
Spermatic Microbiome Characteristics in Infertile Patients: Impact on Sperm Count, Mobility, and Morphology




Abstract
:1. Introduction
2. Methods
2.1. Selected Samples
2.2. Sample Preparation
2.2.1. Nucleic Acid Extraction
2.2.2. Library Preparation and Sequencing
2.3. Bioinformatic Analyses
2.3.1. Quality Filtering and Diversity Analysis
2.3.2. Relative Abundance Graphs
3. Results
3.1. Study Population
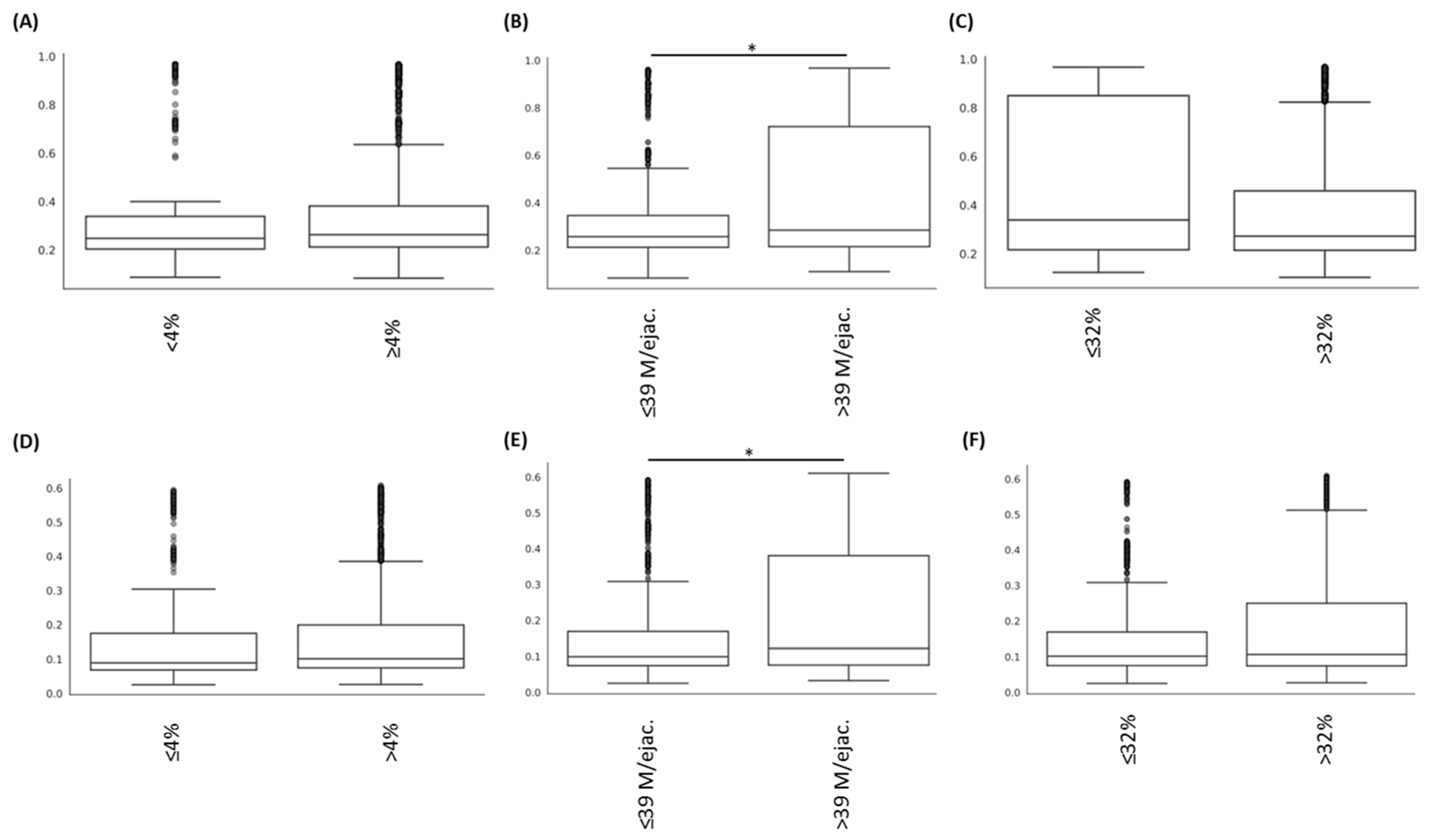
3.2. Alteration of the Sperm Microbiome Diversities Is Associated with Sperm Abnormalities

{kind=link}
{kind=link}
{kind=link}
| Clinical Characteristic | Motility | Sperm Count | Morphology | |||
|---|---|---|---|---|---|---|
| Motility < 32% (n = 29; 31.8%) | Motility ≥ 32% (n = 62; 68.2%) | Sperm Count ≤ 39 Millions per Ejaculate (n = 20; 22.0%) | Sperm Count > 39 Millions per Ejaculate (n = 71; 78%) | Normal Morphology < 4% (n = 38; 41.8%) | Normal Morphology ≥ 4% (n = 53; 58.2%) | |
| Age (in years) (med; [IQR]) | 32 [31–35] | 34.5 [30–37.8] | 33 [30.5–37.8] | 33 [30–37] | 33.5 [31–36.8] | 33 [30–38] |
| Duration of sexual abstinence in days (med; [IQR]) | 4 [3–5] | 4 [3–5.8] | 4 [2.8–5] | 4 [3–6] | 4 [3–5] | 4 [3–6] |
| Ejaculate volume (in mL) (med; [IQR]) | 3.2 [2.5–4.3] | 3.9 [2.8–4.7] | 3.4 [2.5–3.9] | 4 [2.9–4.8] | 3.3 [2.6–4.3] | 3.9 [2.8–4.7] |
| Tobacco consumption (ongoing/stopped/never smoked) | 7/5/15 | 21/14/19 | 5/3/8 | 23/16/26 | 13/5/15 | 15/14/19 |
| Spermoculture results (% of positive results) | 5 (17.2%) | 17 (27.4%) | 2 (10%) | 20 (28.2%) | 7 (18.4%) | 15 (28.3%) |
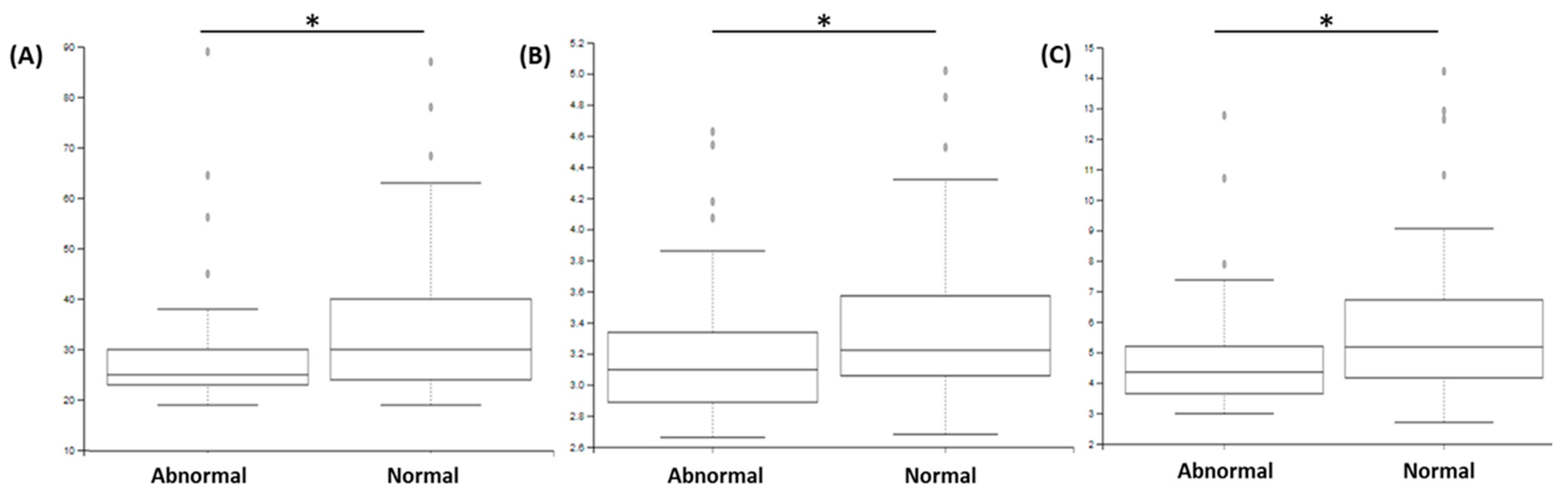
3.3. Sperm Abnormalities Are Associated with Modification in Specific Bacteria
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blondel, B.; Supernant, K. Enquête Nationale Périnatale 2003—Situation en 2003 et Évolution Depuis 1998. 2003, p. 112. Available online: https://solidarites-sante.gouv.fr/IMG/pdf/Les_naissances_en_2010_et_leur_evolution_depuis_2003.pdf (accessed on 13 February 2022).
- Thonneau, P.; Marchand, S.; Tallec, A.; Ferial, M.-L.; Ducot, B.; Lansac, J.; Lopes, P.; Tabaste, J.-M.; Spira, A. Incidence and main causes of infertility in a resident population (1,850,000) of three French regions (1988–1989)*. Hum. Reprod. 1991, 6, 811–816. [Google Scholar] [CrossRef] [PubMed]
- Jungwirth, A.; Diemer, T.; Kopa, Z.; Krausz, C.; Minhas, S.; Tournaye, H. EAU Guidelines on Male Infertility; European Association of Urology: Arnhem, The Netherlands, 2018. [Google Scholar]
- Bonde, J.P.E.; Ernst, E.; Jensen, T.K.; Hjollund, N.H.; Kolstad, H.; Henriksen, T.B.; Scheike, T.; Giwercman, A.; Olsen, J.; Skakkebæk, N.E. Relation between semen quality and fertility: A population-based study of 430 first-pregnancy planners. Lancet 1998, 352, 1172–1177. [Google Scholar] [CrossRef]
- Moretti, E.; Capitani, S.; Figura, N.; Pammolli, A.; Federico, M.G.; Giannerini, V.; Collodel, G. The presence of bacteria species in semen and sperm quality. J. Assist. Reprod. Genet. 2008, 26, 47–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Francesco, M.A.; Negrini, R.; Ravizzola, G.; Galli, P.; Manca, N. Bacterial species present in the lower male genital tract: A five-year retrospective study. Eur. J. Contracept. Reprod. Health Care 2010, 16, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Hannachi, H.; Elloumi, H.; Hamdoun, M.; Kacem, K.; Zhioua, A.; Bahri, O. La bactériospermie: Effets sur les paramètres spermatiques. Gynécol. Obs. Fertil. Sénol. 2018, 46, 518–523. [Google Scholar] [CrossRef] [PubMed]
- Berktas, M.; Aydin, S.; Yilmaz, Y.; Cecen, K.; Bozkurt, H. Sperm motility changes after coincubation with various uropathogenic microorganisms: An in vitro experimental study. Int. Urol. Nephrol. 2008, 40, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Fraczek, M.; Wiland, E.; Piasecka, M.; Boksa, M.H.; Gączarzewicz, D.; Szumala-Kakol, A.; Kolanowski, T.; Beutin, L.; Kurpisz, M. Fertilizing potential of ejaculated human spermatozoa during in vitro semen bacterial infection. Fertil. Steril. 2014, 102, 711–719.e1. [Google Scholar] [CrossRef]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLOS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [Green Version]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The Human Microbiome Project. Nature 2007, 449, 804–810. [Google Scholar] [CrossRef]
- Pichon, M.; Lina, B.; Josset, L. Caractérisation et impact du microbiote bactérien respiratoire sur les maladies virales. Virologie 2018, 22, 161–172. [Google Scholar] [CrossRef]
- Moreno, I.; Codoñer, F.M.; Vilella, F.; Valbuena, D.; Martinez-Blanch, J.F.; Jimenez-Almazán, J.; Alonso, R.; Alamá, P.; Remohí, J.; Pellicer, A.; et al. Evidence that the endometrial microbiota has an effect on implantation success or failure. Am. J. Obstet. Gynecol. 2016, 215, 684–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organisation. WHO Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar]
- Saïdi, R.; Gruel, E.; Roset-Blessman, J.; Mousset-Siméon, N.; Menon, S.; Macé, B.; Rives, N. Evaluation morphologique des spermatozoïdes. Basic Clin. Androl. 2008, 18, 158–167. [Google Scholar] [CrossRef] [Green Version]
- Langevin, S.; Pichon, M.; Smith, E.; Morrison, J.; Bent, Z.; Green, R.; Barker, K.; Solberg, O.; Gillet, Y.; Javouhey, E.; et al. Early nasopharyngeal microbial signature associated with severe influenza in children: A retrospective pilot study. J. Gen. Virol. 2017, 98, 2425–2437. [Google Scholar] [CrossRef] [PubMed]
- Price, L.B.; Liu, C.M.; Johnson, K.E.; Aziz, M.; Lau, M.K.; Bowers, J.; Ravel, J.; Keim, P.S.; Serwadda, D.; Wawer, M.J.; et al. The Effects of Circumcision on the Penis Microbiome. PLoS ONE 2010, 5, e8422. [Google Scholar] [CrossRef] [Green Version]
- Molina, N.M.; Plaza-Díaz, J.; Vilchez-Vargas, R.; Sola-Leyva, A.; Vargas, E.; Mendoza-Tesarik, R.; Galán-Lázaro, M.; de Guevara, N.M.-L.; Tesarik, J.; Altmäe, S. Assessing the testicular sperm microbiome: A low-biomass site with abundant contamination. Reprod. Biomed. Online 2021, 43, 523–531. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Kazutaka, K.; Misakwa, K.; Kei-ichi, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Toh, H. Recent developments in the MAFFT multiple sequence alignment program. Brief. Bioinform. 2008, 9, 286–298. [Google Scholar] [CrossRef] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Faith, D.P. The Role of the Phylogenetic Diversity Measure, PD, in Bio-Informatics: Getting the Definition Right. Evol. Bioinform. 2006, 2, 277–283. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Knight, R. Global patterns in bacterial diversity. Proc. Natl. Acad. Sci. USA 2007, 104, 11436–11440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozupone, C.; Knight, R. UniFrac: A New Phylogenetic Method for Comparing Microbial Communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandal, S.; Van Treuren, W.; White, R.A.; Eggesbø, M.; Knight, R.T.; Peddada, S.D. Analysis of composition of microbiomes: A novel method for studying microbial composition. Microb. Ecol. Health Dis. 2015, 26, 27663. [Google Scholar] [CrossRef] [Green Version]
- Punzón-Jiménez, P.; Labarta, E. The impact of the female genital tract microbiome in women health and reproduction: A review. J. Assist. Reprod. Genet. 2021, 38, 2519–2541. [Google Scholar] [CrossRef]
- Weng, S.-L.; Chiu, C.-M.; Lin, F.-M.; Huang, W.-C.; Liang, C.; Yang, T.; Yang, T.-L.; Liu, C.-Y.; Wu, W.-Y.; Chang, Y.-A.; et al. Bacterial Communities in Semen from Men of Infertile Couples: Metagenomic Sequencing Reveals Relationships of Seminal Microbiota to Semen Quality. PLoS ONE 2014, 9, e110152. [Google Scholar] [CrossRef] [Green Version]
- Baud, D.; Pattaroni, C.; Vulliemoz, N.; Castella, V.; Marsland, B.J.; Stojanov, M. Sperm Microbiota and Its Impact on Semen Parameters. Front. Microbiol. 2019, 10, 234. [Google Scholar] [CrossRef] [Green Version]
- Lundy, S.D.; Sangwan, N.; Parekh, N.V.; Selvam, M.K.P.; Gupta, S.; McCaffrey, P.; Bessoff, K.; Vala, A.; Agarwal, A.; Sabanegh, E.S.; et al. Functional and Taxonomic Dysbiosis of the Gut, Urine, and Semen Microbiomes in Male Infertility. Eur. Urol. 2021, 79, 826–836. [Google Scholar] [CrossRef]
- Hooks, K.B.; O’Malley, M.A. Dysbiosis and Its Discontents. mBio 2017, 8, e01492-17. [Google Scholar] [CrossRef] [Green Version]
- Ho, C.L.T.; Vaughan-Constable, D.R.; Ramsay, J.; Jayasena, C.; Tharakan, T.; Yap, T.; Whiteman, I.; Graham, N.; Minhas, S.; Homa, S.T. The relationship between genitourinary microorganisms and oxidative stress, sperm DNA fragmentation and semen parameters in infertile men. Andrologia 2021, 54, e14322. [Google Scholar] [CrossRef]
- Kiessling, A.A.; Desmarais, B.M.; Yin, H.-Z.; Loverde, J.; Eyre, R.C. Detection and identification of bacterial DNA in semen. Fertil. Steril. 2008, 90, 1744–1756. [Google Scholar] [CrossRef] [PubMed]
- Hou, D.; Zhou, X.; Zhong, X.; Settles, M.; Herring, J.; Wang, L.; Abdo, Z.; Forney, L.J.; Xu, C. Microbiota of the seminal fluid from healthy and infertile men. Fertil. Steril. 2013, 100, 1261–1269.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menkveld, R. Clinical significance of the low normal sperm morphology value as proposed in the fifth edition of the WHO Laboratory Manual for the Examination and Processing of Human Semen. Asian J. Androl. 2010, 12, 47–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyman, R.W.; Herndon, C.N.; Jiang, H.; Palm, C.; Fukushima, M.; Bernstein, D.; Vo, K.C.; Zelenko, Z.; Davis, R.W.; Giudice, L.C. The dynamics of the vaginal microbiome during infertility therapy with in vitro fertilization-embryo transfer. J. Assist. Reprod. Genet. 2012, 29, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Haahr, T. Vaginal Microbiota and IVF Outcomes—Development of a Simple Diagnostic Tool to Predict Patients at Risk of a Poor Reproductive Outcome. In Human Reproduction; Oxford University Press: Oxford, UK, 2017. [Google Scholar]
- Chełmońska, A. The influence of Haemophilus somnus on bull sperms examined in vitro. Pol. Arch. Weter. 1990, 30, 141–154. [Google Scholar] [PubMed]
- Kyono, K.; Hashimoto, T.; Kikuchi, S.; Nagai, Y.; Sakuraba, Y. A pilot study and case reports on endometrial microbiota and pregnancy outcome: An analysis using 16S rRNA gene sequencing among IVF patients, and trial therapeutic intervention for dysbiotic endometrium. Reprod. Med. Biol. 2019, 18, 72–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valcarce, D.G.; Genovés, S.; Riesco, M.F.; Martorell, P.; Herraez, P.; Ramón, D.; Robles, V. Probiotic administration improves sperm quality in asthenozoospermic human donors. Benef. Microbes 2017, 8, 193–206. [Google Scholar] [CrossRef]
- Alvarez, C.; Castilla, J.; Martínez, L.; Ramírez, J.; Vergara, F.; Gaforio, J. Biological variation of seminal parameters in healthy subjects. Hum. Reprod. 2003, 18, 2082–2088. [Google Scholar] [CrossRef] [Green Version]

| Bray–Curtis Index | Unifrac Weighted Index | ||||
|---|---|---|---|---|---|
| Pseudo-F | p-Value | Pseudo-F | p-Value | ||
| Sperm count (M of spermatozoa per ejaculation) | ≤39 vs. >39 | 3.05 | 0.022 * | 3.71 | 0.025 * |
| Motility (% of mobile spermatozoa) | <32% vs. ≥32% | 1.73 | 0.098 | 2.61 | 0.074 |
| Morphology (% of morphologically normal spermatozoa) | <4% vs. ≥4% | 0.80 | 0.508 | 0.59 | 0.585 |
| Studies | Weng et al., 2014 [29] | Baud et al., 2019 [30] | Hou et al., 2013 [35] |
|---|---|---|---|
| Population | 36 normal semen parameters, 60 men with semen abnormalities | 26 normal semen parameters, 68 men with semen abnormalities | 19 healthy sperm donors, 58 men with semen abnormalities |
| Technology | MiSeq system (Illumina) Synthesis with reversible dye terminators | MiSeq system (Illumina) Synthesis with reversible dye terminators | Roche 454 GS-FLX (Roche-454 Life Sciences) Pyrosequencing |
| Targeted sequences | V4 16S rRNA gene | V1–V2 16S rRNA gene | V1–V2 16S rRNA gene |
| Semen parameters | Volume, mobility, count, morphology, anti-sperm antibody, leucocytes, and CASA | Count, mobility, and morphology | Volume, mobility, and count |
| Most common species | Lactobacillus, Gardnerella, Pseudomonas, Prevotella, Streptococcus, Finegoldia, Haemophilus, and Rhodanobacter | Corynebacterium, Prevotella, Lactobacillus, Streptococcus, Staphylococcus, Finegoldia, Haemophilus, and Burkholderia | Ralstonia, Lactobacillus, Corynebacterium, Streptococcus, Prevotella, Finegoldia, Anaerococcus, and Peptinophilus |
| Groups | Pseudomonas predominant (G1), Lactobacillus predominant (G2), Prevotella predominant (G3) | Prevotella enriched, Lactobacillus enriched, polymicrobial | 6 groups characterized by high proportion of same species, but none predominant |
| Associations | G2 associated with normal semen parameters; G1 and G3 associated with abnormal semen parameters | Prevotella associated with defective semen mobility; Lactobacillus associated with normal semen morphology; Staphylococcus associated with normal semen parameters | No clusters were associated with semen parameters Anaerococcus: negative association with semen quality in general |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gachet, C.; Prat, M.; Burucoa, C.; Grivard, P.; Pichon, M. Spermatic Microbiome Characteristics in Infertile Patients: Impact on Sperm Count, Mobility, and Morphology. J. Clin. Med. 2022, 11, 1505. https://doi.org/10.3390/jcm11061505
Gachet C, Prat M, Burucoa C, Grivard P, Pichon M. Spermatic Microbiome Characteristics in Infertile Patients: Impact on Sperm Count, Mobility, and Morphology. Journal of Clinical Medicine. 2022; 11(6):1505. https://doi.org/10.3390/jcm11061505
Chicago/Turabian StyleGachet, Clémence, Manon Prat, Christophe Burucoa, Philippe Grivard, and Maxime Pichon. 2022. "Spermatic Microbiome Characteristics in Infertile Patients: Impact on Sperm Count, Mobility, and Morphology" Journal of Clinical Medicine 11, no. 6: 1505. https://doi.org/10.3390/jcm11061505
APA StyleGachet, C., Prat, M., Burucoa, C., Grivard, P., & Pichon, M. (2022). Spermatic Microbiome Characteristics in Infertile Patients: Impact on Sperm Count, Mobility, and Morphology. Journal of Clinical Medicine, 11(6), 1505. https://doi.org/10.3390/jcm11061505