
Correlations of Host and Bacterial Characteristics with Clinical Parameters and Survival in Staphylococcus aureus Bacteremia





Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Clinical Data
2.3. Administered Antimicrobial Therapy
2.4. Phenotypic Characterization of Isolates
2.5. Biofilm Assay
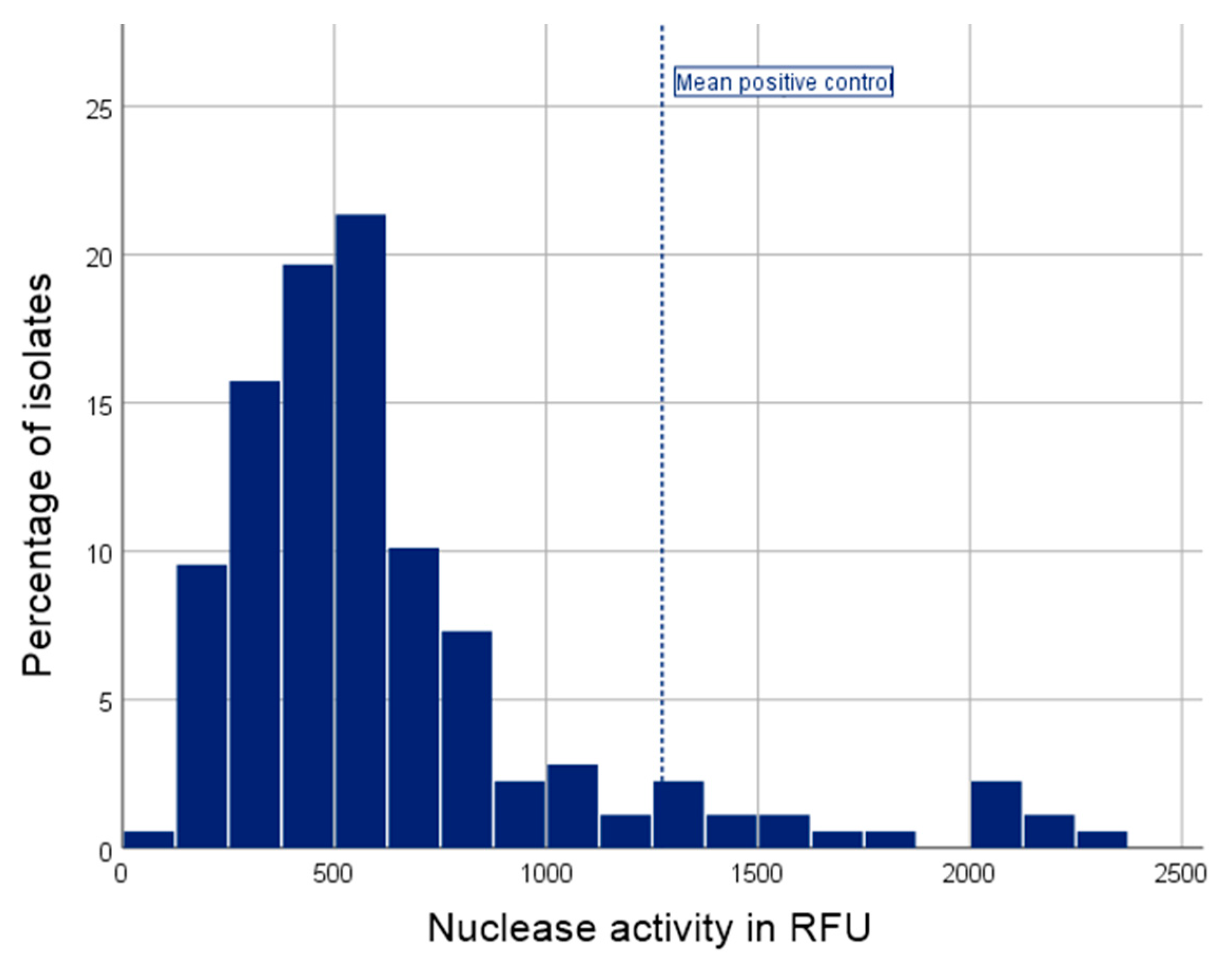
2.6. Nuclease Assay
2.7. spa Typing
2.8. Analysis of Methicillin Resistance, agr Type and Prevalence of Toxin Genes
2.9. Statistical Analyses
2.10. Ethics Statement
3. Results
3.1. Study Cohort and Clinical Data
3.2. Administered Antimicrobial Therapy
3.3. Characterization of Isolates
spa Typing
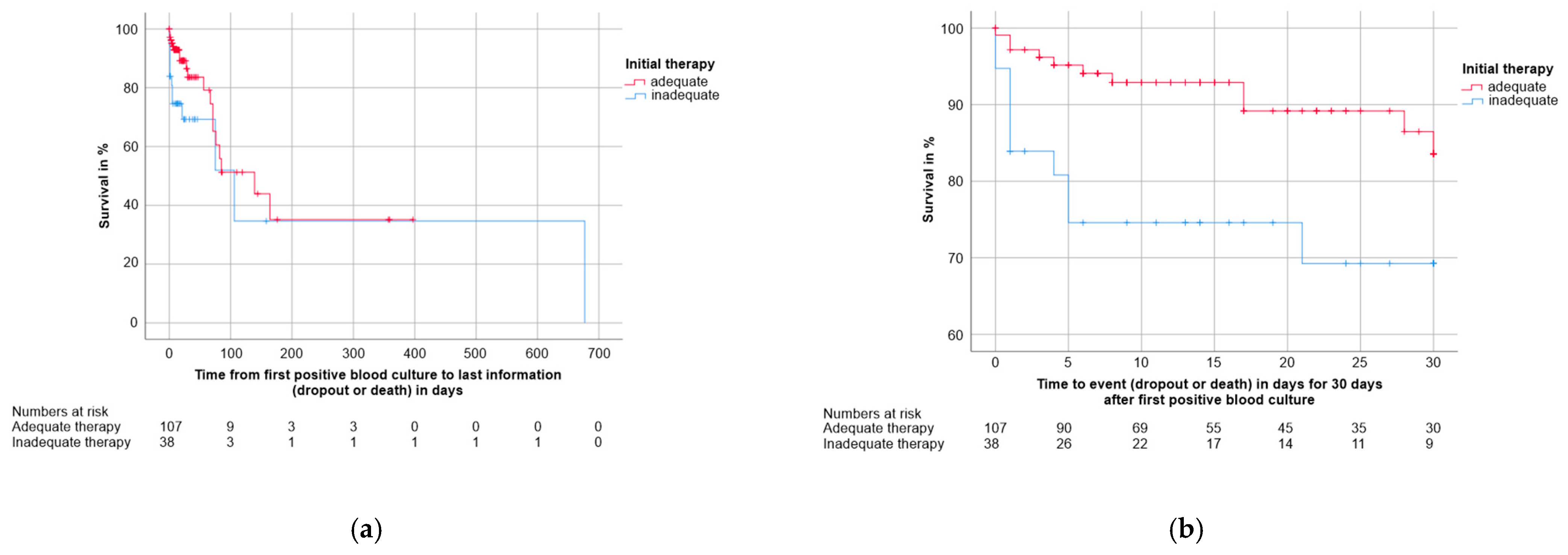
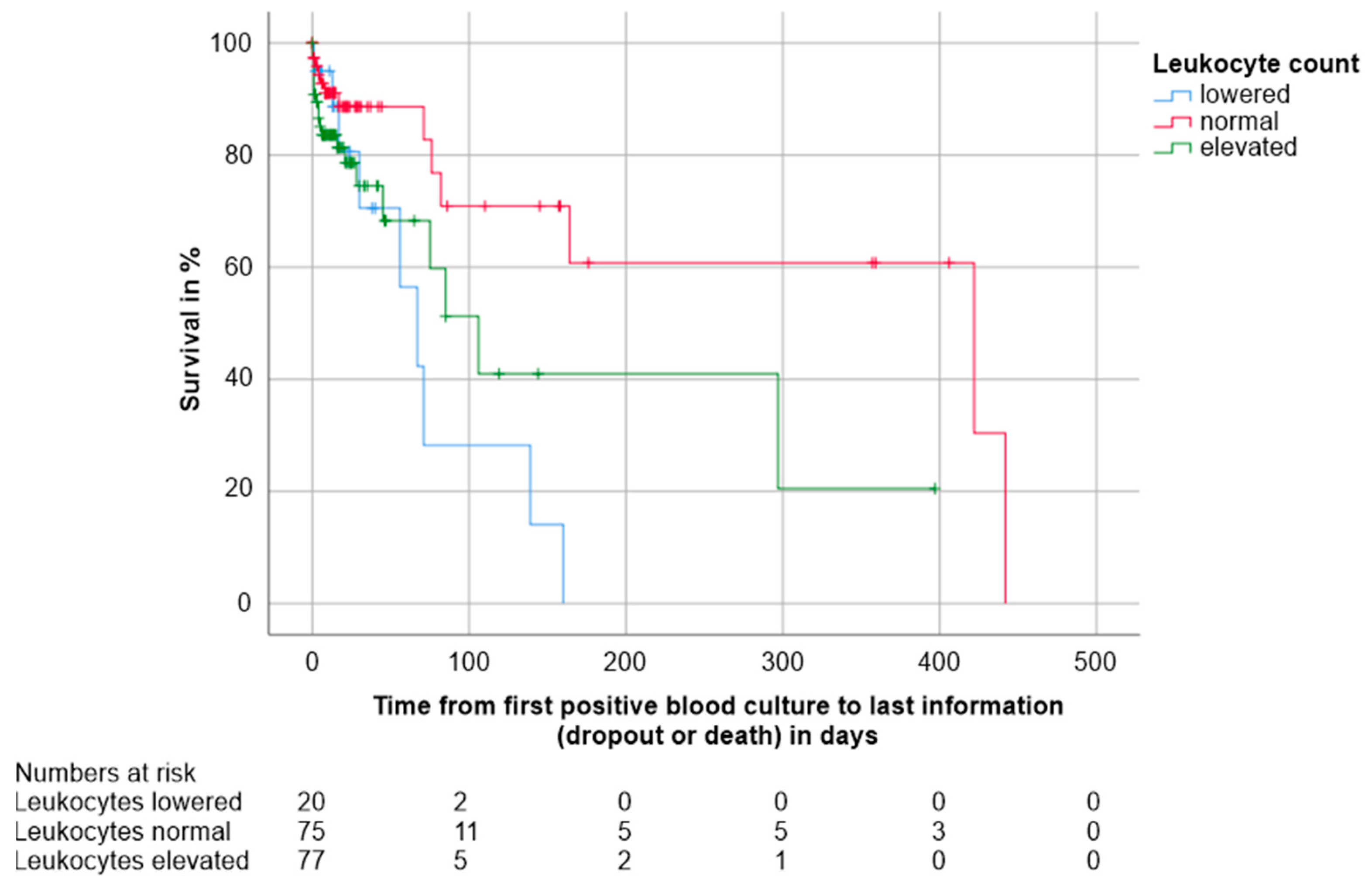
3.4. Factors with an Impact on Survival and Mortality
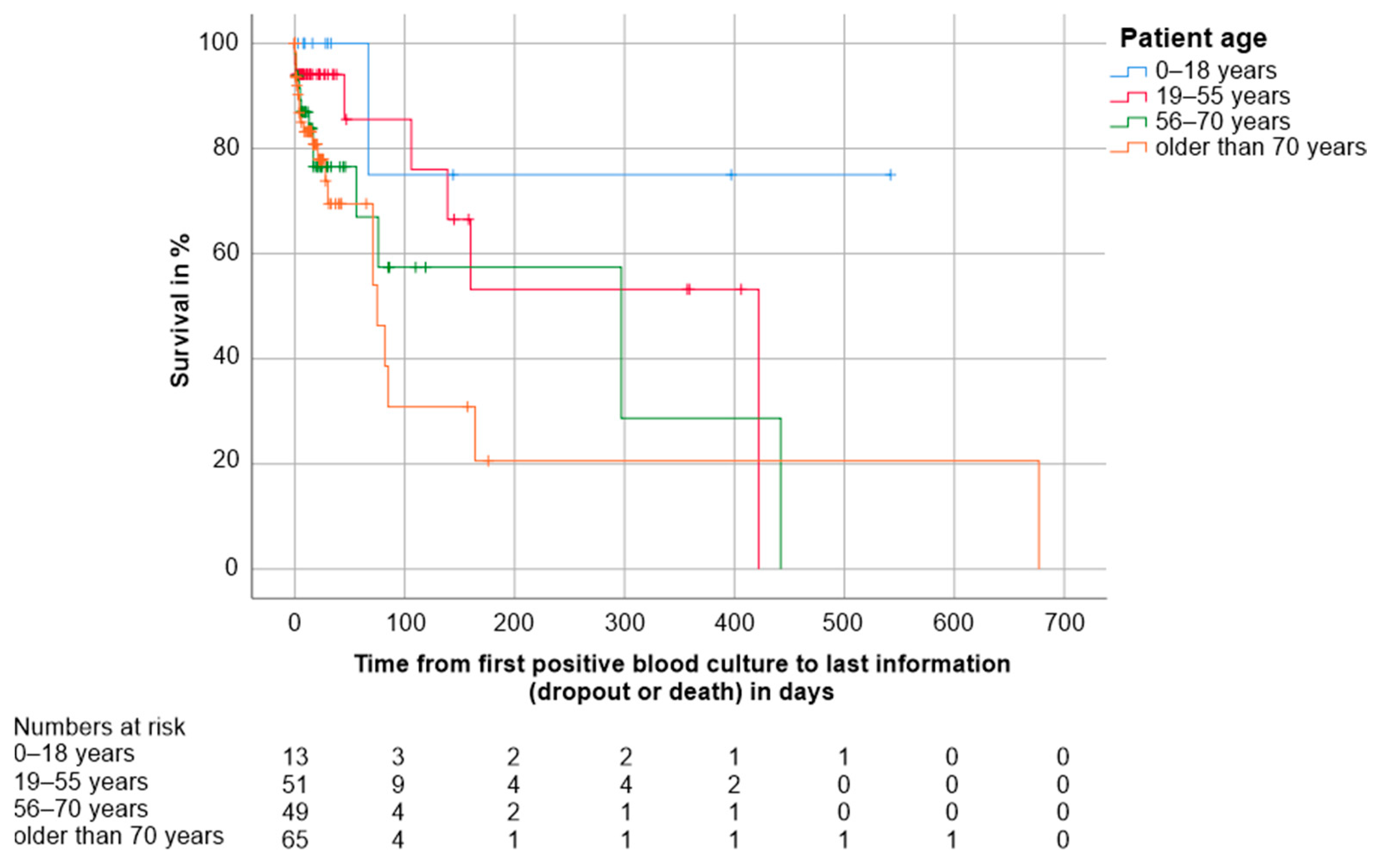
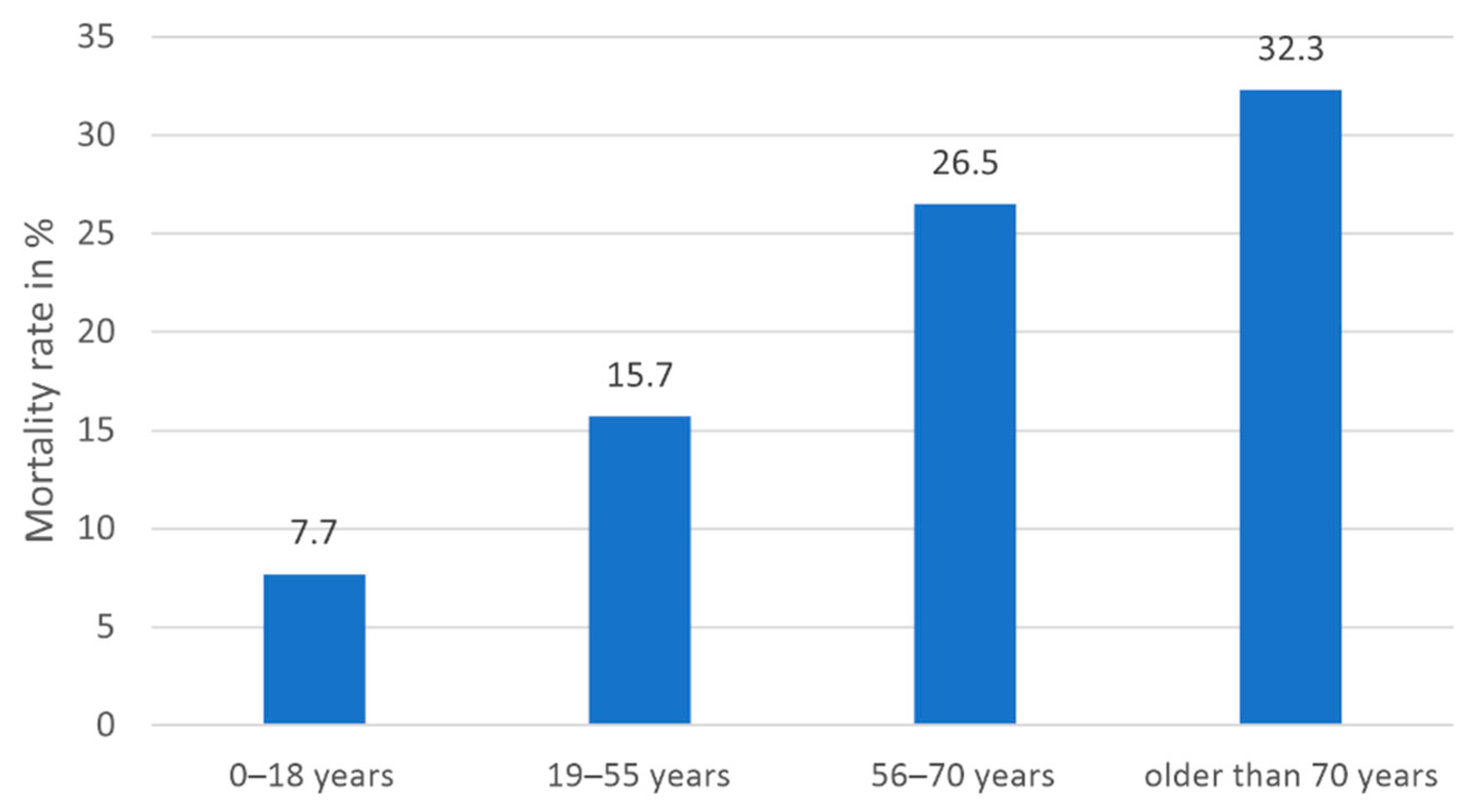
3.4.1. Host-Related Factors
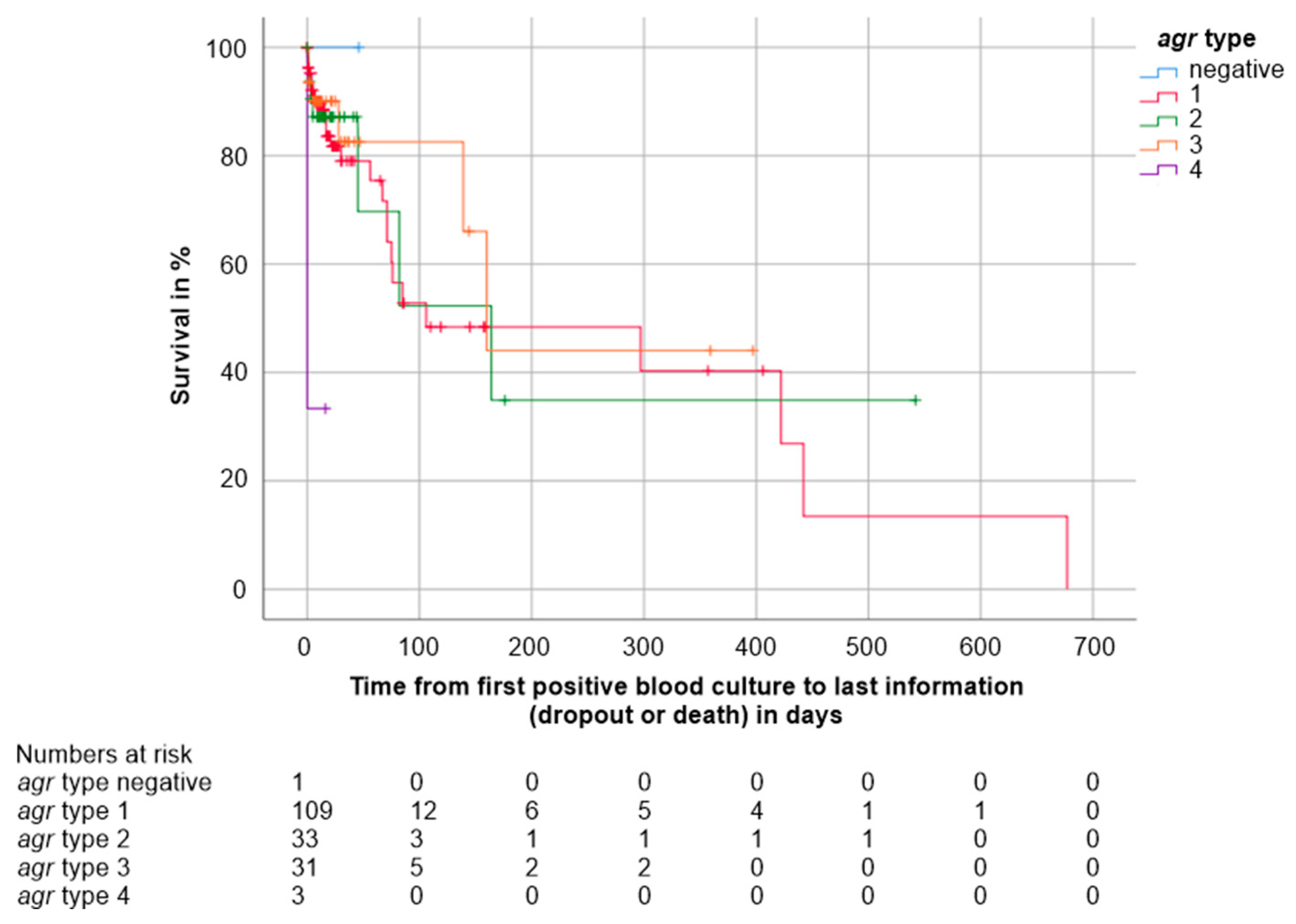
3.4.2. Pathogen-Related Factors
3.4.3. Clinical Factors
3.5. Factors with an Impact on Clinical Parameters and Treatment
3.5.1. Host-Related Factors
3.5.2. Pathogen-Related Factors
3.5.3. Host-Pathogen Interactions
3.6. Correlations between Pathogen Features
4. Discussion
4.1. Demographic and Clinical Data
4.2. Administered Antimicrobial Therapy
4.3. Colony Size and Hemolysis Activity
4.4. Biofilm
4.5. Nuclease
4.6. Clonality
4.7. Methicillin Resistance
4.8. agr Type and Toxin Gene Prevalence
4.9. Study Limitations and Strengths
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Used in | Reference |
|---|---|---|
| S. aureus RN4220 | Delta-toxin assay | [14] |
| S. aureus RN6607 | Delta-toxin assay | [14] |
| S. aureus Mu3 | Delta-toxin assay | [14] |
| S. epidermidis RP62A | Biofilm assay | [97] |
| S. carnosus TM300 | Biofilm assay | [97] |
| S. aureus AH1263 wt | Nuclease assay | [69] |
| S. aureus AH1680 (Δnuc1) | Nuclease assay | [69] |
| Gene | Primer | Sequence (5′–3′) | Reference |
|---|---|---|---|
| spa | forward 1095F | AGACGATCCTTCGGTGAGC | [19] |
| reverse 1517R | GCTTTTGCAATGTCATTTACTG | ||
| mecA | mec-5 forward | AAAATCGATGGTAAAGGTTGGC | [21] |
| mec-6 reverse | AGTTCTGCAGTACCGGATTTGC | ||
| agr | agrSA-KON1 forward | ATGCACATGGTGCACATGC | [22] |
| agrSA1-2 reverse | GTCACAAGTACTATAAGCTGCGAT | ||
| agrSA2-2 reverse | TATTACTAATTGAAAAGTGCCATAGC | ||
| agrSA3-2 reverse | GTAATGTAATAGCTTGTATAATAATACCCAG | ||
| agrSA4-2 reverse | CGATAATGCCGTAATACCCG | ||
| sea | sea-3 forward | CCTTTGGAAACGGTTAAAACG | [23] |
| sea-4 reverse | TCTGAACCTTCCCATCAAAAAC | ||
| seb | seb-1 forward | TCGCATCAAACTGACAAACG | [23] |
| seb-4 reverse | GCAGGTACTCTATAAGTGCCTGC | ||
| sec | sec-3 forward | CTCAAGAACTAGACATAAAAGCTAGG | [23] |
| sec-4 reverse | TCAAAATCGGATTAACATTATCC | ||
| sed | sed-3 forward | CTAGTTTGGTAATATCTCCTTTAAACG | [23] |
| sed-4 reverse | TTAATGCTATATCTTATAGGGTAAACATC | ||
| see | see-2 forward | TAACTTACCGTGGACCCTTC | [23] |
| see-3 reverse | CAGTACCTATAGATAAAGTTAAAACAAGC | ||
| seg | seg-1 forward | AATGCTCAACCCGATCCTA | [24] |
| seg-4 reverse | CTTCCTTCAACAGGTGGAGAC | ||
| seh | seh-1 forward | TTAGAAATCAAGGTGATAGTGGC | [24] |
| seh-2 reverse | TTTTGAATACCATCTACCCAAAC | ||
| sei | sei-1 forward | GCCACTTTATCAGGACAATACTT | [24] |
| sei-2 reverse | AAAACTTACAGGCAGTCCATCTC | ||
| sej | sej-1 forward | CTCCCTGACGTTAACACTACTAATAA | [24] |
| sej-2 reverse | TTGTCTGGATATTGACCTATAACATT | ||
| tst | tst-3 foward | AAGCCCTTTGTTGCTTGCG | [23] |
| tst-6 reverse | ATCGAACTTTGGCCCATACTTT | ||
| pvl | pvl-1 forward | ATCATTAGGTAAAATGTCTGGACATGATCCA | [25] |
| pvl-2 reverse | GCATCAASTGTATTGGATAGCAAAAGC | ||
| hlg | hlg-1 forward | GCCAATCCGTTATTAGAAAATGC | [25] |
| hlg-2 reverse | CCATAGAAGTAGCAACGGAT | ||
| eta | eta-3 forward | CTAGTGCATTTGTTATTCAAGACG | [23] |
| eta-4 reverse | TGCATTGACACCATAGTACTTATTC | ||
| etb | etb-3 forward | ACGGCTATATACATTCAATTCAATG | [23] |
| etb-4 reverse | AAAGTTATTCATTTAATGCACTGTCTC | ||
| etd | etd-1 forward | AACTATCATGTATCAAGG | [26] |
| etd-2 reverse | CAGAATTTCCCGACTCAG |
References
- European Centre for Disease Prevention and Control. Point Prevalence Survey of Healthcare-Associated Infections and Antimicrobial Use in European Acute Care Hospitals 2011–2012; European Centre for Disease Prevention and Control: Stockholm, Sweden, 2013. [Google Scholar] [CrossRef]
- Laupland, K.B. Incidence of bloodstream infection: A review of population-based studies. Clin. Microbiol. Infect. 2013, 19, 492–500. [Google Scholar] [CrossRef]
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed]
- Huggan, P.J.; Wells, J.E.; Browne, M.; Richardson, A.; Murdoch, D.R.; Chambers, S.T. Population-based epidemiology of Staphylococcus aureus bloodstream infection in Canterbury, New Zealand. Intern. Med. J. 2010, 40, 117–125. [Google Scholar] [CrossRef]
- Bodmann, K.-F.; Höhl, R.; Krüger, W.; Grabein, B.; Graninger, W. Calculated initial parenteral treatment of bacterial infections: Sepsis. Gms Infect. Dis. 2020, 8, Doc09. [Google Scholar] [CrossRef]
- Kampmeier, S.; Correa-Martinez, C.L.; Peters, G.; Mellmann, A.; Kahl, B.C. Personal microbiological consultations improve the therapeutic management of Staphylococcus aureus bacteremia. J. Infect. 2018, 77, 349–356. [Google Scholar] [CrossRef]
- van Hal, S.J.; Jensen, S.O.; Vaska, V.L.; Espedido, B.A.; Paterson, D.L.; Gosbell, I.B. Predictors of Mortality in Staphylococcus aureus Bacteremia. Clin. Microbiol. Rev. 2012, 25, 362–386. [Google Scholar] [CrossRef] [PubMed]
- Kaasch, A.J.; Barlow, G.; Edgeworth, J.D.; Fowler, V.G.; Hellmich, M.; Hopkins, S.; Kern, W.V.; Llewelyn, M.J.; Rieg, S.; Rodriguez-Baño, J.; et al. Staphylococcus aureus bloodstream infection: A pooled analysis of five prospective, observational studies. J. Infect. 2014, 68, 242–251. [Google Scholar] [CrossRef]
- Seifert, H.; Wisplinghoff, H.; Kaasch, A.; Achilles, K.; Langhorst, A.; Peyerl-Hoffmann, G.; Woehrmann, A.; Fätkenheuer, G.; Salzberger, B.; Kern, W.V. A prospective multicenter study of Staphylococcus aureus bacteremia—Results from the “Invasive Staphylococcus aureus Infection Cohort” (INSTINCT) study. Dtsch. Med. Wochenschr. 2008, 133, 340–345. [Google Scholar] [CrossRef]
- Kuehl, R.; Morata, L.; Boeing, C.; Subirana, I.; Seifert, H.; Rieg, S.; Kern, W.V.; Kim, H.B.; Kim, E.S.; Liao, C.H.; et al. Defining persistent Staphylococcus aureus bacteraemia: Secondary analysis of a prospective cohort study. Lancet Infect. Dis. 2020, 20, 1409–1417. [Google Scholar] [CrossRef]
- Le Moing, V.; Alla, F.; Doco-Lecompte, T.; Delahaye, F.; Piroth, L.; Chirouze, C.; Tattevin, P.; Lavigne, J.-P.; Erpelding, M.-L.; Hoen, B.; et al. Staphylococcus aureus Bloodstream Infection and Endocarditis—A Prospective Cohort Study. PLoS ONE 2015, 10, e0127385. [Google Scholar] [CrossRef] [PubMed]
- Bodmann, K.F.; Grabein, B.; Expert Commission of the Paul-Ehrlich Society for Chemotherapy. Recommendations for empiric parenteral initial antibiotic therapy of bacterial diseases in adults: Update 2010. GMS Infect. Dis. 2014. [Google Scholar] [CrossRef]
- Smyth, C.J.; Mollby, R.; Wadstrom, T. Phenomenon of hot cold hemolysis: Chelator induced lysis of sphingomyelinase treated erythrocytes. Infect. Immun. 1975, 12, 1104–1111. [Google Scholar] [CrossRef]
- Traber, K.E.; Lee, E.; Benon, S.; Corrigan, R.; Cantera, M.; Shopsin, B.; Novick, R.P. agr function in clinical Staphylococcus aureus isolates. Microbiology 2008, 154, 2265–2274. [Google Scholar] [CrossRef]
- Christensen, G.D.; Simpson, W.A.; Bisno, A.L.; Beachey, E.H. Adherence of slime-producing strains of Staphylococcus epidermidis to smooth surfaces. Infect. Immun. 1982, 37, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Stepanović, S.; Vuković, D.; Dakić, I.; Savić, B.; Švabić-Vlahović, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Schilcher, K.; Andreoni, F.; Uchiyama, S.; Ogawa, T.; Schuepbach, R.A.; Zinkernagel, A.S. Increased neutrophil extracellular trap-mediated Staphylococcus aureus clearance through inhibition of nuclease activity by clindamycin and immunoglobulin. J. Infect. Dis. 2014, 210, 473–782. [Google Scholar] [CrossRef] [PubMed]
- Herzog, S.; Dach, F.; de Buhr, N.; Niemann, S.; Schlagowski, J.; Chaves-Moreno, D.; Neumann, C.; Goretzko, J.; Schwierzeck, V.; Mellmann, A.; et al. High Nuclease Activity of Long Persisting Staphylococcus aureus Isolates Within the Airways of Cystic Fibrosis Patients Protects Against NET-Mediated Killing. Front. Immunol. 2019, 10, 2552. [Google Scholar] [CrossRef]
- Harmsen, D.; Claus, H.; Witte, W.; Rothgänger, J.; Claus, H.; Turnwald, D.; Vogel, U. Typing of Methicillin-Resistant Staphylococcus aureus in a University Hospital Setting by Using Novel Software for spa Repeat Determination and Database Management. J. Clin. Microbiol. 2003, 41, 5442–5448. [Google Scholar] [CrossRef] [PubMed]
- Mellmann, A.; Weniger, T.; Berssenbrügge, C.; Rothgänger, J.; Sammeth, M.; Stoye, J.; Harmsen, D. Based Upon Repeat Pattern (BURP): An algorithm to characterize the long-term evolution of Staphylococcus aureus populations based on spa polymorphisms. BMC Microbiol. 2007, 7, 98. [Google Scholar] [CrossRef]
- Murakami, K.; Minamide, W.; Wada, K.; Nakamura, E.; Teraoka, H.; Watanabe, S. Identification of methicillin-resistant strains of staphylococci by polymerase chain reaction. J. Clin. Microbiol. 1991, 29, 2240–2244. [Google Scholar] [CrossRef]
- Lina, G.; Boutite, F.; Tristan, A.; Bes, M.; Etienne, J.; Vandenesch, F. Bacterial Competition for Human Nasal Cavity Colonization: Role of Staphylococcal agr Alleles. Appl. Environ. Microbiol. 2003, 69, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Roth, R.; Peters, G. Rapid and specific detection of toxigenic Staphylococcus aureus: Use of two multiplex PCR enzyme immunoassays for amplification and hybridization of staphylococcal enterotoxin genes, exfoliative toxin genes, and toxic shock syndrome toxin 1 gene. J. Clin. Microbiol. 1998, 36, 2548–2553. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Friedrich, A.W.; Lubritz, G.; Weilert, M.; Peters, G.; Von Eiff, C. Prevalence of genes encoding pyrogenic toxin superantigens and exfoliative toxins among strains of Staphylococcus aureus isolated from blood and nasal specimens. J. Clin. Microbiol. 2003, 41, 1434–1439. [Google Scholar] [CrossRef]
- Von Eiff, C.; Friedrich, A.W.; Peters, G.; Becker, K. Prevalence of genes encoding for members of the staphylococcal leukotoxin family among clinical isolates of Staphylococcus aureus. Diagn. Microbiol. Infect. Dis. 2004, 49, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Nishifuji, K.; Sasaki, M.; Fudaba, Y.; Aepfelbacher, M.; Takata, T.; Ohara, M.; Komatsuzawa, H.; Amagai, M.; Sugai, M. Identification of the Staphylococcus aureus etd pathogenicity island which encodes a novel exfoliative toxin, ETD, and EDIN-B. Infect. Immun. 2002, 70, 5835–5845. [Google Scholar] [CrossRef]
- Yahav, D.; Schlesinger, A.; Shaked, H.; Goldberg, E.; Paul, M.; Bishara, J.; Leibovici, L. Clinical presentation, management and outcomes of Staph aureus bacteremia (SAB) in older adults. Aging Clin. Exp. Res. 2017, 29, 127–133. [Google Scholar] [CrossRef]
- Gasch, O.; Camoez, M.; Dominguez, M.A.; Padilla, B.; Pintado, V.; Almirante, B.; Molina, J.; Lopez-Medrano, F.; Ruiz, E.; Martinez, J.A.; et al. Predictive factors for mortality in patients with methicillin-resistant Staphylococcus aureus bloodstream infection: Impact on outcome of host, microorganism and therapy. Clin. Microbiol. Infect. 2013, 19, 1049–1057. [Google Scholar] [CrossRef]
- McClelland, R.S.; Fowler, V.G.; Sanders, L.L.; Gottlieb, G.; Kong, L.K.; Sexton, D.J.; Schmader, K.; Lanclos, K.D.; Corey, G.R. Staphylococcus aureus bacteremia among elderly vs younger adult patients: Comparison of clinical features and mortality. Arch. Intern. Med. 1999, 159, 1244–1247. [Google Scholar] [CrossRef]
- Cassone, M.; Mody, L. Colonization with Multidrug-Resistant Organisms in Nursing Homes: Scope, Importance, and Management. Curr. Geriatr. Rep. 2015, 4, 87–95. [Google Scholar] [CrossRef]
- Katz, M.J.; Roghmann, M.C. Healthcare-associated infections in the elderly: What’s new. Curr. Opin. Infect. Dis. 2016, 29, 388–393. [Google Scholar] [CrossRef]
- Mölkänen, T.; Rostila, A.; Ruotsalainen, E.; Alanne, M.; Perola, M.; Järvinen, A. Genetic polymorphism of the C-reactive protein (CRP) gene and a deep infection focus determine maximal serum CRP level in Staphylococcus aureus bacteremia. Eur. J. Clin. Microbiol. Infect. Dis. 2010, 29, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Guillamet, M.C.V.; Vazquez, R.; Deaton, B.; Shroba, J.; Vazquez, L.; Mercier, R.C. Host-Pathogen-Treatment Triad: Host Factors Matter Most in Methicillin-Resistant Staphylococcus aureus Bacteremia Outcomes. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [PubMed]
- Kaech, C.; Elzi, L.; Sendi, P.; Frei, R.; Laifer, G.; Bassetti, S.; Fluckiger, U. Course and outcome of Staphylococcus aureus bacteraemia: A retrospective analysis of 308 episodes in a Swiss tertiary-care centre. Clin. Microbiol. Infect. 2006, 12, 345–352. [Google Scholar] [CrossRef]
- Recker, M.; Laabei, M.; Toleman, M.S.; Reuter, S.; Saunderson, R.B.; Blane, B.; Török, M.E.; Ouadi, K.; Stevens, E.; Yokoyama, M.; et al. Clonal differences in Staphylococcus aureus bacteraemia-associated mortality. Nat. Microbiol. 2017, 2, 1381–1388. [Google Scholar] [CrossRef]
- Pinchuk, I.V.; Beswick, E.J.; Reyes, V.E. Staphylococcal Enterotoxins. Toxins 2010, 2, 2177–2197. [Google Scholar] [CrossRef] [PubMed]
- Jarraud, S.; Cozon, G.; Vandenesch, F.; Bes, M.; Etienne, J.; Lina, G. Involvement of Enterotoxins G and I in Staphylococcal Toxic Shock Syndrome and Staphylococcal Scarlet Fever. J. Clin. Microbiol. 1999, 37, 2446–2449. [Google Scholar] [CrossRef]
- Paul, M.; Kariv, G.; Goldberg, E.; Raskin, M.; Shaked, H.; Hazzan, R.; Samra, Z.; Paghis, D.; Bishara, J.; Leibovici, L. Importance of appropriate empirical antibiotic therapy for methicillin-resistant Staphylococcus aureus bacteraemia. J. Antimicrob. Chemother. 2010, 65, 2658–2665. [Google Scholar] [CrossRef] [PubMed]
- Rieg, S.; Peyerl-Hoffmann, G.; de With, K.; Theilacker, C.; Wagner, D.; Hübner, J.; Dettenkofer, M.; Kaasch, A.; Seifert, H.; Schneider, C.; et al. Mortality of S. aureus bacteremia and infectious diseases specialist consultation—A study of 521 patients in Germany. J. Infect. 2009, 59, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, T.C.; Price, C.S.; Sabel, A.L.; Mehler, P.S.; Burman, W.J. Impact of routine infectious diseases service consultation on the evaluation, management, and outcomes of Staphylococcus aureus bacteremia. Clin. Infect. Dis. 2008, 46, 1000–1008. [Google Scholar] [CrossRef]
- Proctor, R.A.; von Eiff, C.; Kahl, B.C.; Becker, K.; McNamara, P.; Herrmann, M.; Peters, G. Small colony variants: A pathogenic form of bacteria that facilitates persistent and recurrent infections. Nat. Rev. Microbiol. 2006, 4, 295–305. [Google Scholar] [CrossRef]
- Kahl, B.C.; Becker, K.; Löffler, B. Clinical significance and pathogenesis of staphylococcal small colony variants in persistent infections. Clin. Microbiol. Rev. 2016, 29, 401–427. [Google Scholar] [CrossRef] [PubMed]
- Bhakdi, S.; Tranum-Jensen, J. Alpha-toxin of Staphylococcus aureus. Microbiol. Rev. 1991, 55, 733–751. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Basis of Virulence in Community-Associated Methicillin-Resistant Staphylococcus aureus. Annu. Rev. Microbiol. 2010, 64, 143–162. [Google Scholar] [CrossRef]
- Wilke, G.A.; Wardenburg, J.B. Role of a disintegrin and metalloprotease 10 in Staphylococcus aureus α-hemolysin—Mediated cellular injury. Proc. Natl. Acad. Sci. USA 2010, 107, 13473–13478. [Google Scholar] [CrossRef] [PubMed]
- Sharma-Kuinkel, B.K.; Wu, Y.; Tabor, D.E.; Mok, H.; Sellman, B.R.; Jenkins, A.; Yu, L.; Jafri, H.S.; Rude, T.H.; Ruffin, F.; et al. Characterization of Alpha-Toxin hla Gene Variants, Alpha-Toxin Expression Levels, and Levels of Antibody to Alpha-Toxin in Hemodialysis and Postsurgical Patients with Staphylococcus aureus. J. Clin. Microbiol. 2015, 53, 227–236. [Google Scholar] [CrossRef]
- Powers, M.E.; Wardenburg, J.B. Igniting the Fire: Staphylococcus aureus Virulence Factors in the Pathogenesis of Sepsis. PLoS Pathog. 2014, 10, e1003871. [Google Scholar] [CrossRef]
- Oganesyan, V.; Peng, L.; Damschroder, M.M.; Cheng, L.; Sadowska, A.; Tkaczyk, C.; Sellman, B.R.; Wu, H.; Dall’Acqua, W.F. Mechanisms of neutralization of a human anti-α-toxin antibody. J. Biol. Chem. 2014, 289, 29874–29880. [Google Scholar] [CrossRef]
- Huseby, M.; Shi, K.; Kent Brown, C.; Digre, J.; Mengistu, F.; Keun, S.S.; Bohach, G.A.; Schlievert, P.M.; Ohlendorf, D.H.; Earhart, C.A. Structure and Biological Activities of Beta Toxin from Staphylococcus aureus. J. Bacteriol. 2007, 189, 8719–8726. [Google Scholar] [CrossRef]
- Vandenesch, F.; Lina, G.; Henry, T. Staphylococcus aureus hemolysins, bi-component leukocidins, and cytolytic peptides: A redundant arsenal of membrane-damaging virulence factors? Front. Cell. Infect. Microbiol. 2012, 2, 12. [Google Scholar] [CrossRef]
- Winkler, K.C.; de Waart, J.; Grootsen, C. Lysogenic conversion of staphylococci to loss of beta-toxin. J. Gen. Microbiol. 1965, 39, 321–333. [Google Scholar] [CrossRef]
- Coleman, D.C.; Arbuthnott, J.P.; Pomeroy, H.M.; Birkbeck, T.H. Cloning and expression in Escherichia coli and Staphylococcus aureus of the beta-lysin determinant from Staphylococcus aureus: Evidence that bacteriophage conversion of beta-lysin activity is caused by insertional inactivation of the beta-lysin determinant. Microb. Pathog. 1986, 1, 549–564. [Google Scholar] [CrossRef]
- Salgado-Pabón, W.; Herrera, A.; Vu, B.G.; Stach, C.S.; Merriman, J.A.; Spaulding, A.R.; Schlievert, P.M. Staphylococcus aureus β-toxin production is common in strains with the β-toxin gene inactivated by bacteriophage. J. Infect. Dis. 2014, 210, 784–792. [Google Scholar] [CrossRef] [PubMed]
- De Haas, C.J.C.; Veldkamp, K.E.; Peschel, A.; Weerkamp, F.; Van Wamel, W.J.B.; Heezius, E.C.J.M.; Poppelier, M.J.J.G.; Van Kessel, K.P.M.; Van Strijp, J.A.G. Chemotaxis Inhibitory Protein of Staphylococcus aureus, a Bacterial Antiinflammatory Agent. J. Exp. Med. 2004, 199, 687–695. [Google Scholar] [CrossRef]
- Aarestrup, F.M.; Larsen, H.D.; Eriksen, N.H.R.; Elsberg, C.S.; Jensen, N.E. Frequency of α- and β-haemolysin in Staphylococcus aureus of bovine and human origin. APMIS 1999, 107, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Monecke, S.; Kuhnert, P.; Hotzel, H.; Slickers, P.; Ehricht, R. Microarray based study on virulence-associated genes and resistance determinants of Staphylococcus aureus isolates from cattle. Vet. Microbiol. 2007, 125, 128–140. [Google Scholar] [CrossRef]
- Van Alen, S.; Ballhausen, B.; Kaspar, U.; Köck, R.; Becker, K. Prevalence and genomic structure of bacteriophage phi3 in human-derived livestock-associated methicillin-resistant Staphylococcus aureus isolates from 2000 to 2015. J. Clin. Microbiol. 2018, 56, e00140-18. [Google Scholar] [CrossRef]
- Recsei, P.; Kreiswirth, B.; O’Reilly, M.; Schlievert, P.; Gruss, A.; Novick, R.P. Regulation of exoprotein gene expression in Staphylococcus aureus by agr. MGG Mol. Gen. Genet. 1986, 202, 58–61. [Google Scholar] [CrossRef] [PubMed]
- Verdon, J.; Girardin, N.; Lacombe, C.; Berjeaud, J.M.; Héchard, Y. δ-hemolysin, an update on a membrane-interacting peptide. Peptides 2009, 30, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Gagnaire, J.; Dauwalder, O.; Boisset, S.; Khau, D.; Freydière, A.M.; Ader, F.; Bes, M.; Lina, G.; Tristan, A.; Reverdy, M.E.; et al. Detection of Staphylococcus aureus Delta-Toxin Production by Whole-Cell MALDI-TOF Mass Spectrometry. PLoS ONE 2012, 7, e40660. [Google Scholar] [CrossRef]
- Sakoulas, G.; Eliopoulos, G.M.; Moellering, R.C.; Wennersten, C.; Venkataraman, L.; Novick, R.P.; Gold, H.S. Accessory Gene Regulator (agr) Locus in Geographically Diverse Staphylococcus aureus Isolates with Reduced Susceptibility to Vancomycin. Antimicrob. Agents Chemother. 2002, 46, 1492–1502. [Google Scholar] [CrossRef] [PubMed]
- Kwiecinski, J.M.; Jacobsson, G.; Horswill, A.R.; Josefsson, E.; Jin, T. Biofilm formation by Staphylococcus aureus clinical isolates correlates with the infection type. Infect. Dis. (Auckl.) 2019, 51, 446–451. [Google Scholar] [CrossRef]
- Guembe, M.; Alonso, B.; Lucio, J.; Pérez-Granda, M.J.; Cruces, R.; Sánchez-Carrillo, C.; Fernández-Cruz, A.; Bouza, E. Biofilm production is not associated with poor clinical outcome in 485 patients with Staphylococcus aureus bacteraemia. Clin. Microbiol. Infect. 2018, 24, 659.e1–659.e3. [Google Scholar] [CrossRef]
- Naicker, P.R.; Karayem, K.; Hoek, K.G.P.; Harvey, J.; Wasserman, E. Biofilm formation in invasive Staphylococcus aureus isolates is associated with the clonal lineage. Microb. Pathog. 2016, 90, 41–49. [Google Scholar] [CrossRef]
- Seidl, K.; Bayer, A.S.; Fowler, V.G.; McKinnell, J.A.; Abdel Hady, W.; Sakoulas, G.; Yeaman, M.R.; Xiong, Y.Q.; Xiong, Y.Q. Combinatorial Phenotypic Signatures Distinguish Persistent from Resolving Methicillin-Resistant Staphylococcus aureus Bacteremia Isolates. Antimicrob. Agents Chemother. 2011, 55, 575–582. [Google Scholar] [CrossRef]
- Trouillet-Assant, S.; Lelièvre, L.; Martins-Simões, P.; Gonzaga, L.; Tasse, J.; Valour, F.; Rasigade, J.-P.; Vandenesch, F.; Muniz Guedes, R.L.; Ribeiro de Vasconcelos, A.T.; et al. Adaptive processes of Staphylococcus aureus isolates during the progression from acute to chronic bone and joint infections in patients. Cell. Microbiol. 2016, 18, 1405–1414. [Google Scholar] [CrossRef]
- Lister, J.L.; Horswill, A.R. Staphylococcus aureus biofilms: Recent developments in biofilm dispersal. Front. Cell. Infect. Microbiol. 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Vuong, C.; Saenz, H.L.; Götz, F.; Otto, M. Impact of the agr Quorum-Sensing System on Adherence to Polystyrene in Staphylococcus aureus. J. Infect. Dis. 2000, 182, 1688–1693. [Google Scholar] [CrossRef]
- Kiedrowski, M.R.; Kavanaugh, J.S.; Malone, C.L.; Mootz, J.M.; Voyich, J.M.; Smeltzer, M.S.; Bayles, K.W.; Horswill, A.R. Nuclease Modulates Biofilm Formation in Community-Associated Methicillin-Resistant Staphylococcus aureus. PLoS ONE 2011, 6, e26714. [Google Scholar] [CrossRef] [PubMed]
- Tsang, L.H.; Cassat, J.E.; Shaw, L.N.; Beenken, K.E.; Smeltzer, M.S. Factors Contributing to the Biofilm-Deficient Phenotype of Staphylococcus aureus sarA Mutants. PLoS ONE 2008, 3, e03361. [Google Scholar] [CrossRef]
- Izano, E.A.; Amarante, M.A.; Kher, W.B.; Kaplan, J.B. Differential Roles of Poly-N-Acetylglucosamine Surface Polysaccharide and Extracellular DNA in Staphylococcus aureus and Staphylococcus epidermidis Biofilms. Appl. Environ. Microbiol. 2008, 74, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Mann, E.E.; Rice, K.C.; Boles, B.R.; Endres, J.L.; Ranjit, D.; Chandramohan, L.; Tsang, L.H.; Smeltzer, M.S.; Horswill, A.R.; Bayles, K.W. Modulation of eDNA Release and Degradation Affects Staphylococcus aureus Biofilm Maturation. PLoS ONE 2009, 4, e05822. [Google Scholar] [CrossRef] [PubMed]
- Kinney, K.J.; Tran, P.M.; Gibson-Corley, K.N.; Forsythe, A.N.; Kulhankova, K.; Salgado-Pabón, W. Staphylococcal Enterotoxin C promotes Staphylococcus aureus Infective Endocarditis Independent of Superantigen Activity. bioRxiv 2019. [Google Scholar] [CrossRef]
- Bui, L.M.G.; Turnidge, J.D.; Kidd, S.P. The induction of Staphylococcus aureus biofilm formation or Small Colony Variants is a strain-specific response to host-generated chemical stresses. Microbes Infect. 2015, 17, 77–82. [Google Scholar] [CrossRef]
- Berends, E.T.M.; Horswill, A.R.; Haste, N.M.; Monestier, M.; Nizet, V.; Köckritz-Blickwede, M. von Nuclease Expression by Staphylococcus aureus Facilitates Escape from Neutrophil Extracellular Traps. J. Innate Immun. 2010, 2, 576. [Google Scholar] [CrossRef] [PubMed]
- Olson, M.E.; Nygaard, T.K.; Ackermann, L.; Watkins, R.L.; Zurek, O.W.; Pallister, K.B.; Griffith, S.; Kiedrowski, M.R.; Flack, C.E.; Kavanaugh, J.S.; et al. Staphylococcus aureus nuclease is an SaeRS-dependent virulence factor. Infect. Immun. 2013, 81, 1316–1324. [Google Scholar] [CrossRef]
- Humphreys, H.; Fitzpatick, F.; Harvey, B.J. Gender Differences in Rates of Carriage and Bloodstream Infection Caused by Methicillin-Resistant Staphylococcus aureus: Are They Real, Do They Matter and Why? Clin. Infect. Dis. 2015, 61, civ576. [Google Scholar] [CrossRef]
- Laupland, K.B.; Lyytikäinen, O.; Søgaard, M.; Kennedy, K.J.; Knudsen, J.D.; Ostergaard, C.; Galbraith, J.C.; Valiquette, L.; Jacobsson, G.; Collignon, P.; et al. The changing epidemiology of Staphylococcus aureus bloodstream infection: A multinational population-based surveillance study. Clin. Microbiol. Infect. 2013, 19, 465–471. [Google Scholar] [CrossRef]
- Lamagni, T.L.; Potz, N.; Powell, D.; Pebody, R.; Wilson, J.; Duckworth, G. Mortality in patients with meticillin-resistant Staphylococcus aureus bacteraemia, England 2004–2005. J. Hosp. Infect. 2011, 77, 16–20. [Google Scholar] [CrossRef]
- David, M.Z.; Daum, R.S.; Bayer, A.S.; Chambers, H.F.; Fowler, V.G.; Miller, L.G.; Ostrowsky, B.; Baesa, A.; Boyle-Vavra, S.; Eells, S.J.; et al. Staphylococcus aureus Bacteremia at 5 US Academic Medical Centers, 2008-2011: Significant Geographic Variation in Community-Onset Infections. Clin. Infect. Dis. 2014, 59, 798–807. [Google Scholar] [CrossRef]
- Becker, K.; Schaumburg, F.; Fegeler, C.; Friedrich, A.W.; Köck, R. Staphylococcus aureus from the German general population is highly diverse. Int. J. Med. Microbiol. 2017, 307, 21–27. [Google Scholar] [CrossRef]
- Souli, M.; Ruffin, F.; Choi, S.H.; Park, L.P.; Gao, S.; Lent, N.C.; Sharma-Kuinkel, B.K.; Thaden, J.T.; Maskarinec, S.A.; Wanda, L.; et al. Changing Characteristics of Staphylococcus aureus Bacteremia: Results from a 21-Year, Prospective, Longitudinal Study. Clin. Infect. Dis. 2019, 69, 1868–1877. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. Surveillance of Antimicrobial Resistance in Europe 2018; European Centre for Disease Prevention and Control: Stockholm, Sweden, 2019. [Google Scholar] [CrossRef]
- Cuny, C.; Wieler, L.H.; Witte, W. Livestock-Associated MRSA: The impact on humans. Antibiotics 2015, 4, 521–543. [Google Scholar] [CrossRef]
- Goerge, T.; Lorenz, M.B.; van Alen, S.; Hübner, N.O.; Becker, K.; Köck, R. MRSA colonization and infection among persons with occupational livestock exposure in Europe: Prevalence, preventive options and evidence. Vet. Microbiol. 2017, 200, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Köck, R.; Schaumburg, F.; Mellmann, A.; Köksal, M.; Jurke, A.; Becker, K.; Friedrich, A.W. Livestock-Associated Methicillin-Resistant Staphylococcus aureus (MRSA) as Causes of Human Infection and Colonization in Germany. PLoS ONE 2013, 8, e55040. [Google Scholar] [CrossRef]
- Kadlec, K.; Entorf, M.; Peters, T. Occurrence and characteristics of livestock-associated methicillin-resistant Staphylococcus aureus in quarter milk samples from dairy cows in Germany. Front. Microbiol. 2019, 10, 1295. [Google Scholar] [CrossRef] [PubMed]
- Cuny, C.; Layer, F.; Werner, G.; Harmsen, D.; Daniels-Haardt, I.; Jurke, A.; Mellmann, A.; Witte, W.; Köck, R. State-wide surveillance of antibiotic resistance patterns and spa types of methicillin-resistant Staphylococcus aureus from blood cultures in North Rhine-Westphalia, 2011–2013. Clin. Microbiol. Infect. 2015, 21, 750–757. [Google Scholar] [CrossRef]
- Cuny, C.; Köck, R.; Witte, W. Livestock associated MRSA (LA-MRSA) and its relevance for humans in Germany. Int. J. Med. Microbiol. 2013, 303, 331–337. [Google Scholar] [CrossRef]
- van Alen, S.; Ballhausen, B.; Peters, G.; Friedrich, A.W.; Mellmann, A.; Köck, R.; Becker, K. In the centre of an epidemic: Fifteen years of LA-MRSA CC398 at the University Hospital Münster. Vet. Microbiol. 2017, 200, 19–24. [Google Scholar] [CrossRef]
- Ji, G.; Beavis, R.; Novick, R.P. Bacterial interference caused by autoinducing peptide variants. Science 1997, 276, 2027–2030. [Google Scholar] [CrossRef]
- Jarraud, S.; Lyon, G.J.; Figueiredo, A.M.S.; Gerard, L.; Vandenesch, F.; Etienne, J.; Muir, T.W.; Novick, R.P. Exfoliatin-Producing Strains Define a Fourth agr Specificity Group in Staphylococcus aureus. J. Bacteriol. 2000, 182, 6517–6522. [Google Scholar] [CrossRef]
- Jarraud, S.; Mougel, C.; Thioulouse, J.; Lina, G.; Meugnier, H.; Forey, F.; Nesme, X.; Etienne, J.; Vandenesch, F. Relationships between Staphylococcus aureus Genetic Background, Virulence Factors, agr Groups (Alleles), and Human Disease. Infect. Immun. 2002, 70, 631–641. [Google Scholar] [CrossRef] [PubMed]
- Lange, J.; Heidenreich, K.; Higelin, K.; Dyck, K.; Marx, V.; Reichel, C.; Van Wamel, W.; Den Reijer, M.; Görlich, D.; Kahl, B.C. Staphylococcus aureus Pathogenicity in Cystic Fibrosis Patients-Results from an Observational Prospective Multicenter Study Concerning Virulence Genes, Phylogeny, and Gene Plasticity. Toxins 2020, 12, 279. [Google Scholar] [CrossRef] [PubMed]
- Grumann, D.; Nübel, U.; Bröker, B.M. Staphylococcus aureus toxins—Their functions and genetics. Infect. Genet. Evol. 2014, 21, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Nhan, T.X.; Leclercq, R.; Cattoir, V. Prevalence of toxin genes in consecutive clinical isolates of Staphylococcus aureus and clinical impact. Eur. J. Clin. Microbiol. Infect. Dis. 2011, 30, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Christensen, G.D.; Bisno, A.L.; Parisi, J.T.; McLaughlin, B.; Hester, M.G.; Luther, R.W. Nosocomial septicemia due to multiply antibiotic-resistant Staphylococcus epidermidis. Ann. Intern. Med. 1982, 96, 1–10. [Google Scholar] [CrossRef]








| First-Line Therapy | Second-Line Therapy |
|---|---|
| linezolid daptomycin glycopeptides (e.g., vancomycin, teicoplanin) |
| Parameter | Data |
|---|---|
| Sex - Female/male, n (%) | 74/104 (41.6/58.4) |
| Age in years - Median (IQR 1) - Mean (SD 2) - Range | 63.0 (49.8–74.0) 58.7 (21.0) 0–94 |
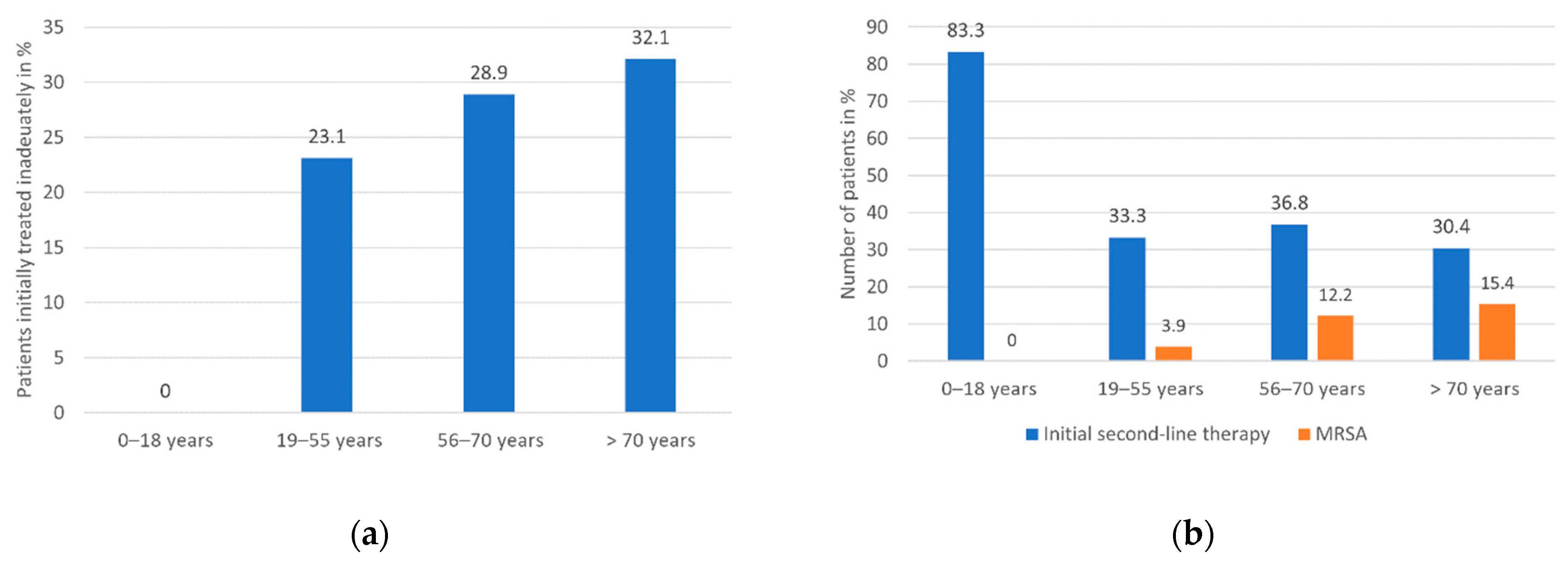
| Age in categories, n (%) - 0–18 years - 19–55 years - 56–70 years - >70 years | 13 (7.3) 51 (28.7) 49 (27.5) 65 (36.5) |
| Infection focus, n (%) - Intravascular device - Abscess and soft tissue - Pneumonia - Endocarditis - Bone or joint - Bone or joint implant - Others - Unclear - Missing | 71 (40.1) 23 (13.0) 22 (12.4) 16 (9.0) 11 (6.2) 10 (5.6) 5 (2.8) 19 (10.7) 1 (0.6) |
| Presence of fever ≥38 °C - Positive/negative, n (%) - Missing, n | 112/44 (71.8/28.2) 22 |
| CRP - Median (IQR) in mg/dL - Missing, n | 13.6 (7.3–23.5) 12 |
| CRP categories - Normal (≤0.5 mg/dL), n (%) - Elevated (>0.5 mg/dL), n (%) - Missing, n | 7 (4.2) 159 (95.8) 12 |
| Leukocyte count - Median (IQR) in × 10³/µL - Missing, n | 10.2 (6.5–15.5) 6 |
| Leukocyte categories - Lowered (<3.9 × 10³/µL), n (%) - Normal (3.9–10.9 × 10³/µL), n (%) - Elevated (>10.9 × 10³/µL), n (%) - Missing, n | 20 (11.6) 75 (43.6) 77 (44.8) 6 |
| Mortality, n (%) - Patient deaths overall - Patient deaths within first 30 days | 43 (24.2) 26 (14.6) |
| - Median follow-up time in days (CI 3) | 21.0 (16.5–25.5) |
| - Median overall survival in days (CI) | 160.0 (75.2–244.8) |
| Initial antimicrobial therapy - Adequate/inadequate, n (%) - MSSA: adequate/inadequate, n (%) - MRSA: adequate/inadequate, n (%) - Second-line antibiotics, n (%) - of these: applied to MRSA/MSSA, n (%) - of these: information missing, n - Missing, n | 107/38 (73.8/26.2) 96/36 (72.7/27.3) 10/2 (83.3/16.7) 54 (37.2) 10/43 (18.9/81.1) 1 33 |
| Parameter | Data |
|---|---|
| Colony size, n (%) - Normal phenotype (NP) - Small colony variant (SCV) phenotype - Mixed (NP and SCV) | 174 (97.8) 1 (0.6) 3 (1.7) |
| Alpha-toxin, n (%) - Positive/negative | 175/3 (98.3/1.7) |
| Beta-toxin, n (%) - Positive - Negative - Mixed | 19 (10.7) 156 (87.6) 3 (1.7) |
| Delta-toxin, n (%) - Positive/negative | 87/91 (48.9/51.1) |
| Biofilm in % of the positive control - Median (IQR) - Range | 0.2 (−0.5–1.5) −1.9–53.8 |
| Biofilm categories, n (%) - No biofilm formation - Low biofilm formation - Moderate biofilm formation - Strong biofilm formation | 157 (88.2) 18 (10.1) 2 (1.1) 1 (0.6) |
| Nuclease activity in RFU - Median (IQR) - Range | 528.6 (358.8–743.9) 97.8–2281.9 |
| Methicillin resistance - MSSA/MRSA, n (%) - Missing, n | 159/18 (89.8/10.2) 1 |
| agr type - agr 1, n (%) - agr 2, n (%) - agr 3, n (%) - agr 4, n (%) - Negative for all agr types, n (%) - Missing, n | 109 (61.6) 33 (18.6) 31 (17.5) 3 (1.7) 1 (0.6) 1 |
| Prevalence of toxin genes - sea, n (%) - seb, n (%) - sec, n (%) - sed, n (%) - see, n (%) - seg, n (%) - seh, n (%) - sei, n (%) - sej, n (%) - tst-1, n (%) - pvl, n (%) - hlg, n (%) - eta, n (%) - etb, n (%) - etd, n (%) - Missing, n | 13 (7.3) 6 (3.4) 37 (20.9) 10 (5.6) 2 (1.1) 103 (58.2) 9 (5.1) 104 (58.8) 10 (5.6) 23 (13.0) 2 (1.1) 176 (99.4) 2 (1.1) 0 (0) 3 (1.7) 1 |
| spa-CC | Number of Isolates | spa Types | Methicillin Resistance | agr Type |
|---|---|---|---|---|
| 084 | 38 | t084 (10), t091 (19), t144, t360, t491, t2616, t4802, t7071, t11193, t12178, t18220 | MSSA | agr 1 (20) agr 2 (16) agr 3 (1) |
| 012 | 21 | t012 (9), t018 (2), t021, t046, t090, t122 (2), t363, t726, t840, t964, t1654 | MSSA | agr 3 |
| 015 | 19 | t015, t050 (2), t069, t095, t102, t133, t230, t302, t550, t583, t728 (2), t1231, t1510, t2195, t4153, t5032, t18219 | MSSA | agr 1 |
| 005/032 | 18 | t005 (3), t032 (6), t223, t310, t608, t612, t1770, t2816, t5857, t12754, t18079 | 9/18 MRSA | agr 1 |
| 068/008 | 11 | t008 (5), t068, t068, t211, t2455, t7222, t18713 | 1/11 MRSA | agr 1 |
| 034/011 | 5 | t011 (2), t034, t571, t2576 | 4/5 MRSA | agr 1 |
| 864 | 4 | t153, t166, t352, t864 | MSSA | agr 3 (3) negative (1) |
| 065 | 3 | t040, t065, t6137 | MSSA | agr 1 |
| NF1 #1 | 4 | t002 (3), t311 | MSSA | agr 2 |
| NF #2 | 4 | t156 (2), t160 (2) | MSSA | agr 2 |
| NF #3 | 3 | t003 (2), t264 | MRSA | agr 2 |
| NF #4 | 3 | t267 (2), t692 | MSSA | agr 1 (2) agr 3 (1) |
| NF #5 | 2 | t127, t177 | MSSA | agr 3 |
| NF #6 | 2 | t364, t493 | MSSA | agr 1 |
| singletons | 29 | t056 (2), t078, t092, t100, t131, t148 (2), t159, t189, t216, t280, t335, t351, t428, t845, t933, t1305, t1430, t2227, t5488, t8108 (2), t17517, t18076, t18218, t18622, t18636, t18712 | 1/29 MRSA | mixed |
| excluded | 10 | t026 (4), t643, t693, t748, t1050, t1991, t3625 | MSSA | mixed |
| 0–18 Years | 19–55 Years | 56–70 Years | >70 Years | |
|---|---|---|---|---|
| Overall mortality in % | 7.7 | 15.7 | 26.5 | 32.3 |
| Median overall survival time in days (CI) | n.a. 1 | 422.0 (n.a.1) | 297.0 (0.0–618.4) | 75.0 (35.2–114.8) |
| Adequate Initial Therapy | Inadequate Initial Therapy | Log-Rank p | |
|---|---|---|---|
| Overall mortality in % | 18.7 | 34.2 | 0.041 |
| Median overall survival time in days (CI) | 139.0 (36.6–241.4) | 106.0 (20.2–191.8) | |
| 30-day mortality in % | 10.3 | 26.3 | 0.010 |
| Leukocytes Lowered | Leukocytes Normal | Leukocytes Elevated | |
|---|---|---|---|
| Overall mortality in % | 45.0 | 17.3 | 26.0 |
| Median overall survival time in days (CI) | 67.0 (40.5–93.5) | 422.0 (49.5–794.5) | 106.0 (62.7–149.3) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wächter, H.; Yörük, E.; Becker, K.; Görlich, D.; Kahl, B.C. Correlations of Host and Bacterial Characteristics with Clinical Parameters and Survival in Staphylococcus aureus Bacteremia. J. Clin. Med. 2021, 10, 1371. https://doi.org/10.3390/jcm10071371
Wächter H, Yörük E, Becker K, Görlich D, Kahl BC. Correlations of Host and Bacterial Characteristics with Clinical Parameters and Survival in Staphylococcus aureus Bacteremia. Journal of Clinical Medicine. 2021; 10(7):1371. https://doi.org/10.3390/jcm10071371
Chicago/Turabian StyleWächter, Hannah, Erdal Yörük, Karsten Becker, Dennis Görlich, and Barbara C. Kahl. 2021. "Correlations of Host and Bacterial Characteristics with Clinical Parameters and Survival in Staphylococcus aureus Bacteremia" Journal of Clinical Medicine 10, no. 7: 1371. https://doi.org/10.3390/jcm10071371
APA StyleWächter, H., Yörük, E., Becker, K., Görlich, D., & Kahl, B. C. (2021). Correlations of Host and Bacterial Characteristics with Clinical Parameters and Survival in Staphylococcus aureus Bacteremia. Journal of Clinical Medicine, 10(7), 1371. https://doi.org/10.3390/jcm10071371