
Olive Pomace Phenolic Compounds: From an Agro-Industrial By-Product to a Promising Ocular Surface Protection for Dry Eye Disease

 ,
,  , , , and
, , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Extract Preparation
2.2. In Vitro Immunosuppressive Effect
2.2.1. Cell Isolation and Culture
2.2.2. Preparation of Phenolic Solutions
2.2.3. hTCD4+ Cell Proliferation Inhibition Study
2.3. Desiccating Stress (DS) Mouse Model
2.3.1. Animals and DS Conditions
2.3.2. Topical Treatments
2.3.3. Assessment of Clinical DED Signs
2.3.4. Flow Cytometry Analysis (FACS): % Count of CD3+, CD4+ and CD8+ in Lymph Nodes
2.3.5. Cytokine/Chemokine Gene Expression
2.4. Statistical Analysis
3. Results
3.1. In Vitro Effect of HT and OPT3 on hTCD4+ Cell Proliferation
3.2. Effect of HT and OPT3 on the DS-Induced DED Animal Model
3.2.1. CFS
3.2.2. Tear Volume
3.2.3. FACS Analysis: % Count of CD3+, CD4+ and CD8+ in Cervical Lymph Nodes
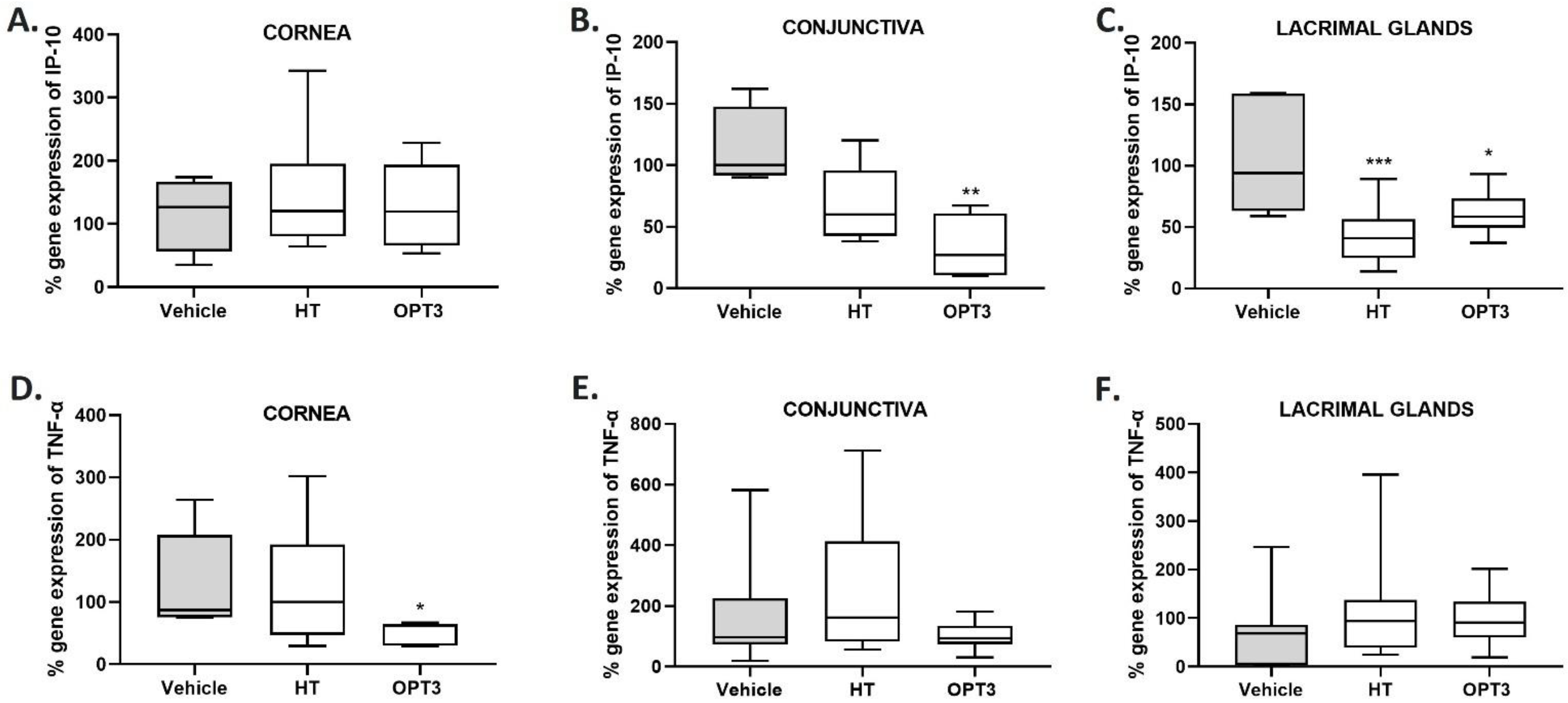
3.2.4. IP-10 and TNF-α Gene Expression in Cornea, Conjunctiva, and Lacrimal Glands
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| APC | Allophycocyanin |
| APC-Cy7 | Allophycocyanin-cyanine7 |
| BSA | N,O-bis(trimethylsilyl)acetamide |
| CFS | Corneal fluorescein staining |
| DED | Dry eye disease |
| DPBS | Dulbecco’s phosphate buffered saline |
| DS | Desiccating stress |
| EDTA | Ethylenediaminetetraacetic acid |
| EtOH | Ethanol |
| FACS | Flow cytometry analysis |
| FBS | Fetal bovine serum |
| FITC | Fluorescein isothiocyanate |
| HPRT | Hypoxanthine-guanine phosphoribosyl-transferase |
| HIV | Human immunodeficiency virus |
| HT | Hydroxytyrosol |
| hTCD4+ | human CD4+ T cells |
| IL | Interleukin |
| IP-10 | Interferon γ-induced protein-10 |
| OP | Olive pomace |
| OPT3 | Selected crude olive pomace extract |
| PBMC | Peripheral blood mononuclear cells |
| PHA-M | Phytohemagglutinin—M form |
| qRT-PCR | Quantitative real-time polymerase chain reaction |
| RPMI | Roswell Park Memorial Institute |
| SEM | Standard error of the mean |
| TNF-α | Tumor necrosis factor-α |
References
- Stapleton, F.; Alves, M.; Bunya, V.Y.; Jalbert, I.; Lekhanont, K.; Malet, F.; Na, K.-S.; Schaumberg, D.; Od, S.D.; Uchino, M.; et al. TFOS DEWS II Epidemiology Report. Ocul. Surf. 2017, 15, 334–365. [Google Scholar] [CrossRef] [PubMed]
- McDonald, M.; Patel, D.A.; Keith, M.S.; Snedecor, S.J. Economic and Humanistic Burden of Dry Eye Disease in Europe, North America, and Asia: A Systematic Literature Review. Ocul. Surf. 2016, 14, 144–167. [Google Scholar] [CrossRef]
- Stern, M.E.; Schaumburg, C.S.; Pflugfelder, S.C. Dry eye as a mucosal autoimmune disease. Int. Rev. Immunol. 2013, 32, 19–41. [Google Scholar] [CrossRef] [PubMed]
- Pflugfelder, S.C.; Solomon, A.; Stern, M.E. The diagnosis and management of dry eye: A twenty-five-year review. Cornea 2000, 19, 644–649. [Google Scholar] [CrossRef] [PubMed]
- Enríquez-de-Salamanca, A.; Castellanos, E.; Stern, M.E.; Fernández, I.; Carreño, E.; García-Vázquez, C.; Herreras, J.M.; Calonge, M. Tear cytokine and chemokine analysis and clinical correlations in evaporative-type dry eye disease. Mol. Vis. 2010, 16, 862–873. [Google Scholar] [PubMed]
- Jones, L.; Downie, L.E.; Korb, D.; Benitez-del-Castillo, J.M.; Dana, R.; Deng, S.X.; Dong, P.N.; Geerling, G.; Hida, R.Y.; Liu, Y.; et al. TFOS DEWS II Management and Therapy Report. Ocul. Surf. 2017, 15, 575–628. [Google Scholar] [CrossRef]
- Garrott, H.M.; Walland, M.J. Glaucoma from topical corticosteroids to the eyelids. Clin. Exp. Ophthalmol. 2004, 32, 224–226. [Google Scholar] [CrossRef]
- Steven, P.; Cursiefen, C. Anti-inflammatory treatment in dry eye disease. Klin. Monbl. Augenheilkd. 2012, 229, 500–505. [Google Scholar] [CrossRef]
- Rice, M.L.; Wong, B.; Horn, P.S.; Yang, M.B. Cataract development associated with long-term glucocorticoid therapy in Duchenne muscular dystrophy patients. J. Am. Assoc. Pediatr. Ophthalmol. Strabismus 2018, 22, 192–196. [Google Scholar] [CrossRef]
- Kunert, K.S.; Tisdale, A.S.; Stern, M.E.; Smith, J.A.; Gipson, I.K. Analysis of Topical Cyclosporine Treatment of Patients with Dry Eye Syndrome: Effect on Conjunctival Lymphocytes. Arch. Ophthalmol. 2000, 118, 1489–1496. [Google Scholar] [CrossRef]
- Donnenfeld, E.; Pflugfelder, S.C. Topical Ophthalmic Cyclosporine: Pharmacology and Clinical Uses. Surv. Ophthalmol. 2009, 54, 321–338. [Google Scholar] [CrossRef] [PubMed]
- Lallemand, F.; Schmitt, M.; Bourges, J.L.; Gurny, R.; Benita, S.; Garrigue, J.S. Cyclosporine A delivery to the eye: A comprehensive review of academic and industrial efforts. Eur. J. Pharm. Biopharm. 2017, 117, 14–28. [Google Scholar] [CrossRef] [PubMed]
- Business Wire. Novaliq Announces Positive Topline Results of Phase 2 Clinical Trial Evaluating CyclASol® in Adults with Moderate to Severe Dry Eye Disease; Business Wire: San Francisco, CA, USA, 2016. [Google Scholar]
- Sheppard, J.D.; Scoper, S.V.; Samudre, S. Topical Loteprednol Pretreatment Reduces Cyclosporine Stinging in Chronic Dry Eye Disease. J. Ocul. Pharmacol. Ther. 2011, 27, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Perez, V.L.; Pflugfelder, S.C.; Zhang, S.; Shojaei, A.; Haque, R. Lifitegrast, a Novel Integrin Antagonist for Treatment of Dry Eye Disease. Ocul. Surf. 2016, 14, 207–215. [Google Scholar] [CrossRef]
- Donnenfeld, E.D.; Karpecki, P.M.; Majmudar, P.A.; Nichols, K.K.; Raychaudhuri, A.; Roy, M.; Semba, C.P. Safety of Lifitegrast Ophthalmic Solution 5.0% in Patients With Dry Eye Disease: A 1-Year, Multicenter, Randomized, Placebo-Controlled Study. Cornea 2016, 35, 741–748. [Google Scholar] [CrossRef]
- U.S. Food and Drug Administration. FDA Approves New Medication for Dry Eye Disease. Available online: https://www.fda.gov/news-events/press-announcements/fda-approves-new-medication-dry-eye-disease (accessed on 10 November 2021).
- Novartis Withdraws Marketing Application of Dry Eye Drug Xiidra in Europe-Eyewire+. Available online: https://eyewire.news/news/novartis-pulls-eu-filing-seeking-approval-of-xiidra-for-dry-eye-disease (accessed on 11 November 2021).
- Kumar, N.; Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotechnol. Reports 2019, 24, e00370. [Google Scholar] [CrossRef]
- Albuquerque, B.R.; Heleno, S.A.; Oliveira, M.B.P.; Barros, L.; Ferreira, I.C. Phenolic compounds: Current industrial applications, limitations and future challenges. Food Funct. 2021, 12, 14–29. [Google Scholar] [CrossRef]
- González, R.; Ballester, I.; López-Posadas, R.; Suárez, M.D.; Zarzuelo, A.; Martínez-Augustin, O.; Sánchez de Medina, F. Effects of flavonoids and other polyphenols on inflammation. Crit. Rev. Food Sci. Nutr. 2011, 51, 331–362. [Google Scholar] [CrossRef]
- Shakoor, H.; Feehan, J.; Apostolopoulos, V.; Platat, C.; Dhaheri, A.S.; Ali, H.I.; Ismail, L.C.; Bosevski, M.; Stojanovska, L. Immunomodulatory Effects of Dietary Polyphenols. Nutrients 2021, 13, 728. [Google Scholar] [CrossRef]
- Abengózar-Vela, A.; Schaumburg, C.S.; Stern, M.E.; Calonge, M.; Enríquez-de-Salamanca, A.; González-García, M.J. Topical Quercetin and Resveratrol Protect the Ocular Surface in Experimental Dry Eye Disease. Ocul. Immunol. Inflamm. 2019, 27, 1023–1032. [Google Scholar] [CrossRef]
- Abengózar-Vela, A.; Calonge, M.; Stern, M.E.; González-García, M.J.; Enríquez-De-Salamanca, A. Quercetin and resveratrol decrease the inflammatory and oxidative responses in human ocular surface epithelial cells. Investig. Ophthalmol. Vis. Sci. 2015, 56, 2709–2719. [Google Scholar] [CrossRef] [PubMed]
- Cavet, M.E.; Harrington, K.L.; Vollmer, T.R.; Ward, K.W.; Zhang, J.Z. Anti-inflammatory and anti-oxidative effects of the green tea polyphenol epigallocatechin gallate in human corneal epithelial cells. Mol. Vis. 2011, 17, 533–542. [Google Scholar] [PubMed]
- Soo Lee, H.; Chauhan, S.K.; Okanobo, A.; Nallasamy, N.; Dana, R. Therapeutic Efficacy of Topical Epigallocatechin Gallate in Murine Dry Eye. Cornea 2011, 30, 1465–1472. [Google Scholar] [CrossRef]
- Obied, H.K.; Allen, M.S.; Bedgood, D.R.; Prenzler, P.D.; Robards, K.; Stockmann, R. Bioactivity and analysis of biophenols recovered from olive mill waste. J. Agric. Food Chem. 2005, 53, 823–837. [Google Scholar] [CrossRef]
- Lozano-Sánchez, J.; Giambanelli, E.; Quirantes-Piné, R.; Cerretani, L.; Bendini, A.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Wastes generated during the storage of extra virgin olive oil as a natural source of phenolic compounds. J. Agric. Food Chem. 2011, 59, 11491–11500. [Google Scholar] [CrossRef]
- Obied, H.K.; Bedgood, D.R.; Prenzler, P.D.; Robards, K. Chemical screening of olive biophenol extracts by hyphenated liquid chromatography. Anal. Chim. Acta 2007, 603, 176–189. [Google Scholar] [CrossRef]
- Chanioti, S.; Tzia, C. Optimization of ultrasound-assisted extraction of oil from olive pomace using response surface technology: Oil recovery, unsaponifiable matter, total phenol content and antioxidant activity. LWT-Food Sci. Technol. 2017, 79, 178–189. [Google Scholar] [CrossRef]
- Kapellakis, I.E.; Tsagarakis, K.P.; Crowther, J.C. Olive oil history, production and by-product management. Rev. Environ. Sci. Biotechnol. 2008, 7, 1–26. [Google Scholar] [CrossRef]
- FAOSTAT. Food and Agriculture Organization Database. Available online: http://www.fao.org/home/search/en/?q=Olivemillwastesproduction (accessed on 19 July 2022).
- Gómez-Muñoz, B.; Hatch, D.J.; Bol, R.; Garca-Ruiz, R. The Compost of Olive Mill Pomace: From a Waste to a Resource-Environmental Benefits of Its Application in Olive Oil Groves. In Sustainable Development-Authoritative and Leading Edge Content for Environmental Management; IntechOpen: London, UK, 2012. [Google Scholar]
- Souilem, S.; El-Abbassi, A.; Kiai, H.; Hafidi, A.; Sayadi, S.; Galanakis, C.M. Olive oil production sector: Environmental effects and sustainability challenges. In Olive Mill Waste: Recent Advances for Sustainable Management; Academic Press: Cambridge, MA, USA, 2017; pp. 1–28. [Google Scholar]
- Romero-García, J.M.; Niño, L.; Martínez-Patiño, C.; Álvarez, C.; Castro, E.; Negro, M.J. Biorefinery based on olive biomass. State of the art and future trends. Bioresour. Technol. 2014, 159, 421–432. [Google Scholar] [CrossRef]
- Roig, A.; Cayuela, M.L.; Sánchez-Monedero, M.A. An overview on olive mill wastes and their valorisation methods. Waste Manag. 2006, 26, 960–969. [Google Scholar] [CrossRef]
- Dermeche, S.; Nadour, M.; Larroche, C.; Moulti-Mati, F.; Michaud, P. Olive mill wastes: Biochemical characterizations and valorization strategies. Process Biochem. 2013, 48, 1532–1552. [Google Scholar] [CrossRef]
- Napolitano, A.; De Lucia, M.; Panzella, L.; d’Ischia, M. The Chemistry of Tyrosol and Hydroxytyrosol: Implications for Oxidative Stress. In Olives and Olive Oil in Health and Disease Prevention; Elsevier: Amsterdam, The Netherlands, 2010; pp. 1225–1232. [Google Scholar]
- Dierkes, G.; Krieger, S.; Dück, R.; Bongartz, A.; Schmitz, O.J.; Hayen, H. High-Performance Liquid Chromatography–Mass Spectrometry Profiling of Phenolic Compounds for Evaluation of Olive Oil Bitterness and Pungency. J. Agric. Food Chem. 2012, 60, 7597–7606. [Google Scholar] [CrossRef] [PubMed]
- Savarese, M.; De Marco, E.; Sacchi, R. Characterization of phenolic extracts from olives (Olea europaea cv. Pisciottana) by electrospray ionization mass spectrometry. Food Chem. 2007, 105, 761–770. [Google Scholar] [CrossRef]
- Katsinas, N.; Bento da Silva, A.; Enríquez-de-Salamanca, A.; Fernández, N.; Bronze, M.R.; Rodríguez-Rojo, S. Pressurized Liquid Extraction Optimization from Supercritical Defatted Olive Pomace: A Green and Selective Phenolic Extraction Process. ACS Sustain. Chem. Eng. 2021, 9, 5590–5602. [Google Scholar] [CrossRef]
- De Marco, E.; Savarese, M.; Paduano, A.; Sacchi, R. Characterization and fractionation of phenolic compounds extracted from olive oil mill wastewaters. Food Chem. 2007, 104, 858–867. [Google Scholar] [CrossRef]
- De Leonardis, A.; Aretini, A.; Alfano, G.; Macciola, V.; Ranalli, G. Isolation of a hydroxytyrosol-rich extract from olive leaves (Olea Europaea L.) and evaluation of its antioxidant properties and bioactivity. Eur. Food Res. Technol. 2007, 226, 653–659. [Google Scholar] [CrossRef]
- Fernández-Mar, M.I.; Mateos, R.; García-Parrilla, M.C.; Puertas, B.; Cantos-Villar, E. Bioactive compounds in wine: Resveratrol, hydroxytyrosol and melatonin: A review. Food Chem. 2012, 130, 797–813. [Google Scholar] [CrossRef]
- Granner, T.; Maloney, S.; Antecka, E.; Correa, J.A.; Burnier, M.N. 3,4 Dihydroxyphenyl Ethanol Reduces Secretion of Angiogenin in Human Retinal Pigment Epithelial Cells. Br. J. Ophthalmol. 2013, 97, 371–374. [Google Scholar] [CrossRef]
- Zou, X.; Feng, Z.; Li, Y.; Wang, Y.; Wertz, K.; Weber, P.; Fu, Y.; Liu, J. Stimulation of GSH synthesis to prevent oxidative stress-induced apoptosis by hydroxytyrosol in human retinal pigment epithelial cells: Activation of Nrf2 and JNK-p62/SQSTM1 pathways. J. Nutr. Biochem. 2012, 23, 994–1006. [Google Scholar] [CrossRef]
- González-Correa, J.A.; Rodríguez-Pérez, M.D.; Márquez-Estrada, L.; López-Villodres, J.A.; Reyes, J.J.; Rodriguez-Gutierrez, G.; Fernández-Bolaños, J.; De La Cruz, J.P. Neuroprotective Effect of Hydroxytyrosol in Experimental Diabetic Retinopathy: Relationship with Cardiovascular Biomarkers. J. Agric. Food Chem. 2018, 66, 637–644. [Google Scholar] [CrossRef]
- Di Mauro, M.D.; Fava, G.; Spampinato, M.; Aleo, D.; Melilli, B.; Saita, M.G.; Centonze, G.; Maggiore, R.; D’antona, N. Polyphenolic fraction from olive mill wastewater: Scale-up and in vitro studies for ophthalmic nutraceutical applications. Antioxidants 2019, 8, 462. [Google Scholar] [CrossRef] [PubMed]
- Katsinas, N.; Rodríguez-Rojo, S.; Enríquez-de-Salamanca, A. Olive Pomace Phenolic Compounds and Extracts Can Inhibit Inflammatory- and Oxidative-Related Diseases of Human Ocular Surface Epithelium. Antioxidants 2021, 10, 1150. [Google Scholar] [CrossRef] [PubMed]
- Dursun, D.; Wang, M.; Monroy, D.; Li, D.Q.; Lokeshwar, B.L.; Stern, M.E.; Pflugfelder, S.C. A mouse model of keratoconjunctivitis sicca. Investig. Ophthalmol. Vis. Sci. 2002, 43, 632–638. [Google Scholar]
- Gehlsen, U.; Braun, T.; Notara, M.; Krösser, S.; Steven, P. A semifluorinated alkane (F4H5) as novel carrier for cyclosporine A: A promising therapeutic and prophylactic option for topical treatment of dry eye. Graefe’s Arch. Clin. Exp. Ophthalmol. 2017, 255, 767–775. [Google Scholar] [CrossRef]
- Jain, S.M.; Priyadarshan, P.M. Breeding Plantation Tree Crops: Tropical Species; Springer: New York, NY, USA, 2009. [Google Scholar]
- Katsinas, N.; Enríquez-de-Salamanca, A.; da Silva, A.B.; Bronze, M.R.; Rodríguez-Rojo, S. Olive Pomace Phenolic Compounds Stability and Safety Evaluation: From Raw Material to Future Ophthalmic Applications. Molecules 2021, 26, 6002. [Google Scholar] [CrossRef]
- Bron, A.J.; Evans, V.E.; Smith, J.A. Grading Of Corneal and Conjunctival Staining in the Context of Other Dry Eye Tests. Cornea 2003, 22, 640–650. [Google Scholar] [CrossRef]
- Gehlsen, U.; Stary, D.; Maass, M.; Riesner, K.; Musial, G.; Stern, M.E.; Penack, O.; Steven, P. Ocular Graft-versus-Host Disease in a Chemotherapy-Based Minor-Mismatch Mouse Model Features Corneal (Lymph-) Angiogenesis. Int. J. Mol. Sci. 2021, 22, 6191. [Google Scholar] [CrossRef]
- Mirzaei, H.; Salehi, H.; Oskuee, R.K.; Mohammadpour, A.; Mirzaei, H.R.; Sharifi, M.R.; Salarinia, R.; Darani, H.Y.; Mokhtari, M.; Masoudifar, A.; et al. The therapeutic potential of human adipose-derived mesenchymal stem cells producing CXCL10 in a mouse melanoma lung metastasis model. Cancer Lett. 2018, 419, 30–39. [Google Scholar] [CrossRef]
- Salabarria, A.-C.; Koch, M.; Schönberg, A.; Zinser, E.; Hos, D.; Hamdorf, M.; Imhof, T.; Braun, G.; Cursiefen, C.; Bock, F. Topical VEGF-C/D Inhibition Prevents Lymphatic Vessel Ingrowth into Cornea but Does Not Improve Corneal Graft Survival. J. Clin. Med. 2020, 9, 1270. [Google Scholar] [CrossRef]
- Marković, A.K.; Torić, J.; Barbarić, M.; Brala, C.J. Hydroxytyrosol, Tyrosol and Derivatives and Their Potential Effects on Human Health. Molecules 2019, 24, 2001. [Google Scholar] [CrossRef]
- Alvarez-Laderas, I.; Ramos, T.L.; Medrano, M.; Caracuel-García, R.; Barbado, M.V.; Sánchez-Hidalgo, M.; Zamora, R.; Alarcón-de-la-Lastra, C.; Hidalgo, F.J.; Piruat, J.I.; et al. Polyphenolic Extract (PE) from Olive Oil Exerts a Potent Immunomodulatory Effect and Prevents Graft-versus-Host Disease in a Mouse Model. Biol. Blood Marrow Transplant. 2020, 26, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Dana, R. Autoimmunity in dry eye disease—An updated review of evidence on effector and memory Th17 cells in disease pathogenicity. Autoimmun. Rev. 2021, 20, 102933. [Google Scholar] [CrossRef] [PubMed]
- Pflugfelder, S.C.; Corrales, R.M.; de Paiva, C.S. T helper cytokines in dry eye disease. Exp. Eye Res. 2013, 117, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Craig, J.P.; Nelson, J.D.; Azar, D.T.; Belmonte, C.; Bron, A.J.; Chauhan, S.K.; de Paiva, C.S.; Gomes, J.A.P.; Hammitt, K.M.; Jones, L.; et al. TFOS DEWS II Report Executive Summary. Ocul. Surf. 2017, 15, 802–812. [Google Scholar] [CrossRef]
- De Paiva, C.S.; Villarreal, A.L.; Corrales, R.M.; Rahman, H.T.; Chang, V.Y.; Farley, W.J.; Stern, M.E.; Niederkorn, J.Y.; Li, D.-Q.; Pflugfelder, S.C. Dry Eye–Induced Conjunctival Epithelial Squamous Metaplasia Is Modulated by Interferon-γ. Investig. Ophthalmol. Vis. Sci. 2007, 48, 2553–2560. [Google Scholar] [CrossRef]
- Solomon, A.; Dursun, D.; Liu, Z.; Xie, Y.; Macri, A.; Pflugfelder, S.C. Pro- and anti-inflammatory forms of interleukin-1 in the tear fluid and conjunctiva of patients with dry-eye disease. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2283–2292. [Google Scholar]
- Stern, M.E.; Gao, J.; Schwalb, T.A.; Ngo, M.; Tieu, D.D.; Chan, C.C.; Reis, B.L.; Whitcup, S.M.; Thompson, D.; Smith, J.A. Conjunctival T-cell subpopulations in Sjögren’s and non-Sjögren’s patients with dry eye. Investig. Ophthalmol. Vis. Sci. 2002, 43, 2609–2614. [Google Scholar]
- Lee-Huang, S.; Huang, P.L.; Zhang, D.; Lee, J.W.; Bao, J.; Sun, Y.; Chang, Y.-T.; Zhang, J.; Huang, P.L. Discovery of small-molecule HIV-1 fusion and integrase inhibitors oleuropein and hydroxytyrosol: Part I. Integrase inhibition. Biochem. Biophys. Res. Commun. 2007, 354, 872. [Google Scholar] [CrossRef]
- Ford, C.T.; Richardson, S.; McArdle, F.; Lotito, S.B.; Crozier, A.; McArdle, A.; Jackson, M.J. Identification of (poly)phenol treatments that modulate the release of pro-inflammatory cytokines by human lymphocytes. Br. J. Nutr. 2016, 115, 1699–1710. [Google Scholar] [CrossRef][Green Version]
- Zou, T.; Yang, Y.; Xia, F.; Huang, A.; Gao, X.; Fang, D.; Xiong, S.; Zhang, J. Resveratrol Inhibits CD4+ T cell activation by enhancing the expression and activity of Sirt1. PLoS ONE 2013, 8, e75139. [Google Scholar] [CrossRef]
- Sun, Y.; Wu, X.X.; Yin, Y.; Gong, F.Y.; Shen, Y.; Cai, T.T.; Zhou, X.B.; Wu, X.F.; Xu, Q. Novel immunomodulatory properties of cirsilineol through selective inhibition of IFN-γ signaling in a murine model of inflammatory bowel disease. Biochem. Pharmacol. 2010, 79, 229–238. [Google Scholar] [CrossRef]
- Yamanaka, D.; Tamiya, Y.; Motoi, M.; Ishibashi, K.; Miura, N.N.; Adachi, Y.; Ohno, N. The Effect of Enzymatically Polymerised Polyphenols on CD4 Binding and Cytokine Production in Murine Splenocytes. PLoS ONE 2012, 7, e36025. [Google Scholar] [CrossRef] [PubMed]
- Park, B.; Lee, I.S.; Hyun, S.W.; Jo, K.; Lee, T.G.; Kim, J.S.; Kim, C.S. The protective effect of Polygonum cuspidatum (PCE) aqueous extract in a dry eye model. Nutrients 2018, 10, 1550. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.J.; Lai, J.Y. Epigallocatechin Gallate-Loaded Gelatin-g-Poly (N-Isopropylacrylamide) as a New Ophthalmic Pharmaceutical Formulation for Topical Use in the Treatment of Dry Eye Syndrome. Sci. Rep. 2017, 7, 9380. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.J.; Luo, L.J.; Harroun, S.G.; Wei, S.C.; Unnikrishnan, B.; Chang, H.T.; Huang, Y.F.; Lai, J.Y.; Huang, C.C. Synergistically dual-functional nano eye-drops for simultaneous anti-inflammatory and anti-oxidative treatment of dry eye disease. Nanoscale 2019, 11, 5580–5594. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.; Lee, J.B.; Cui, L.; Li, Y.; Li, Z.; Choi, J.S.; Lee, H.S.; Yoon, K.C. Therapeutic Efficacy of Topically Applied Antioxidant Medicinal Plant Extracts in a Mouse Model of Experimental Dry Eye. Oxid. Med. Cell. Longev. 2016, 2016, 4727415. [Google Scholar] [CrossRef]
- Cui, L.; Lee, H.S.; Li, Y.; Choi, J.H.; Yun, J.J.; Jung, J.E.; Choi, W.; Yoon, K.C. Experimental and clinical applications of chamaecyparis obtusa extracts in dry eye disease. Oxid. Med. Cell. Longev. 2017, 2017, 4523673. [Google Scholar] [CrossRef]
- Saito, K.; Mori, S.; Date, F.; Hong, G. Epigallocatechin gallate stimulates the neuroreactive salivary secretomotor system in autoimmune sialadenitis of MRL-Faslpr mice via activation of cAMP-dependent protein kinase A and inactivation of nuclear factor κb. Autoimmunity 2015, 48, 379–388. [Google Scholar] [CrossRef]
- Oh, H.N.; Kim, C.E.; Lee, J.H.; Yang, J.W. Effects of Quercetin in a Mouse Model of Experimental Dry Eye. Cornea 2015, 34, 1130–1136. [Google Scholar] [CrossRef]
- de Paiva, C.; Pflugfelder, S.; Ng, S.; Akpek, E. Topical cyclosporine A therapy for dry eye syndrome. Cochrane Database Syst. Rev. 2019, 9, CD010051. [Google Scholar] [CrossRef]
- Gasteiger, G.; Ataide, M.; Kastenmüller, W. Lymph node—An organ for T-cell activation and pathogen defense. Immunol. Rev. 2016, 271, 200–220. [Google Scholar] [CrossRef] [PubMed]
- El Annan, J.; Chauhan, S.K.; Ecoiffier, T.; Zhang, Q.; Saban, D.R.; Dana, R. Characterization of Effector T Cells in Dry Eye Disease. Investig. Ophthalmol. Vis. Sci. 2009, 50, 3802–3807. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.C.; Park, C.S.; You, I.C.; Choi, H.J.; Lee, K.H.; Im, S.K.; Park, H.Y.; Pflugfelder, S.C. Expression of CXCL9, -10, -11, and CXCR3 in the tear film and ocular surface of patients with dry eye syndrome. Investig. Ophthalmol. Vis. Sci. 2010, 51, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Corrales, R.M.; Villarreal, A.; Farley, W.; Stern, M.E.; Li, D.Q.; Pflugfelder, S.C. Strain-related cytokine profiles on the murine ocular surface in response to desiccating stress. Cornea 2007, 26, 579–584. [Google Scholar] [CrossRef]
- Yoon, K.C.; De Paiva, C.S.; Qi, H.; Chen, Z.; Farley, W.J.; Li, D.Q.; Pflugfelder, S.C. Expression of Th-1 chemokines and chemokine receptors on the ocular surface of C57BL/6 mice: Effects of desiccating stress. Investig. Ophthalmol. Vis. Sci. 2007, 48, 2561–2569. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.Y.; Wang, M.C.; Chen, Z.Y.; Chiu, W.-Y.; Chen, K.-H.; Lin, I.-C.; Vivian Yang, W.-C.; Wu, C.-C.; Tseng, C.-L. Gelatin–epigallocatechin gallate nanoparticles with hyaluronic acid decoration as eye drops can treat rabbit dry-eye syndrome effectively via inflammatory relief. Int. J. Nanomed. 2018, 13, 7251–7273. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.J.; Kim, J.M.; Lee, M.J.; Sohn, Y.S.; Kang, K.K.; Yoo, M. The therapeutic effect of DA-6034 on ocular inflammation via suppression of MMP-9 and inflammatory cytokines and activation of the MAPK signaling pathway in an experimental dry eye model. Curr. Eye Res. 2010, 35, 165–175. [Google Scholar] [CrossRef]
- Lee, H.; Shim, W.; Kim, C.E.; Choi, S.Y.; Lee, H.; Yang, J. Therapeutic efficacy of nanocomplex of poly (ethylene glycol) and catechin for dry eye disease in a mouse model. Investig. Ophthalmol. Vis. Sci. 2017, 58, 1682–1691. [Google Scholar] [CrossRef]
- Lee, H.S.; Choi, J.H.; Cui, L.; Li, Y.; Yang, J.M.; Yun, J.J.; Jung, J.E.; Choi, W.; Yoon, K.C. Anti-inflammatory and antioxidative effects of Camellia japonica on human corneal epithelial cells and experimental dry eye: In vivo and in vitro study. Investig. Ophthalmol. Vis. Sci. 2017, 58, 1196–1207. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody (Clone) | Target | Conjugation | Manufacturer | Catalog No. | Concentration (mg/mL) |
|---|---|---|---|---|---|
| Anti-mouse CD3 (17A2) | CD3+ T cells | allophycocyanin-cyanine 7 (APC-Cy7) | Biolegend (San Diego, CA, USA) | 100221 | 0.10 |
| Anti-mouse CD4 (GK1.5) | CD4+ T cells | fluorescein isothiocyanate (FITC) | 100405 | 0.125 | |
| Anti-mouse CD8a (53–6.7) | CD8+ T cells | APC | 100711 | 0.05 |
| mRNA | Sequence | Annealing Temperature |
|---|---|---|
| HPRT | F: 5′-TTGGATACAGGCCAGACTTTGTTG-3′ R: 5′-GATTCAACTTGCGCTCATCTTAGGC-3′ | 60 °C |
| IP-10 | F: 5′-ATATACGCGTTGACATTGATTATTGACTAG-3′ R: 5′-ATTGCTAG-CAGCTGGTTCTTTCCGCCTC-3′ | 60 °C |
| TNF-α | F: 5′- AGGACTCAAATGGGCTTTCC-3′ R: 5′-CAGAGGCAACCTGACCACTC-3′ | 63 °C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katsinas, N.; Gehlsen, U.; García-Posadas, L.; Rodríguez-Rojo, S.; Steven, P.; González-García, M.J.; Enríquez-de-Salamanca, A. Olive Pomace Phenolic Compounds: From an Agro-Industrial By-Product to a Promising Ocular Surface Protection for Dry Eye Disease. J. Clin. Med. 2022, 11, 4703. https://doi.org/10.3390/jcm11164703
Katsinas N, Gehlsen U, García-Posadas L, Rodríguez-Rojo S, Steven P, González-García MJ, Enríquez-de-Salamanca A. Olive Pomace Phenolic Compounds: From an Agro-Industrial By-Product to a Promising Ocular Surface Protection for Dry Eye Disease. Journal of Clinical Medicine. 2022; 11(16):4703. https://doi.org/10.3390/jcm11164703
Chicago/Turabian StyleKatsinas, Nikolaos, Uta Gehlsen, Laura García-Posadas, Soraya Rodríguez-Rojo, Philipp Steven, María J. González-García, and Amalia Enríquez-de-Salamanca. 2022. "Olive Pomace Phenolic Compounds: From an Agro-Industrial By-Product to a Promising Ocular Surface Protection for Dry Eye Disease" Journal of Clinical Medicine 11, no. 16: 4703. https://doi.org/10.3390/jcm11164703
APA StyleKatsinas, N., Gehlsen, U., García-Posadas, L., Rodríguez-Rojo, S., Steven, P., González-García, M. J., & Enríquez-de-Salamanca, A. (2022). Olive Pomace Phenolic Compounds: From an Agro-Industrial By-Product to a Promising Ocular Surface Protection for Dry Eye Disease. Journal of Clinical Medicine, 11(16), 4703. https://doi.org/10.3390/jcm11164703