
Evaluation of Circulating Chitotriosidase Activity in Children with Obesity

 ,
,  ,
,  , , ,
, , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Clinical Variables
2.2. Plasma CHIT1 Assay
2.3. CHIT1 Genotyping
2.4. Statistical Analysis
3. Results
3.1. Description of the Study Sample
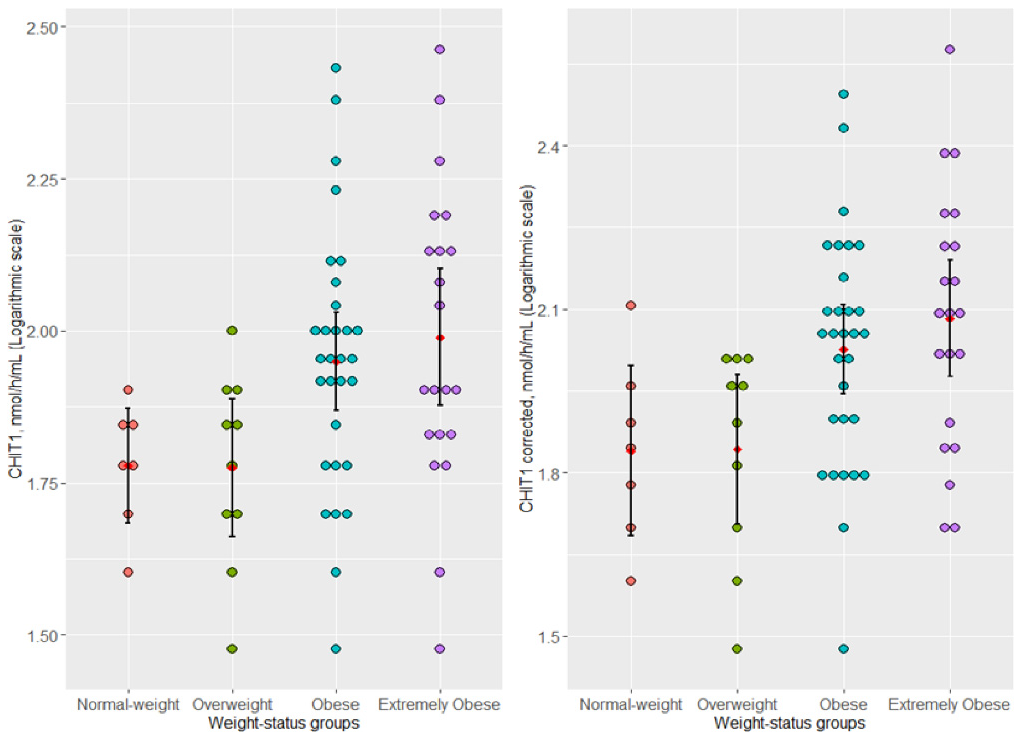
3.2. Association of Plasma-Uncorrected (CHIT1) and -Corrected (CHIT1-Corrected) Chitotriosidase Activity and Weight-Status Subgroups
3.3. Genotype Distribution of dup24 and G102S Gene Polymorphisms in Weight-Status Subgroups
3.4. Association of Plasma CHIT1 Activity with dup24 and G102S Polymorphisms within Weight-Status Subgroups
3.5. The Effect of the dup24, G102S Polymorphism, and BMI-for-Age z Score upon Variation of CHIT1 Plasma Activity in Children with Overweight, Obesity, and Extreme Obesity
4. Discussion
4.1. Association of Plasma Chitotriosidase Activity and Weight-Status Subgroups
4.2. Dup24 and G102S Gene Polymorphisms and CHIT1 Activity
4.3. Multivariable Analysis with CHIT1 Plasma Activity as the Dependent Variable
4.4. Study Limitations and Further Research
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chatzigeorgiou, A.; Karalis, K.P.; Bornstein, S.R.; Chavakis, T. Lymphocytes in obesity-related adipose tissue inflammation. Diabetologia 2012, 55, 2583–2592. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, S.; Manabe, I.; Nagasaki, M.; Eto, K.; Yamashita, H.; Ohsugi, M.; Otsu, M.; Hara, K.; Ueki, K.; Sugiura, S.; et al. CD8+ effector T cells contribute to macrophage recruitment and adipose tissue inflammation in obesity. Nat. Med. 2009, 15, 914–920. [Google Scholar] [CrossRef]
- McLaughlin, T.; Ackerman, S.E.; Shen, L.; Engleman, E. Role of innate and adaptive immunity in obesity-associated metabolic disease. J. Clin. Investig. 2017, 127, 5–13. [Google Scholar] [CrossRef]
- Zabarsky, G.; Beek, C.; Hagman, E.; Pierpont, B.; Caprio, S.; Weiss, R. Impact of Severe Obesity on Cardiovascular Risk Factors in Youth. J. Pediatr. 2018, 192, 105–114. [Google Scholar] [CrossRef] [PubMed]
- DeBoer, M.D. Obesity, systemic inflammation, and increased risk for cardiovascular disease and diabetes among adolescents: A need for screening tools to target interventions. Nutrition 2013, 29, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Wilson, P.W.F.; D’Agostino, R.B.; Sullivan, L.; Parise, H.; Kannel, W.B. Overweight and obesity as determinants of cardiovascular risk: The Framingham experience. Arch. Intern. Med. 2002, 162, 1867–1872. [Google Scholar] [CrossRef] [PubMed]
- Juonala, M.; Magnussen, C.G.; Berenson, G.S.; Venn, A.; Burns, T.L.; Sabin, M.A.; Srinivasan, S.R.; Daniels, S.R.; Davis, P.H.; Chen, W.; et al. Childhood adiposity, adult adiposity, and cardiovascular risk factors. N. Engl. J. Med. 2011, 365, 1876–1885. [Google Scholar] [CrossRef]
- Lo, K.; Wong, M.; Khalechelvam, P.; Tam, W. Waist-to-height ratio, body mass index and waist circumference for screening paediatric cardio-metabolic risk factors: A meta-analysis. Obes. Rev. 2016, 17, 1258–1275. [Google Scholar] [CrossRef]
- Sardinha, L.B.; Santos, D.A.; Silva, A.M.; Grøntved, A.; Andersen, L.B.; Ekelund, U. A Comparison between BMI, Waist Circumference, and Waist-To-Height Ratio for Identifying Cardio-Metabolic Risk in Children and Adolescents. PLoS ONE 2016, 11, e0149351. [Google Scholar] [CrossRef]
- Weiss, R.; Dziura, J.; Burgert, T.S.; Tamborlane, W.V.; Taksali, S.E.; Yeckel, C.W.; Allen, K.; Lopes, M.; Savoye, M.; Morrison, J.; et al. Obesity and the metabolic syndrome in children and adolescents. N. Engl. J. Med. 2004, 350, 2362–2374. [Google Scholar] [CrossRef]
- Allender, S.; Orellana, L.; Crooks, N.; Bolton, K.A.; Fraser, P.; Brown, A.D.; Le, H.; Lowe, J.; de la Haye, K.; Millar, L.; et al. Four-Year Behavioral, Health-Related Quality of Life, and BMI Outcomes from a Cluster Randomized Whole of Systems Trial of Prevention Strategies for Childhood Obesity. Obesity 2021, 29, 1022–1035. [Google Scholar] [CrossRef] [PubMed]
- Singer, K.; Lumeng, C.N. The initiation of metabolic inflammation in childhood obesity. J. Clin. Investig. 2017, 127, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Michalsky, M.P.; Inge, T.H.; Simmons, M.; Jenkins, T.M.; Buncher, R.; Helmrath, M.; Brandt, M.L.; Harmon, C.M.; Courcoulas, A.; Chen, M.; et al. Teen-LABS Consortium. Cardiovascular Risk Factors in Severely Obese Adolescents: The Teen Longitudinal Assessment of Bariatric Surgery (Teen-LABS) Study. JAMA Pediatr. 2015, 169, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Tans, R.; van Diepen, J.; Bijlsma, S.; Verschuren, L.; Suppers, A.; Stienstra, R.; Wevers, R.A.; Tack, C.J.; Gloerich, J.; van Gool, A.J. Evaluation of chitotriosidase as a biomarker for adipose tissue inflammation in overweight individuals and type 2 diabetic patients. Int. J. Obes. 2019, 43, 1712–1723. [Google Scholar] [CrossRef]
- Di Rosa, M.; Malaguarnera, G.; De Gregorio, C.; Drago, F.; Malaguarnera, L. Evaluation of CHI3L-1 and CHIT-1 expression in differentiated and polarized macrophages. Inflammation 2013, 36, 482–492. [Google Scholar] [CrossRef]
- Raskovalova, T.; Deegan, P.B.; Mistry, P.K.; Pavlova, E.; Yang, R.; Zimran, A.; Berger, J.; Bourgne, C.; Pereira, B.; Labarère, J.; et al. Accuracy of chitotriosidase activity and CCL18 concentration in assessing type I Gaucher disease severity. A systematic review with meta-analysis of individual participant data. Haematologica 2020, 106, 437–445. [Google Scholar] [CrossRef]
- Guo, Y.; He, W.; Boer, A.M.; Wevers, R.A.; de Bruijn, A.M.; Groener, J.E.; Hollak, C.E.; Aerts, J.M.; Galjaard, H.; van Diggelen, O.P. Elevated plasma chitotriosidase activity in various lysosomal storage disorders. J. Inherit. Metab. Dis. 1995, 18, 717–722. [Google Scholar] [CrossRef]
- Elmonem, M.A.; van den Heuvel, L.P.; Levtchenko, E.N. Immunomodulatory Effects of Chitotriosidase Enzyme. Enzym. Res. 2016, 2016, 2682680. [Google Scholar] [CrossRef]
- Ruggiero, A.D.; Key, C.C.; Kavanagh, K. Adipose tissue macrophage polarization in healthy and unhealthy obesity. Front. Nutr. 2021, 7, 625331. [Google Scholar] [CrossRef]
- Dahlman, I.; Kaaman, M.; Olsson, T.; Tan, G.D.; Bickerton, A.S.T.; Wåhlén, K.; Andersson, J.; Nordström, E.A.; Blomqvist, L.; Sjögren, A.; et al. A Unique Role of Monocyte Chemoattractant Protein 1 among Chemokines in Adipose Tissue of Obese Subjects. J. Clin. Endocrinol. Metab. 2005, 90, 5834–5840. [Google Scholar] [CrossRef]
- Olefsky, J.M.; Glass, C.K. Macrophages, inflammation, and insulin resistance. Annu. Rev. Physiol. 2010, 72, 219–246. [Google Scholar] [CrossRef] [PubMed]
- Di Rosa, M.; Mangano, K.; De Gregorio, C.; Nicoletti, F.; Malaguarnera, L. Association of chitotriosidase genotype with the development of non-alcoholic fatty liver disease. Hepatol. Res. 2013, 43, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Kabaroğlu, C.; Onur, E.; Barutçuoğlu, B.; Özhan, B.; Erdinç, S.; Var, A.; Bayindir, O.; Ersoy, B. Inflammatory marker levels in obese adolescents with glucose intolerance: Increased chitotriosidase activity. Clin. Biochem. 2012, 45, 281–284. [Google Scholar] [CrossRef] [PubMed]
- Tamanaha, P.; D’Almeida, V.; Calegare, B.F.A.; Tomita, L.Y.; Bittencourt, L.R.A.; Tufik, S. 24 bp duplication of CHIT1 gene and determinants of human chitotriosidase activity among participants of EPISONO, a population-based cross-sectional study, São Paulo, Brazil. Clin. Biochem. 2013, 46, 1084–1088. [Google Scholar] [CrossRef] [PubMed]
- Azarsız, E.; Karaca, N.; Levent, E.; Kutukculer, N.; Sozmen, E. Chitotriosidase enzyme activity: Is this a possible chronic inflammation marker in children with common variable immunodeficiency and early atherosclerosis? Ann. Clin. Biochem. 2017, 54, 636–643. [Google Scholar] [CrossRef]
- Elmonem, M.A.; Ramadan, D.I.; Issac, M.S.M.; Selim, L.A.; Elkateb, S.M. Blood spot versus plasma chitotriosidase: A systematic clinical comparison. Clin. Biochem. 2014, 47, 38–43. [Google Scholar] [CrossRef]
- Grace, M.E.; Balwani, M.; Nazarenko, I.; Prakash-Cheng, A.; Desnick, R.J. Type 1 Gaucher disease: Null and hypomorphic novel chitotriosidase mutations-implications for diagnosis and therapeutic monitoring. Hum. Mutat. 2007, 28, 866–873. [Google Scholar] [CrossRef]
- Csongrádi, A.; Altorjay, I.T.; Fülöp, G.Á.; Enyedi, A.; Enyedi, E.E.; Hajnal, P.; Takács, I.; Tóth, A.; Papp, Z.; Fagyas, M. Chitotriosidase gene polymorphisms and mutations limit the determination of chitotriosidase expression in sarcoidosis. Clin. Chim. Acta 2021, 513, 50–56. [Google Scholar] [CrossRef]
- Fusetti, F.; von Moeller, H.; Houston, D.; Rozeboom, H.J.; Dijkstra, B.W.; Boot, R.G.; Aerts, J.M.; van Aalten, D.M. Structure of human chitotriosidase. Implications for specific inhibitor design and function of mammalian chitinase-like lectins. J. Biol. Chem. 2002, 277, 25537–25544. [Google Scholar] [CrossRef]
- Boot, R.G.; Renkema, G.H.; Verhoek, M.; Strijland, A.; Bliek, J.; de Meulemeester, T.M.; Mannens, M.M.A.M.; Aerts, J.M.F.G. The human chitotriosidase gene. Nature of inherited enzyme deficiency. J. Biol. Chem. 1998, 273, 25680–25685. [Google Scholar] [CrossRef]
- Duarte, A.J.; Ribeiro, D.; Amaral, O. CHIT1 genetic defects in the Portuguese population. Blood Cells Mol. Dis. 2013, 50, 50–52. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.; Waalen, J.; Crain, K.; Smargon, A.; Beutler, E. Human Chitotriosidase Polymorphisms G354R and A442V Associated with Reduced Enzyme Activity. Blood Cells Mol. Dis. 2007, 39, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, B.; Ghauharali-van der Vlugt, K.; Helmond, M.T.J.; Out, J.M.M.; Donker-Koopman, W.E.; Groener, J.E.M.; Boot, R.G.; Renkema, G.H.; van der Marel, G.A.; van Boom, J.H.; et al. Transglycosidase activity of chitotriosidase: Improved enzymatic assay for the human macrophage chitinase. J. Biol. Chem. 2003, 278, 40911–40916. [Google Scholar] [CrossRef]
- Bussink, A.P.; Verhoek, M.; Vreede, J.; Ghauharali-van der Vlugt, K.; Donker-Koopman, W.E.; Sprenger, R.R.; Hollak, C.E.; Aerts, J.M.; Boot, R.G. Common G102S polymorphism in chitotriosidase differentially affects activity towards 4-methylumbelliferyl substrates. FEBS J. 2009, 276, 5678–5688. [Google Scholar] [CrossRef]
- Schoonhoven, A.; Rudensky, B.; Elstein, D.; Zimran, A.; Hollak, C.E.M.; Groener, J.E.; Aerts, J.M. Monitoring of Gaucher patients with a novel chitotriosidase assay. Clin. Chim. Acta 2007, 381, 136–139. [Google Scholar] [CrossRef] [PubMed]
- Chung, W.K. An overview of mongenic and syndromic obesities in humans. Pediatr. Blood Cancer 2012, 58, 122–128. [Google Scholar] [CrossRef]
- Schmitz, N.A.; Thakare, R.P.; Chung, C.S.; Lee, C.M.; Elias, J.A.; Lee, C.G.; LeBlanc, B.W. Chitotriosidase Activity Is Counterproductive in a Mouse Model of Systemic Candidiasis. Front. Immunol. 2021, 16, e626798. [Google Scholar] [CrossRef]
- Labadaridis, I.; Dimitriou, E.; Theodorakis, M.; Kafalidis, G.; Velegraki, A.; Michelakakis, H. Chitotriosidase in neonates with fungal and bacterial infections. Arch. Dis. Child. Fetal Neonatal Ed. 2005, 90, F531–F532. [Google Scholar] [CrossRef]
- Sharma, L.; Amick, A.K.; Vasudevan, S.; Lee, S.W.; Marion, C.R.; Liu, W.; Brady, V.; Losier, A.; Bermejo, S.D.; Britto, C.J.; et al. Regulation and Role of Chitotriosidase during Lung Infection with Klebsiella pneumoniae. J. Immunol. 2018, 201, 615–626. [Google Scholar] [CrossRef]
- Barone, R.; Simporè, J.; Malaguarnera, L.; Pignatelli, S.; Musumeci, S. Plasma chitotriosidase activity in acute Plasmodium falciparum malaria. J. Trop. Pediatr. 2003, 49, 63–64. [Google Scholar] [CrossRef][Green Version]
- Bargagli, E.; Bennett, D.; Maggiorelli, C.; Di Sipio, P.; Margollicci, M.; Bianchi, N.; Rottoli, P. Human chitotriosidase: A sensitive biomarker of sarcoidosis. J. Clin. Immunol. 2013, 33, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Brunner, J.K.; Scholl-Bürgi, S.; Hössinger, D.; Wondrak, P.; Prelog, M.; Zimmerhackl, L.B. Chitotriosidase activity in juvenile idiopathic arthritis. Rheumatol. Int. 2008, 28, 949–950. [Google Scholar] [CrossRef]
- Yavuz, F.; Kesikburun, B.; Öztürk, Ö.; Güzelküçük, Ü. Serum chitotriosidase and neopterin levels in patients with ankylosing spondylitis. Ther. Adv. Musculoskelet. Dis. 2019, 11, e1759720X19832321. [Google Scholar] [CrossRef] [PubMed]
- Mazur, M.; Zielińska, A.; Grzybowski, M.M.; Olczak, J.; Fichna, J. Chitinases and Chitinase-Like Proteins as Therapeutic Targets in Inflammatory Diseases, with a Special Focus on Inflammatory Bowel Diseases. Int. J. Mol. Sci. 2021, 22, 6966. [Google Scholar] [CrossRef]
- Majewski, S.; Szewczyk, K.; Jerczyńska, H.; Miłkowska-Dymanowska, J.; Białas, A.J.; Gwadera, Ł.; Piotrowski, W.J. Longitudinal and Comparative Measures of Serum Chitotriosidase and YKL-40 in Patients With Idiopathic Pulmonary Fibrosis. Front. Immunol. 2022, 13, 760776. [Google Scholar] [CrossRef] [PubMed]
- WHO Child Growth Standards: Length/Height-for-Age, Weight-for-Age, Weight-for-Length, Weight-for-Height and Body Mass Index-for-Age: Methods and Development. Available online: https://apps.who.int/iris/handle/10665/43413 (accessed on 3 June 2022).
- Marshall, W.A.; Tanner, J.M. Variations in pattern of pubertal changes in girls. Arch. Dis. Child. 1969, 44, 291–303. [Google Scholar] [CrossRef]
- Marshall, W.A.; Tanner, J.M. Variations in the Pattern of Pubertal Changes in Boys. Arch. Dis. Child. 1970, 45, 13–23. [Google Scholar] [CrossRef]
- Hollak, C.E.; van Weely, S.; van Oers, M.H.; Aerts, J.M. Marked elevation of plasma chitotriosidase activity. A novel hallmark of Gaucher disease. J. Clin. Investig. 1994, 93, 1288–1292. [Google Scholar] [CrossRef]
- Wizard® Genomic DNA Purification Kit Technical Manual. Promega Romania. Available online: https://www.promega.ro/resources/protocols/technical-manuals/0/wizard-genomic-dna-purification-kit-protocol/ (accessed on 3 June 2022).
- UCSC In-Silico PCR. Available online: https://genome.ucsc.edu/cgi-bin/hgPcr (accessed on 3 June 2022).
- Okonechnikov, K.; Golosova, O.; Fursov, M.; UGENE Team. Unipro UGENE: A Unified Bioinformatics Toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria; Available online: https://www.R-project.org/ (accessed on 3 September 2021).
- Moreno, V.; Gonzalez, J.R. Package ‘SNPassoc’ Version 2.0-11. 2021. Available online: https://cran.r-project.org/web/packages/SNPassoc/SNPassoc.pdf (accessed on 10 November 2021).
- Lotrean, L.M.; Popa, I.; Florea, M.; Lazea, C.; Stanescu, A.M.A.; Lencu, C. Actual Weight, Perceived Weight and Desired Weight of Romanian School Children by Parents and Children. Medicina 2021, 57, 333. [Google Scholar] [CrossRef]
- Chirita-Emandi, A.; Barbu, C.G.; Cinteza, E.E.; Chesaru, B.I.; Gafencu, M.; Mocanu, V.; Pascanu, I.M.; Tatar, S.A.; Balgradean, M.; Dobre, M.; et al. Overweight and Underweight Prevalence Trends in Children from Romania-Pooled Analysis of Cross-Sectional Studies between 2006 and 2015. Obes. Facts 2016, 9, 206–220. [Google Scholar] [CrossRef]
- Di Rosa, M.; Musumeci, M.; Scuto, A.; Musumeci, S.; Malaguarnera, L. Effect of interferon-gamma, interleukin-10, lipopolysaccharide and tumor necrosis factor-alpha on chitotriosidase synthesis in human macrophages. Clin. Chem. Lab. Med. 2005, 43, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Malaguarnera, L. Chitotriosidase: The yin and yang. Cell. Mol. Life Sci. 2006, 63, 3018–3029. [Google Scholar] [CrossRef] [PubMed]
- Giraldo, P.; Cenarro, A.; Alfonso, P.; Pérez-Calvo, J.I.; Rubio-Félix, D.; Giralt, M.; Pocoví, M. Chitotriosidase genotype and plasma activity in patients type 1 Gaucher’s disease and their relatives (carriers and non-carriers). Haematologica 2001, 86, 977–984. [Google Scholar]
- Cohen, E.; Margalit, I.; Shochat, T.; Goldberg, E.; Krause, I. Markers of Chronic Inflammation in Overweight and Obese Individuals and the Role of Gender: A Cross-Sectional Study of a Large Cohort. J. Inflamm. Res 2021, 14, 567–573. [Google Scholar] [CrossRef]
- Lecube, A.; Hernández, C.; Pelegrí, D.; Simó, R. Factors accounting for high ferritin levels in obesity. Int. J. Obes. 2008, 32, 1665–1669. [Google Scholar] [CrossRef]
- Grosso, S.; Margollicci, M.A.; Bargagli, E.; Buccoliero, Q.R.; Perrone, A.; Galimberti, D.; Morgese, G.; Balestri, P.; Rottoli, P. Serum levels of chitotriosidase as a marker of disease activity and clinical stage in sarcoidosis. Scand. J. Clin. Lab. Investig. 2004, 64, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Shemesh, E.; Deroma, L.; Bembi, B.; Deegan, P.; Hollak, C.; Weinreb, N.J.; Cox, T.M. Enzyme replacement and substrate reduction therapy for Gaucher disease. Cochrane Database Syst. Rev. 2015, 3, CD010324. [Google Scholar] [CrossRef]
- Chamoles, N.A.; Blanco, M.; Gaggioli, D.; Casentini, C. Gaucher and Niemann-Pick diseases-enzymatic diagnosis in dried blood spots on filter paper: Retrospective diagnoses in newborn-screening cards. Clin. Chim. Acta 2002, 317, 191–197. [Google Scholar] [CrossRef]
- Civallero, G.; Michelin, K.; de Mari, J.; Viapiana, M.; Burin, M.; Coelho, J.C.; Giugliani, R. Twelve different enzyme assays on dried-blood filter paper samples for detection of patients with selected inherited lysosomal storage diseases. Clin. Chim. Acta 2006, 372, 98–102. [Google Scholar] [CrossRef]
- Goldim, M.P.; Garcia, C.; de Castilhos, C.D.; Daitx, V.V.; Mezzalira, J.; Breier, A.C.; Cé, J.; Mello, A.; Andrade, C.V.; Sartori, N.; et al. Screening of high-risk Gaucher disease patients in Brazil using miniaturized dried blood spots and leukocyte techniques. Gene 2012, 508, 197–198. [Google Scholar] [CrossRef]
- Rodrigues, M.D.B.; de Oliveira, A.C.; Müller, K.B.; Martins, A.M.; D’Almeida, V. Chitotriosidase determination in plasma and in dried blood spots: A comparison using two different substrates in a microplate assay. Clin. Chim. Acta 2009, 406, 86–88. [Google Scholar] [CrossRef] [PubMed]
- Palmer, B.F.; Clegg, D.J. The sexual dimorphism of obesity. Mol. Cell. Endocrinol. 2015, 15, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Grove, K.L.; Fried, S.K.; Greenberg, A.S.; Xiao, X.Q.; Clegg, D.J. A microarray analysis of sexual dimorphism of adipose tissues in high-fat-diet-induced obese mice. Int. J. Obes. 2010, 34, 989–1000. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.P.; Santos, I.S.; Matijasevich, A.; Barros, A.J.D. Changes in overall and regional body fatness from childhood to early adolescence. Sci. Rep. 2019, 9, 1888. [Google Scholar] [CrossRef]
- Jura, M.; Kozak, L.P. Obesity and related consequences to ageing. AGE 2016, 38, 23. [Google Scholar] [CrossRef]
- Cutaș, A.; Drugan, C.; Roman, G.; Rusu, A.; Cătană, C.S.; Achimaș-Cadariu, A.; Drugan, T. Evaluation of Chitotriosidase and Neopterin as Biomarkers of Microvascular Complications in Patients with Type 1 Diabetes Mellitus. Diagnostics 2021, 11, 263. [Google Scholar] [CrossRef] [PubMed]
- Ciocan, R.A.; Drugan, C.; Gherman, C.D.; Cătană, C.-S.; Ciocan, A.; Drugan, T.C.; Bolboacă, S.D. Evaluation of Chitotriosidase as a Marker of Inflammatory Status in Critical Limb Ischemia. Ann. Clin. Lab. Sci. 2017, 47, 713–719. [Google Scholar]
- Mahabee-Gittens, E.M.; Merianos, A.L.; Fulkerson, P.C.; Stone, L.; Matt, G.E. The association of environmental tobacco smoke exposure and inflammatory markers in hospitalized children. Int. J. Environ. Res. Public Health 2019, 16, 4625. [Google Scholar] [CrossRef]
- Dinh, J.C.; Hosey-Cojocari, C.M.; Jones, B.L. Pediatric Clinical Endpoint and Pharmacodynamic Biomarkers: Limitations and Opportunities. Pediatr. Drugs 2020, 22, 55–71. [Google Scholar] [CrossRef]
- Panagiotakos, D.B.; Pitsavos, C.; Yannakoulia, M.; Chrysohoou, C.; Stefanadis, C. The implication of obesity and central fat on markers of chronic inflammation: The ATTICA study. Atherosclerosis 2005, 183, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Caminiti, C.; Armeno, M.; Mazza, C.S. Waist-to-height ratio as a marker of low-grade inflammation in obese children and adolescents. J. Pediatr. Endocrinol. Metab. 2016, 29, 543–551. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Forward Primer 5′→3′ Sequence | Reverse Primer 5′→3′ Sequence |
|---|---|
| GAAGAGGTAGCCAGGCTCTGG | CTGCCGTAGCGTCTGGATGAG |
| Variable | Weight-Status Subgroups | ||||
|---|---|---|---|---|---|
| Normal Weight (n1 = 7) | Overweight (n2 = 10) | Obesity (n3 = 30) | Extreme Obesity (n4 = 21) | p-Value | |
| Clinical characteristics | |||||
| Gender | 0.061 | ||||
| Girls | 3 (42.86) | 8 (80.00) | 15 (50.00) | 6 (28.57) | |
| Boys | 4 (57.14) | 2 (20.00) | 15 (50.00) | 15 (71.43) | |
| Tanner staging (a) | 0.023 # | ||||
| prepubertal | 1 (20.00) | 3 (33.33) | 4 (15.38) | 10 (55.56) | |
| pubertal | 2 (40.00) | 1 (11.11) | 19 (73.08) | 8 (44.44) | |
| postpubertal | 2 (40.00) | 5 (55.56) | 3 (11.54) | 0 (0.00) | |
| Age (years) | 12.86 ± 4.38 | 12.63 ± 2.93 | 13.64 ± 2.69 | 9.66 ± 3.00 | 0.0002 |
| Time since weight gain (b) (years) | na | 8 (3.78–11.66) {2.17–17.64} | 5.18 (3.44–7.85) {0.25–17.52} | 4.9 (3.59–6.53) {2.09–11.98} | 0.008 |
| Inflammation-related parameters | |||||
| hCRP (c) (mg/dL) | 0.28 (0.25–0.29) {0.23–0.29} | 0.24 (0.21–0.24) {0.20–0.31} | 0.27 (0.26–0.30){0.18–1.23} | 0.23 (0.21–0.29) {0.14–0.31} | 0.600 |
| Ferritin (d) (ng/mL) | 34.20 (29.9–38.5) {25.6–42.8} | 36.95 (28.0–44.3) {21.3–49.2} | 40.9 (19.83–6.48) {12.9–92.8} | 27.2 (25.1–41.4) {20.4–47.5} | 0.981 |
| WBC (e) (×103/μL) | 5.54 (4.66–5.77) {4.41–7.18} | 8.11 (7.21–8.65) {6.53–10} | 7.27 (6.52–7.91) {6.11–8.24} | 7.82 (7.2–9.18) {6–9.84} | 0.002 |
| ESR (f) (mm/h) | 7 (7–8.5) {3–16} | 16.6(12–22) {6–30} | 13.2 (6–13) {4–36} | 12 (9–15) {3–19} | 0.432 |
| Gene Variant | Normal Weight (n1 = 7) | Overweight (n2 = 10) | Obesity (n3 = 30) | Extreme Obesity (n4 = 21) | p-Value | Adjusted p-Value |
|---|---|---|---|---|---|---|
| G102S (rs2297950) | ||||||
| CC [C] | 4 (57.14) | 2 (20.0) | 10 (33.33) | 5 (23.81) | 0.428 | 0.728 |
| CT | 2 (28.57) | 8 (80.0) | 16 (53.33) | 12 (57.14) | ||
| TT | 1 (14.29) | 0 (0.0) | 4 (13.33) | 4 (19.05) | ||
| CT + TT [D] | 3 (42.86) | 8 (80.0) | 20 (66.67) | 16 (76.19) | 0.370 | 0.161 |
| C [A] | 10 (71.43) | 12 (60.0) | 36 (60.0) | 22 (52.38) | 0.641 | 0.319 |
| T | 4 (28.57) | 8 (40.0) | 24 (40.0) | 20 (47.62) | ||
| HWE (p-value) | 0.441 | 0.173 | 0.711 | 0.675 | ||
| dup24 * (rs3831317) | ||||||
| wt/wt[a] | 4 (57.14) | 7 (70.0) | 24 (80.0) | 18 (85.71) | 0.380 | 0.965 |
| wt/dup24[b] | 3 (42.86) | 3 (30.0) | 6 (20.0) | 3 (14.29) | ||
| wt allele | 11 (78.57) | 17 (85.0) | 54 (90.0) | 39 (92.86) | 0.407 | 0.967 |
| dup24 allele | 3 (21.43) | 3 (15.0) | 6 (10.0) | 3 (7.14) | ||
| HWE (p-value) | 0.471 | 0.577 | 0.543 | 0.012 | ||
| Variables | Unadjusted b [95% CI] | p | Adjusted b [95% CI] | Adjusted β [95% CI] | p |
| logCHIT1 plasma activity | |||||
| BMI z score | 0.08 [0.03; 0.13] | 0.003 * | 0.06 [0.01; 0.12] | 0.30 [0.03; 0.57] | 0.031 * |
| dup24 (wt/dup24 vs. wt/wt) | −0.22 [−0.36; −0.09] | 0.002 * | −0.25 [−0.38; −0.12] | −1.10 [−1.66, −0.55] | <0.001 * |
| G102S (CT + TT vs. CC) | −0.13 [−0.26; −0.003] | 0.045 * | −0.19 [−0.30; −0.08] | −0.83 [−1.32, −0.35] | 0.001 * |
| Age (months) | −0.0008 [−0.002; 0.0007] | 0.281 | 0.0002 [−0.001; 0.002] | 0.04 [−0.23; 0.30] | 0.787 |
| Gender (male vs. female) | 0.08 [−0.03; 0.20] | 0.159 | 0.05 [−0.06, 0.15] | 0.21 [−0.24; 0.65] | 0.361 |
| Time since weight gain (months) | −0.01 [−0.03; 0.004] | 0.142 | −0.003 [−0.02, 0.01] | −0.05 [−0.29; 0.18] | 0.649 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Țaranu, I.; Iancu, M.; Lazea, C.; Alkhzouz, C.; Răcătăianu, N.; Cătană, C.-S.; Mirea, A.-M.; Miclea, D.; Bolboacă, S.D.; Drugan, C. Evaluation of Circulating Chitotriosidase Activity in Children with Obesity. J. Clin. Med. 2022, 11, 3634. https://doi.org/10.3390/jcm11133634
Țaranu I, Iancu M, Lazea C, Alkhzouz C, Răcătăianu N, Cătană C-S, Mirea A-M, Miclea D, Bolboacă SD, Drugan C. Evaluation of Circulating Chitotriosidase Activity in Children with Obesity. Journal of Clinical Medicine. 2022; 11(13):3634. https://doi.org/10.3390/jcm11133634
Chicago/Turabian StyleȚaranu, Ioana, Mihaela Iancu, Cecilia Lazea, Camelia Alkhzouz, Nicoleta Răcătăianu, Cristina-Sorina Cătană, Andreea-Manuela Mirea, Diana Miclea, Sorana D. Bolboacă, and Cristina Drugan. 2022. "Evaluation of Circulating Chitotriosidase Activity in Children with Obesity" Journal of Clinical Medicine 11, no. 13: 3634. https://doi.org/10.3390/jcm11133634
APA StyleȚaranu, I., Iancu, M., Lazea, C., Alkhzouz, C., Răcătăianu, N., Cătană, C.-S., Mirea, A.-M., Miclea, D., Bolboacă, S. D., & Drugan, C. (2022). Evaluation of Circulating Chitotriosidase Activity in Children with Obesity. Journal of Clinical Medicine, 11(13), 3634. https://doi.org/10.3390/jcm11133634