucose
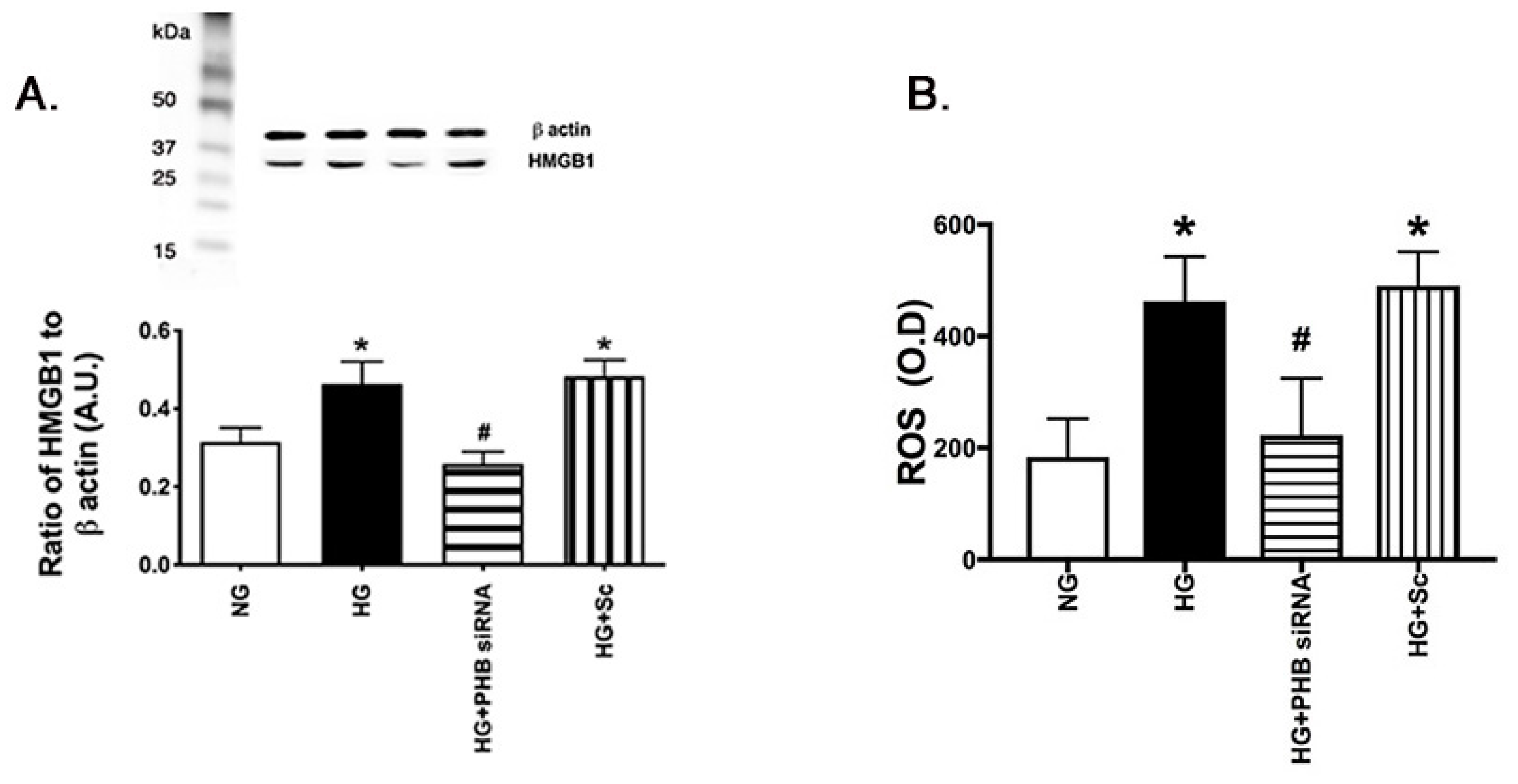
3.4. Reduced Prohibitin 1 Decreased HMGB1 and ROS Levels in the REC Grown in High Glucose
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Ko, K.S.; Tomasi, M.L.; Iglesias-Ara, A.; French, B.A.; French, S.W.; Ramani, K.; Lozano, J.J.; Oh, P.; He, L.; Stiles, B.L.; et al. Liver-specific deletion of prohibitin 1 results in spontaneous liver injury, fibrosis, and hepatocellular carcinoma in mice. Hepatology 2010, 52, 2096–2108. [Google Scholar] [CrossRef]
- Schleicher, M.; Shepherd, B.R.; Suarez, Y.; Fernandez-Hernando, C.; Yu, J.; Pan, Y.; Acevedo, L.M.; Shadel, G.S.; Sessa, W.C. Prohibitin-1 maintains the angiogenic capacity of endothelial cells by regulating mitochondrial function and senescence. J. Cell Biol. 2008, 180, 101–112. [Google Scholar] [CrossRef]
- Xu, Y.; Ye, C.; Tang, L.; Pan, B.; Li, Y. Decreased expression of prohibitin 1 mediates high-mobility group protein 1-induced pulmonary artery endothelial cell injury. Zhong Nan Da Xue Xue Bao Yi Xue Ban 2020, 45, 1024–1034. [Google Scholar] [PubMed]
- Guyot, A.C.; Leuxe, C.; Disdier, C.; Oumata, N.; Costa, N.; Roux, G.L.; Varela, P.F.; Duchon, A.; Charbonnier, J.B.; Herault, Y.; et al. A Small Compound Targeting Prohibitin with Potential Interest for Cognitive Deficit Rescue in Aging mice and Tau Pathology Treatment. Sci. Rep. 2020, 10, 1143. [Google Scholar] [CrossRef] [PubMed]
- Ande, S.R.; Xu, Y.X.Z.; Mishra, S. Prohibitin: A potential therapeutic target in tyrosine kinase signaling. Signal Transduct. Target. Ther. 2017, 2, 17059. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Ande, S.R.; Nyomba, B.L. The role of prohibitin in cell signaling. FEBS J. 2010, 277, 3937–3946. [Google Scholar] [CrossRef] [PubMed]
- Sripathi, S.R.; Sylvester, O.; He, W.; Moser, T.; Um, J.Y.; Lamoke, F.; Ramakrishna, W.; Bernstein, P.S.; Bartoli, M.; Jahng, W.J. Prohibitin as the Molecular Binding Switch in the Retinal Pigment Epithelium. Protein J. 2016, 35, 1–16. [Google Scholar] [CrossRef]
- Patrick, A.T.; He, W.; Madu, J.; Sripathi, S.R.; Choi, S.; Lee, K.; Samson, F.P.; Powell, F.L.; Bartoli, M.; Jee, D.; et al. Mechanistic dissection of diabetic retinopathy using the protein-metabolite interactome. J. Diabetes Metab. Disord. 2020, 19, 829–848. [Google Scholar] [CrossRef]
- Lee, H.; Arnouk, H.; Sripathi, S.; Chen, P.; Zhang, R.; Bartoli, M.; Hunt, R.C.; Hrushesky, W.J.; Chung, H.; Lee, S.H.; et al. Prohibitin as an oxidative stress biomarker in the eye. Int. J. Biol. Macromol. 2010, 47, 685–690. [Google Scholar] [CrossRef]
- Liu, L.; Jiang, Y.; Steinle, J.J. Epac1 and Glycyrrhizin Both Inhibit HMGB1 Levels to Reduce Diabetes-Induced Neuronal and Vascular Damage in the Mouse Retina. J. Clin. Med. 2019, 8, 772. [Google Scholar] [CrossRef]
- Liu, L.; Jiang, Y.; Steinle, J. Epac1 regulates TLR4 signaling in the diabetic retinal vasculature. Cytokine 2021, 144, 155576. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Jiang, Y.; Chahine, A.; Curtiss, E.; Steinle, J.J. Epac1 agonist decreased inflammatory proteins in retinal endothelial cells, and loss of Epac1 increased inflammatory proteins in the retinal vasculature of mice. Mol. Vis. 2017, 23, 1–7. [Google Scholar] [PubMed]
- Jiang, Y.; Steinle, J.J. Epac1 Requires AMPK Phosphorylation to Regulate HMGB1 in the Retinal Vasculature. Investig. Ophthalmol. Vis. Sci. 2020, 61, 33. [Google Scholar] [CrossRef]
- Wang, Y.-J.; Guo, X.-L.; Li, S.-A.; Zhao, Y.-Q.; Liu, Z.-C.; Lee, W.-H.; Xiang, Y.; Zhang, Y. Prohibitin is involved in the activated internalization and degradation of protease-activated receptor 1. Biochim. Biophys. Acta 2014, 1843, 1393–1401. [Google Scholar] [CrossRef]
- Liu, L.; Jiang, Y.; Steinle, J.J. Toll-Like Receptor 4 Reduces Occludin and Zonula Occludens 1 to Increase Retinal Permeability Both in vitro and in vivo. J. Vasc. Res. 2017, 54, 367–375. [Google Scholar] [CrossRef]
- Jiang, Y.; Liu, L.; Steinle, J.J. Epac1 deacetylates HMGB1 through increased IGFBP-3 and SIRT1 levels in the retinal vasculature. Mol. Vis. 2018, 24, 727–732. [Google Scholar]
- Barati, M.T.; Gould, J.C.; Salyer, S.A.; Isaacs, S.; Wilkey, D.W.; Merchant, M.L. Influence of Acute High Glucose on Protein Abundance Changes in Murine Glomerular Mesangial Cells. J. Diabetes Res. 2016, 2016, 3537863. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Park, J.; Ko, K.S. Lipopolysaccharide-induced innate immune responses are exacerbated by Prohibitin 1 deficiency and mitigated by S-adenosylmethionine in murine macrophages. PLoS ONE 2020, 15, e0241224. [Google Scholar] [CrossRef] [PubMed]
- Theiss, A.L.; Idell, R.D.; Srinivasan, S.; Klapproth, J.M.; Jones, D.P.; Merlin, D.; Sitaraman, S.V. Prohibitin protects against oxidative stress in intestinal epithelial cells. FASEB J. 2007, 21, 197–206. [Google Scholar] [CrossRef]
- Dong, W.-Q.; Chao, M.; Lu, Q.-H.; Chai, W.-L.; Zhang, W.; Chen, X.-Y.; Hong-Liang, T.; Wang, L.-B.; Tian, H.-L.; Chen, Y.-G.; et al. Prohibitin overexpression improves myocardial function in diabetic cardiomyopathy. Oncotarget 2015, 7, 66–80. [Google Scholar] [CrossRef]
- Theiss, A.L.; Jenkins, A.K.; Okoro, N.I.; Klapproth, J.-M.A.; Merlin, D.; Sitaraman, S.V. Prohibitin Inhibits Tumor Necrosis Factor alpha–induced Nuclear Factor-kappa B Nuclear Translocation via the Novel Mechanism of Decreasing Importin α3 Expression. Mol. Biol. Cell 2009, 20, 4412–4423. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mishra, S.; Nyomba, B.G. Prohibitin—At the crossroads of obesity-linked diabetes and cancer. Exp. Biol. Med. 2017, 242, 1170–1177. [Google Scholar] [CrossRef] [PubMed]
- Theiss, A.L.; Sitaraman, S.V. The role and therapeutic potential of prohibitin in disease. Biochim. Biophys. Acta 2011, 1813, 1137–1143. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, L.; Jiang, Y.; Steinle, J.J. Prohibitin 1 Regulates Inflammatory Mediators and Reactive Oxygen Species in Retinal Endothelial Cells. J. Clin. Med. 2022, 11, 1915. https://doi.org/10.3390/jcm11071915
Liu L, Jiang Y, Steinle JJ. Prohibitin 1 Regulates Inflammatory Mediators and Reactive Oxygen Species in Retinal Endothelial Cells. Journal of Clinical Medicine. 2022; 11(7):1915. https://doi.org/10.3390/jcm11071915
Chicago/Turabian StyleLiu, Li, Youde Jiang, and Jena J. Steinle. 2022. "Prohibitin 1 Regulates Inflammatory Mediators and Reactive Oxygen Species in Retinal Endothelial Cells" Journal of Clinical Medicine 11, no. 7: 1915. https://doi.org/10.3390/jcm11071915
APA StyleLiu, L., Jiang, Y., & Steinle, J. J. (2022). Prohibitin 1 Regulates Inflammatory Mediators and Reactive Oxygen Species in Retinal Endothelial Cells. Journal of Clinical Medicine, 11(7), 1915. https://doi.org/10.3390/jcm11071915