

Moderate Aerobic Exercise Regulates Follicular Dysfunction by Initiating Brain-Derived Neurotrophic Factor (BDNF)-Mediated Anti-Apoptotic Signaling Pathways in Polycystic Ovary Syndrome

 ,
,
Abstract
1. Introduction
2. Subjects and Methods
2.1. Animal Models and Groups
2.2. Implementation of Moderate Aerobic Exercise
2.3. Isolation and Culture of Granulosa Cells (GCs)
2.4. CCK8 Assay
2.5. Cell Apoptosis Assessment
2.6. Hematoxylin and Eosin (H&E) Staining
2.7. Serum Hormone Measurement
2.8. Nissl Staining
2.9. ELISA
2.10. Immunohistochemistry (IHC) and Immunofluorescence (IF)
3. TUNEL Analysis
3.1. Quantitative Real-Time PCR (qRT-PCR)
3.2. Western Blot
3.3. Statistical Analysis
4. Results
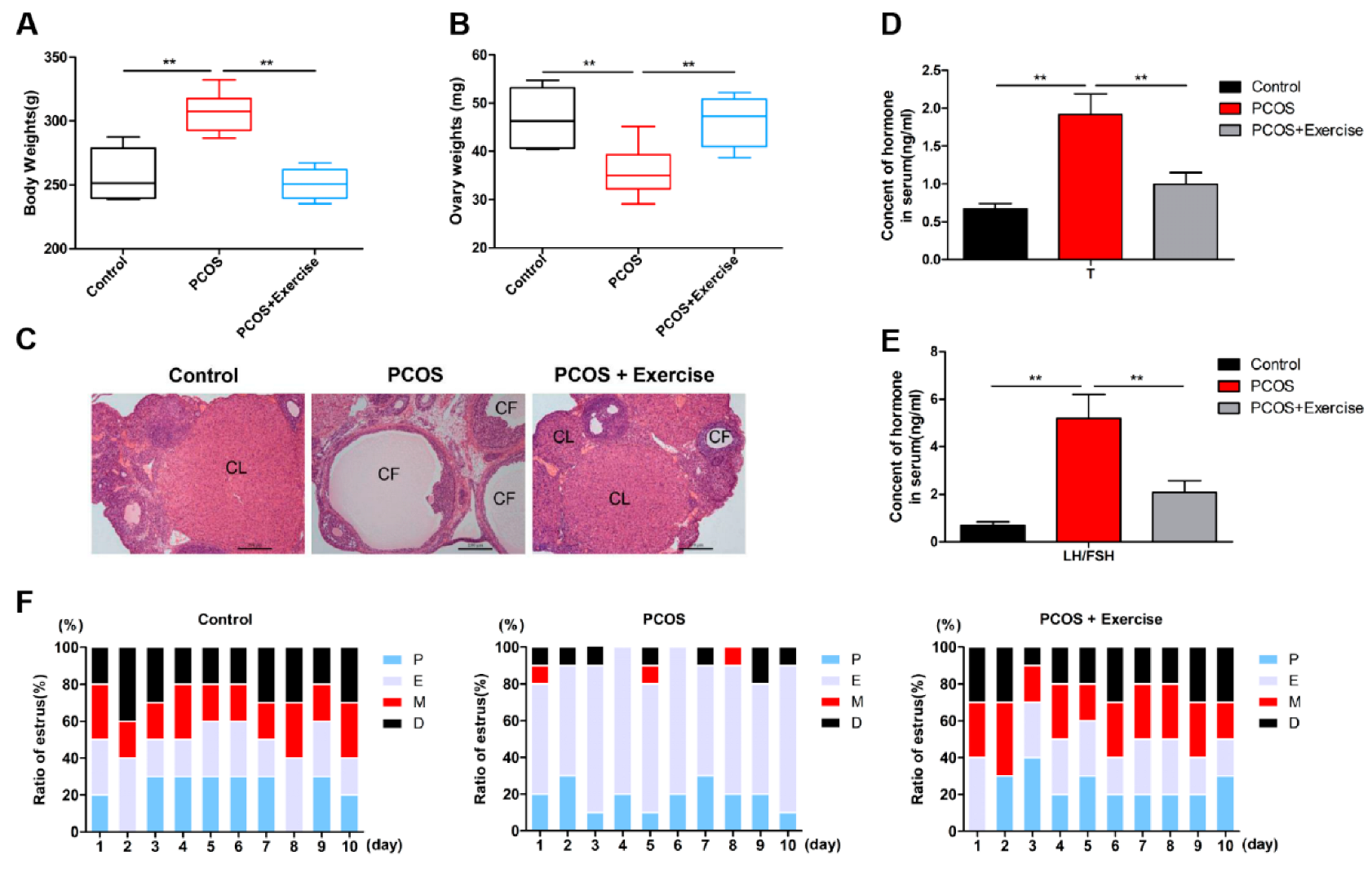
4.1. Effects of Moderate Aerobic Exercise on Ovarian Dysfunction in DHEA-Induced PCOS Rats
4.2. Location and Expression of BDNF in the Hippocampus of PCOS Rats after Moderate Aerobic Exercise
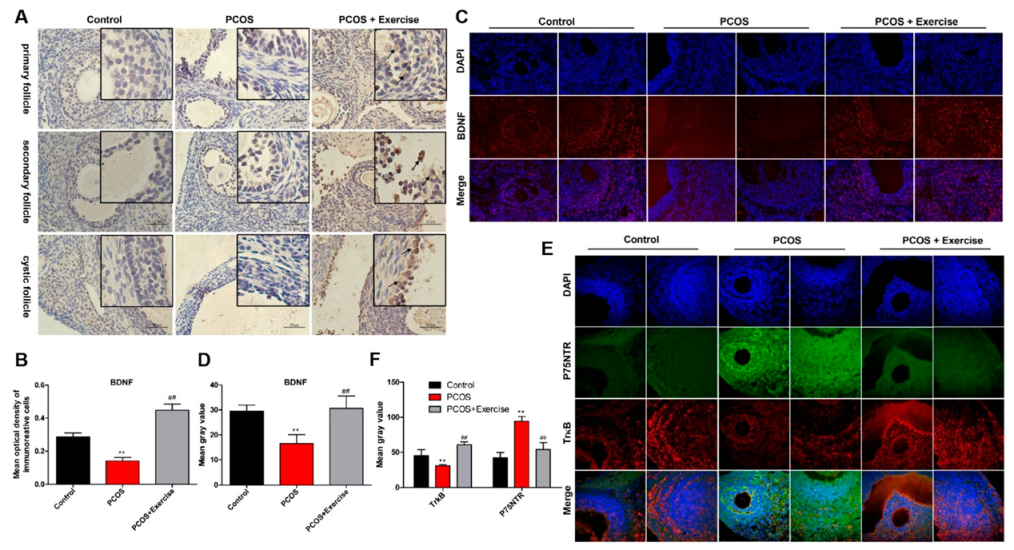
4.3. Location and Expression of BDNF, TrkB, and p75NTR in Ovarian Follicles of PCOS Rats after Moderate Aerobic Exercise
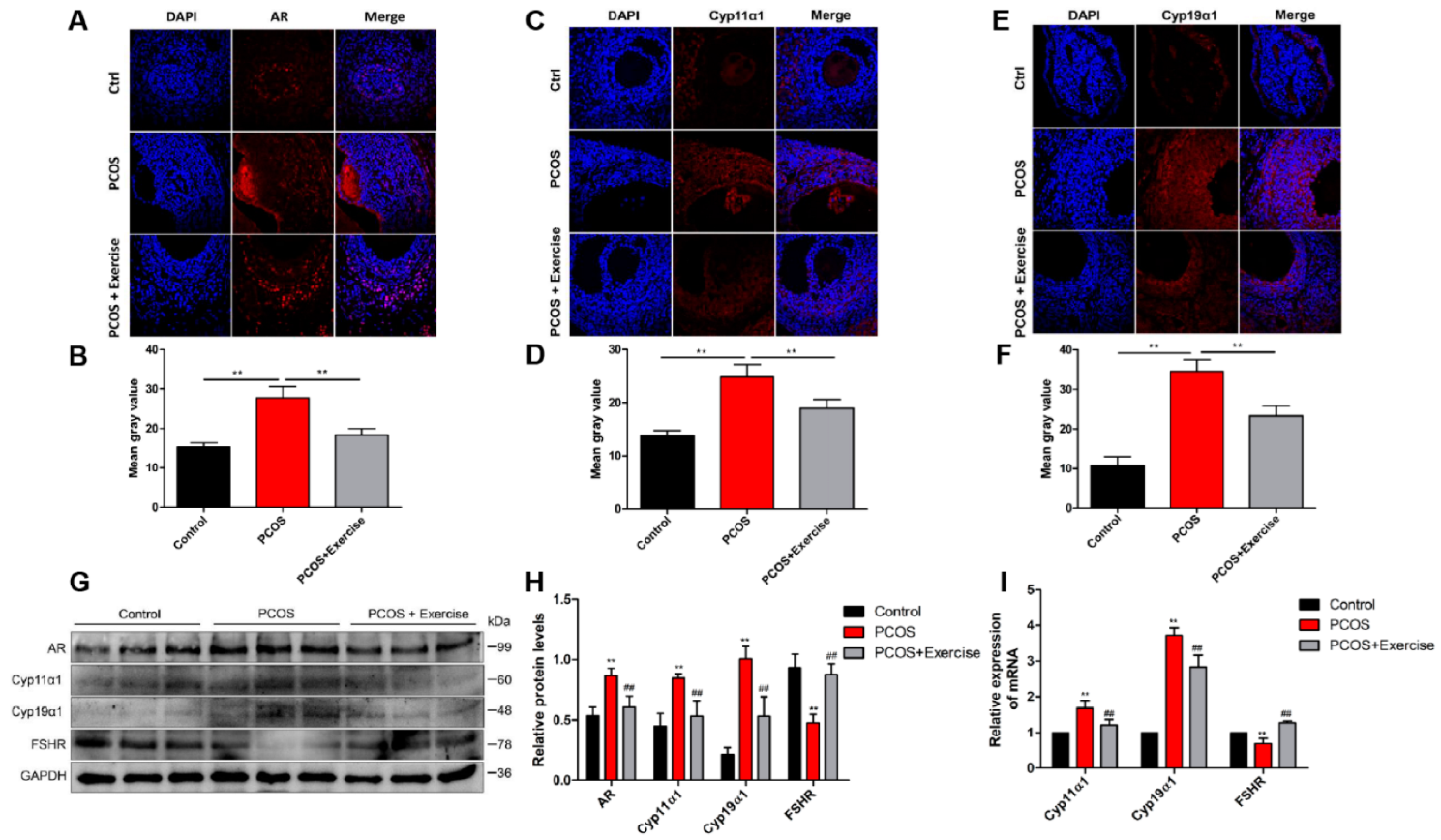
4.4. Activated BDNF Signaling in the Ovary of DHEA-Induced PCOS Rats
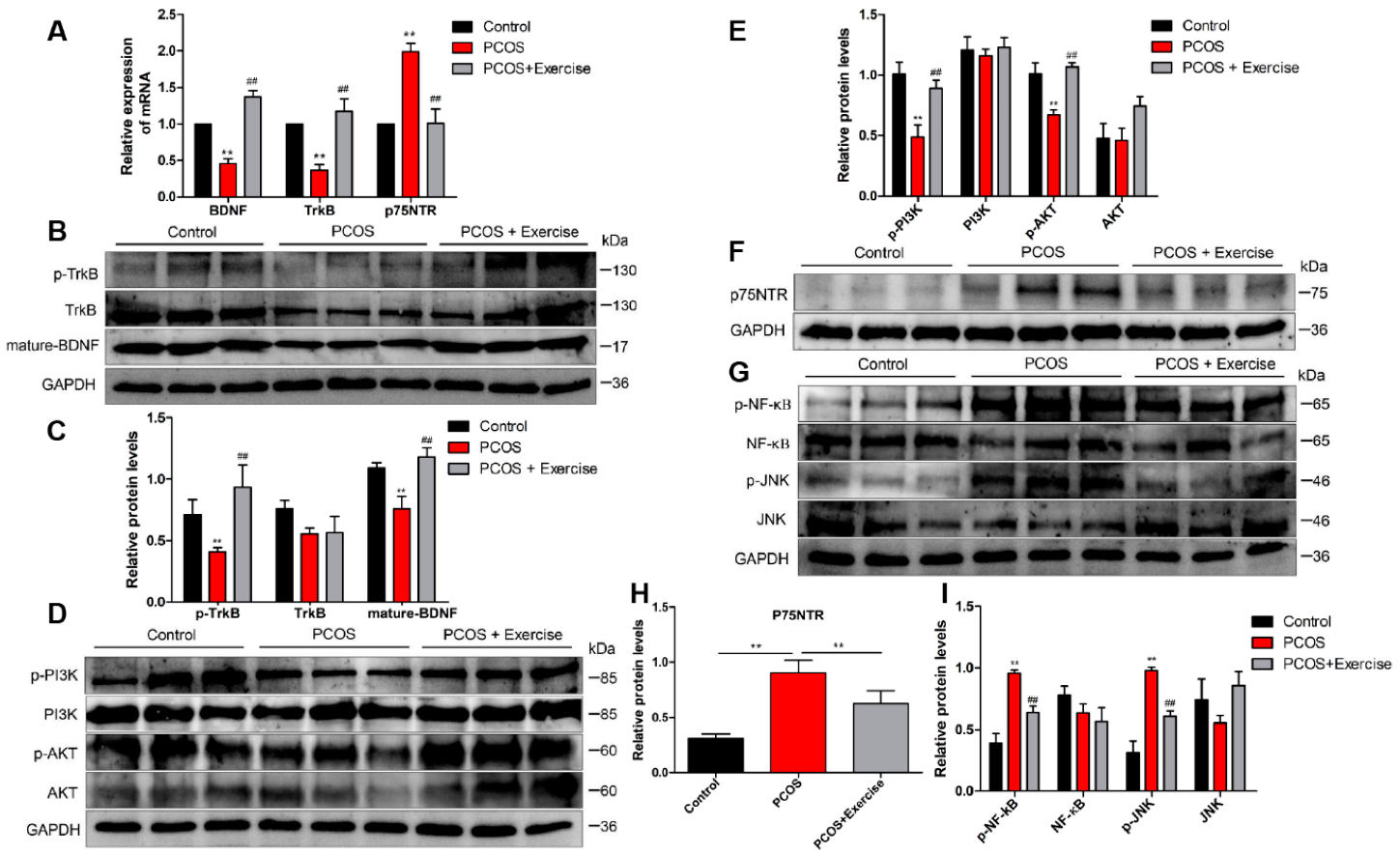
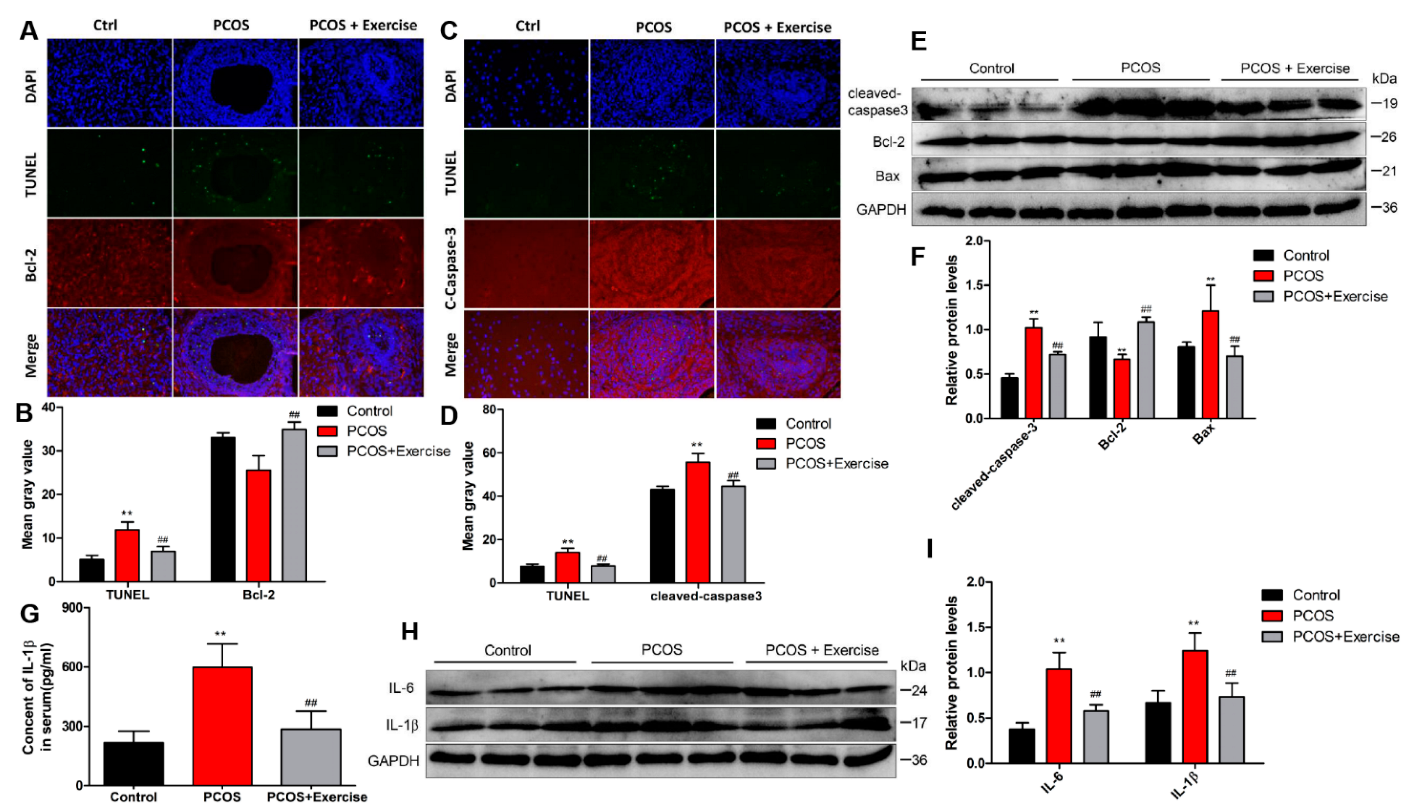
4.5. Moderate Aerobic Exercise Reduced Apoptosis and Inflammation of Ovarian Tissue in PCOS Rats
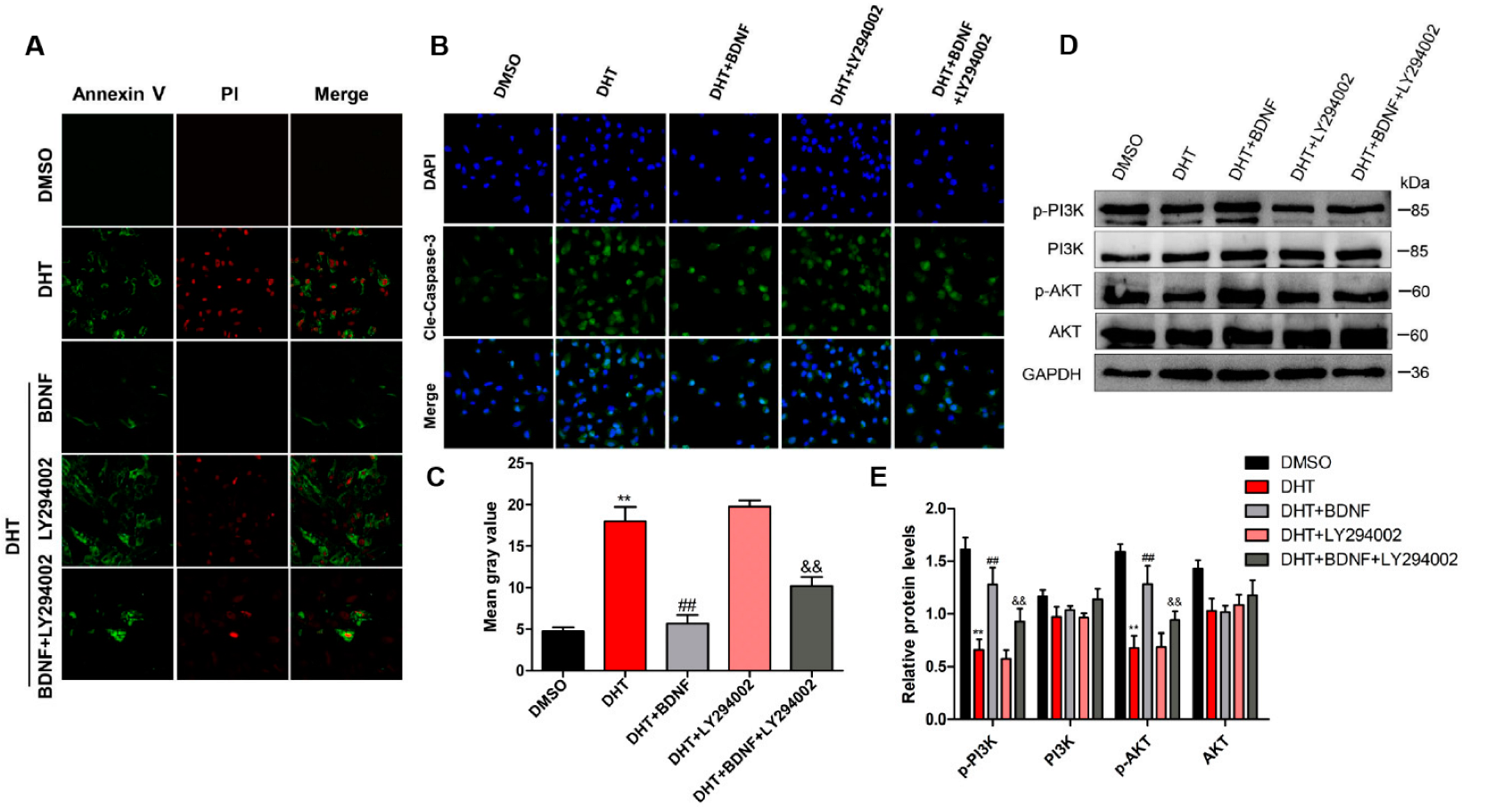
4.6. BDNF Possibly Alleviates DHT-Induced GC Apoptosis by Upregulating the PI3K/AKT Pathway
5. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Azziz, R. Polycystic Ovary Syndrome. Obstet. Gynecol. 2018, 132, 321–336. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wu, D.; Guo, H.; Li, M. Hyperandrogenemia and insulin resistance: The chief culprit of polycystic ovary syndrome. Life Sci. 2019, 236, 116940. [Google Scholar] [CrossRef] [PubMed]
- Dewailly, D.; Robin, G.; Peigne, M.; Decanter, C.; Pigny, P.; Catteau-Jonard, S. Interactions between androgens, FSH, anti-Müllerian hormone and estradiol during folliculogenesis in the human normal and polycystic ovary. Hum. Reprod. Updat. 2016, 22, 709–724. [Google Scholar] [CrossRef] [PubMed]
- Aflatounian, A.; Edwards, M.C.; Paris, V.R.; Bertoldo, M.J.; Desai, R.; Gilchrist, R.B.; Ledger, W.L.; Handelsman, D.J.; Walters, K.A. Androgen signaling pathways driving reproductive and metabolic phenotypes in a PCOS mouse model. J. Endocrinol. 2020, 245, 381–395. [Google Scholar] [CrossRef] [PubMed]
- da Costa, C.S.; Oliveira, T.F.; Freitas-Lima, L.C.; Padilha, A.S.; Krause, M.; Carneiro, M.T.W.; Salgado, B.S.; Graceli, J.B. Subacute cadmium exposure disrupts the hypothalamic-pituitary-gonadal axis, leading to polycystic ovarian syndrome and premature ovarian failure features in female rats. Environ. Pollut. 2020, 269, 116154. [Google Scholar] [CrossRef]
- Wang, D.; Wang, T.; Wang, R.; Zhang, X.; Wang, L.; Xiang, Z.; Zhuang, L.; Shen, S.; Wang, H.; Gao, Q.; et al. Suppression of p66Shc prevents hyperandrogenism-induced ovarian oxidative stress and fibrosis. J. Transl. Med. 2020, 18, 84. [Google Scholar] [CrossRef]
- Wang, D.; Weng, Y.; Zhang, Y.; Wang, R.; Wang, T.; Zhou, J.; Shen, S.; Wang, H.; Wang, Y. Exposure to hyperandrogen drives ovarian dysfunction and fibrosis by activating the NLRP3 inflammasome in mice. Sci. Total Environ. 2020, 745, 141049. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Ma, Y.; Tong, X.; Yang, W.; Dai, Y.; Pan, Y.; Ren, P.; Liu, L.; Fan, H.-Y.; Zhang, Y.; et al. Metformin inhibits testosterone-induced endoplasmic reticulum stress in ovarian granulosa cells via inactivation of p38 MAPK. Hum. Reprod. 2020, 35, 1145–1158. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, A.S.L.; Edwards, M.C.; Desai, R.; Jimenez, M.; Gilchrist, R.B.; Handelsman, D.J.; Walters, K.A. Neuroendocrine androgen action is a key extraovarian mediator in the development of polycystic ovary syndrome. Proc. Natl. Acad. Sci. USA 2017, 114, E3334–E3343. [Google Scholar] [CrossRef]
- Walters, K.A.; Gilchrist, R.; Ledger, W.; Teede, H.J.; Handelsman, D.J.; Campbell, R.E. New Perspectives on the Pathogenesis of PCOS: Neuroendocrine Origins. Trends Endocrinol. Metab. 2018, 29, 841–852. [Google Scholar] [CrossRef]
- Ruddenklau, A.; Campbell, R.E. Neuroendocrine Impairments of Polycystic Ovary Syndrome. Endocrinology 2019, 160, 2230–2242. [Google Scholar] [CrossRef]
- Febbraio, M.A. Exercise metabolism in 2016: Health benefits of exercise-more than meets the eye! Nat. Rev. Endocrinol. 2016, 13, 72–74. [Google Scholar] [CrossRef]
- Moran, L.J.; Hutchison, S.K.; Norman, R.; Teede, H.J. Lifestyle changes in women with polycystic ovary syndrome. Cochrane Database Syst. Rev. 2011, 3, CD007506. [Google Scholar] [CrossRef] [PubMed]
- Hakimi, O.; Cameron, L.-C. Effect of Exercise on Ovulation: A Systematic Review. Sports Med. 2016, 47, 1555–1567. [Google Scholar] [CrossRef]
- Zhang, Y.; Weng, Y.; Wang, D.; Wang, R.; Wang, L.; Zhou, J.; Shen, S.; Wang, H.; Wang, Y. Curcumin in Combination with Aerobic Exercise Improves Follicular Dysfunction via Inhibition of the Hyperandrogen-Induced IRE1α/XBP1 Endoplasmic Reticulum Stress Pathway in PCOS-Like Rats. Oxid. Med. Cell. Longev. 2021, 2021, 7382900. [Google Scholar] [CrossRef]
- Reycraft, J.T.; Islam, H.; Townsend, L.K.; Hayward, G.C.; Hazell, T.J.; Macpherson, R.E.K. Exercise Intensity and Recovery on Circulating Brain-derived Neurotrophic Factor. Med. Sci. Sports Exerc. 2019, 52, 1210–1217. [Google Scholar] [CrossRef]
- Alves, R.L.; Cardoso, B.R.L.; Ramos, I.P.R.; Oliveira, B.D.S.; dos Santos, M.L.; de Miranda, A.S.; de Almeida, T.C.S.; Vieira, M.A.R.; Machado, F.S.; Ferreira, A.J.; et al. Physical training improves exercise tolerance, cardiac function and promotes changes in neurotrophins levels in chagasic mice. Life Sci. 2019, 232, 116629. [Google Scholar] [CrossRef]
- Chow, R.; Wessels, J.M.; Foster, W.G. Brain-derived neurotrophic factor (BDNF) expression and function in the mammalian reproductive Tract. Hum. Reprod. Updat. 2020, 26, 545–564. [Google Scholar] [CrossRef]
- Buyuk, E.; Seifer, D.B. Follicular-fluid neurotrophin levels in women undergoing assisted reproductive technology for different etiologies of infertility. Fertil. Steril. 2008, 90, 1611–1615. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, D.; Zhang, M.; Li, N.; Lu, S.; Du, Y.; Chen, Z.-J. Effects of brain-derived neurotrophic factor on oocyte maturation and embryonic development in a rat model of polycystic ovary syndrome. Reprod. Fertil. Dev. 2016, 28, 1904–1915. [Google Scholar] [CrossRef]
- Wang, F.; Xie, N.; Zhou, J.; Dai, M.; Zhang, Q.; Hardiman, P.J.; Qu, F. Molecular mechanisms underlying altered neurobehavioural development of female offspring of mothers with polycystic ovary syndrome: FOS-mediated regulation of neurotrophins in placenta. eBioMedicine 2020, 60, 102993. [Google Scholar] [CrossRef]
- Russo, N.; Russo, M.; Daino, D.; Bucci, F.; Pluchino, N.; Casarosa, E.; Artini, P.G.; Cela, V.; Luisi, M.; Genazzani, A.R. Polycystic ovary syndrome: Brain-derived neurotrophic factor (BDNF) plasma and follicular fluid levels. Gynecol. Endocrinol. 2012, 28, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Kirmizi, D.A.; Baser, E.; Onat, T.; Demir Caltekin, M.; Yalvac, E.S.; Kara, M.; Gocmen, A.Y. Sexual function and depression in polycystic ovary syndrome: Is it associated with inflammation and neuromodulators? Neuropeptides 2020, 84, 102099. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.-M.; Wu, H.-C.; Sun, Z.; Lian, F.; Leung, P.C.K. Neurotrophins and glial cell line-derived neurotrophic factor in the ovary: Physiological and pathophysiological implications. Hum. Reprod. Updat. 2019, 25, 224–242. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Liu, L.; Liu, H.; Zheng, H.; Sun, H.; Ji, J.; Sun, Y.; Yang, T.; Zhao, H.; Qi, F.; et al. The Bu Shen Yi Sui Formula Promotes Axonal Regeneration via Regulating the Neurotrophic Factor BDNF/TrkB and the Downstream PI3K/Akt Signaling Pathway. Front. Pharmacol. 2019, 10, 796. [Google Scholar] [CrossRef]
- Zanin, J.; Montroull, L.E.; Volosin, M.; Friedman, W.J. The p75 Neurotrophin Receptor Facilitates TrkB Signaling and Function in Rat Hippocampal Neurons. Front. Cell. Neurosci. 2019, 13, 485. [Google Scholar] [CrossRef]
- Yu, F.; Liu, Y.; Xu, J. Pro-BDNF Contributes to Hypoxia/Reoxygenation Injury in Myocardial Microvascular Endothelial Cells: Roles of Receptors p75NTR and Sortilin and Activation of JNK and Caspase 3. Oxid. Med. Cell. Longev. 2018, 2018, 3091424. [Google Scholar] [CrossRef]
- Podyma, B.; Parekh, K.; Güler, A.D.; Deppmann, C.D. Metabolic homeostasis via BDNF and its receptors. Trends Endocrinol. Metab. 2021, 32, 488–499. [Google Scholar] [CrossRef]
- Zhao, X.; Du, F.; Liu, X.; Ruan, Q.; Wu, Z.; Lei, C.; Deng, Y.; Luo, C.; Jiang, J.; Shi, D.; et al. Brain-derived neurotrophic factor (BDNF) is expressed in buffalo (Bubalus bubalis) ovarian follicles and promotes oocyte maturation and early embryonic development. Theriogenology 2019, 130, 79–88. [Google Scholar] [CrossRef]
- Wrann, C.D.; White, J.P.; Salogiannnis, J.; Laznik-Bogoslavski, D.; Wu, J.; Ma, D.; Lin, J.D.; Greenberg, M.E.; Spiegelman, B.M. Exercise Induces Hippocampal BDNF through a PGC-1α/FNDC5 Pathway. Cell Metab. 2013, 18, 649–659. [Google Scholar] [CrossRef]
- Sleiman, S.F.; Henry, J.; Al-Haddad, R.; El Hayek, L.; Abou Haidar, E.; Stringer, T.; Ulja, D.; Karuppagounder, S.S.; Holson, E.B.; Ratan, R.R.; et al. Exercise promotes the expression of brain derived neurotrophic factor (BDNF) through the action of the ketone body β-hydroxybutyrate. eLife 2016, 5, e15092. [Google Scholar] [CrossRef]
- Seifer, D.B.; Feng, B.; Shelden, R.M.; Chen, S.; Dreyfus, C.F. Brain-Derived Neurotrophic Factor: A Novel Human Ovarian Follicular Protein. J. Clin. Endocrinol. Metab. 2002, 87, 655–659. [Google Scholar] [CrossRef]
- Kawamura, K.; Kawamura, N.; Mulders, S.M.; Gelpke, M.D.S.; Hsueh, A.J.W. Ovarian brain-derived neurotrophic factor (BDNF) promotes the development of oocytes into preimplantation embryos. Proc. Natl. Acad. Sci. USA 2005, 102, 9206–9211. [Google Scholar] [CrossRef]
- Manti, M.; Pui, H.; Edström, S.; Risal, S.; Lu, H.; Lindgren, E.; Ohlsson, C.; Jerlhag, E.; Benrick, A.; Deng, Q.; et al. Excess of ovarian nerve growth factor impairs embryonic development and causes reproductive and metabolic dysfunction in adult female mice. FASEB J. 2020, 34, 14440–14457. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Chang, H.; Zhang, L.; Zhu, Y.; Leung, P.C.K. BMP2 increases the production of BDNF through the upregulation of proBDNF and furin expression in human granulosa-lutein cells. FASEB J. 2020, 34, 16129–16143. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, J.T.; Thieme, S.; Zettermann, P.; Leite, A.A.; Zanella, V.G.; Pilar, E.F.S.; Fonseca, F.P.; Mesquita, R.A.; Vargas, P.A.; dos Santos, J.N.; et al. Immunoexpression of BDNF, TrkB, and p75NTR receptors in peripheral neural lesions of the head and neck. J. Oral Pathol. Med. 2020, 50, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Colle, R.; Deflesselle, E.; Martin, S.; David, D.J.; Hardy, P.; Taranu, A.; Falissard, B.; Verstuyft, C.; Corruble, E. BDNF/TRKB/P75NTR polymorphisms and their consequences on antidepressant efficacy in depressed patients. Pharmacogenomics 2015, 16, 997–1013. [Google Scholar] [CrossRef]
- De Sousa, S.M.C.; Norman, R.J. Metabolic syndrome, diet and exercise. Best Pract. Res. Clin. Obstet. Gynaecol. 2016, 37, 140–151. [Google Scholar] [CrossRef]
- Mena, G.P.; Mielke, G.; Brown, W.J. The effect of physical activity on reproductive health outcomes in young women: A systematic review and meta-analysis. Hum. Reprod. Updat. 2019, 25, 541–563. [Google Scholar] [CrossRef]
- de Poli, R.A.B.; Lopes, V.H.F.; Lira, F.S.; Zagatto, A.M.; Jimenez-Maldonado, A.; Antunes, B.M. Peripheral BDNF and psycho-behavioral aspects are positively modulated by high-intensity intermittent exercise and fitness in healthy women. Sci. Rep. 2021, 11, 4113. [Google Scholar] [CrossRef]
- Wu, F.; Li, Z.; Cai, M.; Xi, Y.; Xu, Z.; Zhang, Z.; Li, H.; Zhu, W.; Tian, Z. Aerobic exercise alleviates oxidative stress-induced apoptosis in kidneys of myocardial infarction mice by inhibiting ALCAT1 and activating FNDC5/Irisin signaling pathway. Free Radic. Biol. Med. 2020, 158, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Li, Y.; Yang, Y.-J.; Zhang, D.-Y. The Effect of Moderate-Intensity Treadmill Exercise on Bone Mass and the Transcription of Peripheral Blood Mononuclear Cells in Ovariectomized Rats. Front. Physiol. 2021, 12, 729910. [Google Scholar] [CrossRef] [PubMed]
- Moran, L.J.; Noakes, M.; Clifton, P.; Buckley, J.; Brinkworth, G.; Thomson, R.; Norman, R.J. Predictors of Lifestyle Intervention Attrition or Weight Loss Success in Women with Polycystic Ovary Syndrome Who Are Overweight or Obese. Nutrients 2019, 11, 492. [Google Scholar] [CrossRef] [PubMed]
- Roh, H.-T.; So, W.-Y. The effects of aerobic exercise training on oxidant-antioxidant balance, neurotrophic factor levels, and blood-brain barrier function in obese and non-obese men. J. Sport Health Sci. 2017, 6, 447–453. [Google Scholar] [CrossRef]
- Negaresh, R.; Motl, R.W.; Zimmer, P.; Mokhtarzade, M.; Baker, J.S. Effects of exercise training on multiple sclerosis biomarkers of central nervous system and disease status: A systematic review of intervention studies. Eur. J. Neurol. 2019, 26, 711–721. [Google Scholar] [CrossRef]
- Walsh, J.J.; Tschakovsky, M.E. Exercise and circulating BDNF: Mechanisms of release and implications for the design of exercise interventions. Appl. Physiol. Nutr. Metab. 2018, 43, 1095–1104. [Google Scholar] [CrossRef]
- Marosi, K.; Mattson, M.P. BDNF mediates adaptive brain and body responses to energetic challenges. Trends Endocrinol. Metab. 2013, 25, 89–98. [Google Scholar] [CrossRef]
- Chen, S.-D.; Wu, C.-L.; Hwang, W.-C.; Yang, D.-I. More Insight into BDNF against Neurodegeneration: Anti-Apoptosis, Anti-Oxidation, and Suppression of Autophagy. Int. J. Mol. Sci. 2017, 18, 545. [Google Scholar] [CrossRef]
- Wang, X.; Ma, W.; Wang, T.; Yang, J.; Wu, Z.; Liu, K.; Dai, Y.; Zang, C.; Liu, W.; Liu, J. BDNF-TrkB and proBDNF-p75NTR/Sortilin Signaling Pathways are Involved in Mitochondria-Mediated Neuronal Apoptosis in Dorsal Root Ganglia after Sciatic Nerve Transection. CNS Neurol. Disord. Drug Targets 2020, 19, 66–82. [Google Scholar] [CrossRef]
- Jiajie, T.; Yanzhou, Y.; Hoi-Hung, A.C.; Zi-Jiang, C.; Wai-Yee, C. Conserved miR-10 family represses proliferation and induces apoptosis in ovarian granulosa cells. Sci. Rep. 2017, 7, 41304. [Google Scholar] [CrossRef]
- Chen, S.; Wang, F.; Liu, Z.; Zhao, Y.; Jiang, Y.; Chen, L.; Li, C.; Zhou, X. Brain-derived neurotrophic factor promotes proliferation and progesterone synthesis in bovine granulosa cells. J. Cell. Physiol. 2018, 234, 8776–8787. [Google Scholar] [CrossRef] [PubMed]
- Streiter, S.; Fisch, B.; Sabbah, B.; Ao, A.; Abir, R. The importance of neuronal growth factors in the ovary. Mol. Hum. Reprod. 2015, 22, 3–17. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward | Reverse |
|---|---|---|
| BDNF | 5′-GTGTGACAGTATTAGCGAGTGGG-3′ | 5′-ACGATTGGGTAGTTCGGCATT-3′ |
| TrkB | 5′-GGCATCACCAACAGTCAGC-3′ | 5′-GCATCCTTCAGGGTCTTCA-3′ |
| p75NTR | 5′-AGGGCACATACTCAGACGAA-3′ | 5′-AGATGGAGCAATAGACAGGAAT-3′ |
| FSHR | 5′-CAACCTCCGATATCTGTTAATA-3′ | 5′-CATTCTTACTCAGCCATACAGT-3′ |
| Cyp11α1 | 5′-GGATGCGTCGATACTCTTCTCA-3′ | 5′-GGACGATTCGGTCTTTCTTCCA-3′ |
| Cyp19α1 | 5′-AACCCGAGCCTTTGGAGAA-3′ | 5′-GGCCCGTCAGAGCTTTCA-3′ |
| GAPDH | 5′-AGGTCGTGTGAGGGATTG-3′ | 5′-TTAGTAGTAGTAGGGGGGGTCA-3′ |
| Group | FBG | p |
|---|---|---|
| Ctrl | 3.76 ± 0.47 | - |
| PCOS | 4.68 ± 1.20 | 0.095 |
| PCOS + Exercise | 4.04 ± 0.62 | 0.253 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Chen, D.; Wang, D.; Wang, L.; Weng, Y.; Wang, H.; Wu, X.; Wang, Y. Moderate Aerobic Exercise Regulates Follicular Dysfunction by Initiating Brain-Derived Neurotrophic Factor (BDNF)-Mediated Anti-Apoptotic Signaling Pathways in Polycystic Ovary Syndrome. J. Clin. Med. 2022, 11, 5584. https://doi.org/10.3390/jcm11195584
Zhang Y, Chen D, Wang D, Wang L, Weng Y, Wang H, Wu X, Wang Y. Moderate Aerobic Exercise Regulates Follicular Dysfunction by Initiating Brain-Derived Neurotrophic Factor (BDNF)-Mediated Anti-Apoptotic Signaling Pathways in Polycystic Ovary Syndrome. Journal of Clinical Medicine. 2022; 11(19):5584. https://doi.org/10.3390/jcm11195584
Chicago/Turabian StyleZhang, Yaling, Dejian Chen, Daojuan Wang, Lei Wang, Yajing Weng, Hongwei Wang, Xiaoke Wu, and Yong Wang. 2022. "Moderate Aerobic Exercise Regulates Follicular Dysfunction by Initiating Brain-Derived Neurotrophic Factor (BDNF)-Mediated Anti-Apoptotic Signaling Pathways in Polycystic Ovary Syndrome" Journal of Clinical Medicine 11, no. 19: 5584. https://doi.org/10.3390/jcm11195584
APA StyleZhang, Y., Chen, D., Wang, D., Wang, L., Weng, Y., Wang, H., Wu, X., & Wang, Y. (2022). Moderate Aerobic Exercise Regulates Follicular Dysfunction by Initiating Brain-Derived Neurotrophic Factor (BDNF)-Mediated Anti-Apoptotic Signaling Pathways in Polycystic Ovary Syndrome. Journal of Clinical Medicine, 11(19), 5584. https://doi.org/10.3390/jcm11195584