
The Role of Extracellular Matrix in Skin Wound Healing

 , , ,
, , , 
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
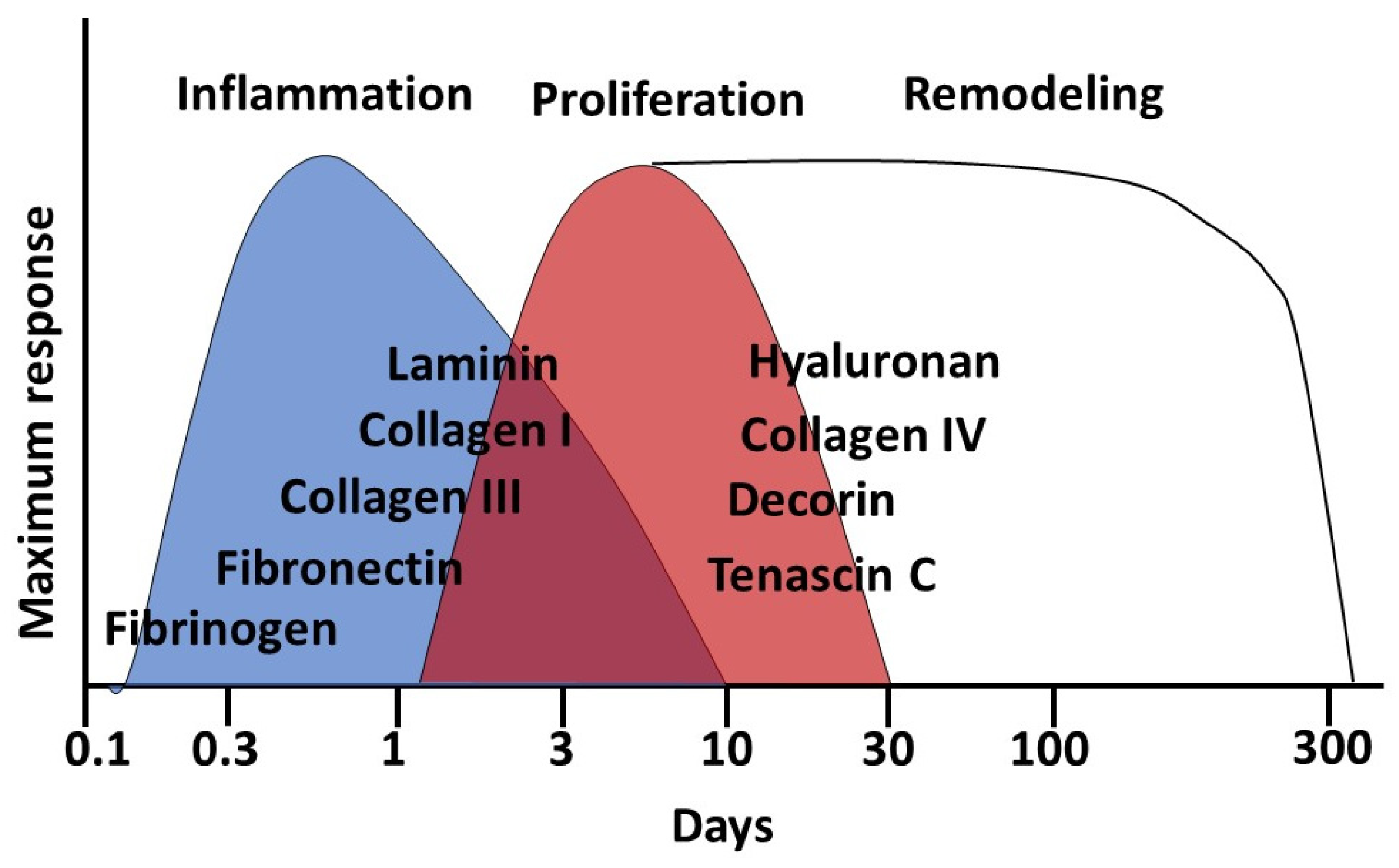
3.1. Stages of Wound Healing
- inflammation (with hemostasis);
- proliferation;
- remodeling.
3.2. Extracellular Matrix
3.3. The Composition of the Extracellular Matrix
3.4. The Role of Extracellular Matrix in Skin Wound Healing
3.5. Genetic Aspects of the Role of Extracellular Matrix in Wound Healing
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mostaço-Guidolin, L.; Rosin, N.L.; Hackett, T.L. Imaging Collagen in Scar Tissue: Developments in Second Harmonic Generation Microscopy for Biomedical Applications. Int. J. Mol. Sci. 2017, 18, 1772. [Google Scholar] [CrossRef] [PubMed]
- Ud-Din, S.; Volk, S.W.; Bayat, A. Regenerative healing, scar-free healing and scar formation across the species: Current concepts and future perspectives. Exp. Dermatol. 2014, 23, 615–619. [Google Scholar] [CrossRef]
- van den Broek, L.J.; Limandjaja, G.C.; Niessen, F.B.; Gibbs, S. Human hypertrophic and keloid scar models: Principles, limitations and future challenges from a tissue engineering perspective. Exp. Dermatol. 2014, 23, 382–386. [Google Scholar] [CrossRef] [Green Version]
- Kaplani, K.; Koutsi, S.; Armenis, V.; Skondra, F.G.; Karantzelis, N.; Champeris Tsaniras, S.; Taraviras, S. Wound healing related agents: Ongoing research and perspectives. Adv. Drug Deliv. Rev. 2018, 129, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Frykberg, R.G.; Banks, J. Challenges in the Treatment of Chronic Wounds. Adv. Wound Care 2015, 4, 560–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shih, B.; Bayat, A. Genetics of keloid scarring. Arch. Dermatol. Res. 2010, 302, 319–339. [Google Scholar] [CrossRef]
- Huang, C.; Nie, F.; Qin, Z.; Li, B.; Zhao, X. A snapshot of gene expression signatures generated using microarray datasets associated with excessive scarring. Am. J. Dermatopathol. 2013, 35, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Borzykh, O.B.; Petrova, M.M.; Shnayder, N.A.; Nasyrova, R.F. Problems of implementation of personalized medicine in medical cosmetology in Russia. Sib. Med. Rev. 2021, 2, 12–22. [Google Scholar] [CrossRef]
- Poetschke, J.; Gauglitz, G.G. Current options for the treatment of pathological scarring. JDDG J. Dtsch. Dermatol. Ges 2016, 14, 467–477. [Google Scholar] [CrossRef]
- Huguier, V.; Giot, J.-P.; Simonneau, M.; Levillain, P.; Charreau, S.; Garcia, M.; Jégou, J.-F.; Bodet, C.; Morel, F.; Lecron, J.-C. Oncostatin M exerts a protective effect against excessive scarring by counteracting the inductive effect of TGFβ1 on fibrosis markers. Sci. Rep. 2019, 9, 2113. [Google Scholar] [CrossRef]
- Deppermann, C.; Cherpokova, D.; Nurden, P.; Schulz, J.N.; Thielmann, I.; Kraft, P.; Vögtle, T.; Kleinschnitz, C.; Dütting, S.; Krohne, G.; et al. Gray platelet syndrome and defective thrombo-inflammation in Nbeal2-deficient mice. J. Clin. Investig. 2014, 124, 237–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eming, S.A.; Koch, M.; Krieger, A.; Brachvogel, B.; Kreft, S.; Bruckner-Tuderman, L.; Krieg, T.; Shannon, J.D.; Fox, J.W. Differential proteomic analysis distinguishes tissue repair biomarker signatures in wound exudates obtained from normal healing and chronic wounds. J. Proteome Res. 2010, 9, 4758–4766. [Google Scholar] [CrossRef]
- Sindrilaru, A.; Peters, T.; Wieschalka, S.; Baican, C.; Baican, A.; Peter, H.; Hainzl, A.; Schatz, S.; Qi, Y.; Schlecht, A.; et al. An unrestrained proinflammatory M1 macrophage population induced by iron impairs wound healing in humans and mice. J. Clin. Investig. 2011, 121, 985–997. [Google Scholar] [CrossRef] [Green Version]
- Stojadinovic, O.; Minkiewicz, J.; Sawaya, A.; Bourne, J.W.; Torzilli, P.; de Rivero Vaccari, J.P.; Dietrich, W.D.; Keane, R.W.; Tomic-Canic, M. Deep tissue injury in development of pressure ulcers: A decrease of inflammasome activation and changes in human skin morphology in response to aging and mechanical load. PLoS ONE 2013, 8, e69223. [Google Scholar] [CrossRef] [Green Version]
- Eming, S.A.; Wynn, T.A.; Martin, P. Inflammation and metabolism in tissue repair and regeneration. Science 2017, 356, 1026–1030. [Google Scholar] [CrossRef] [Green Version]
- Wilgus, T.A. Inflammation as an orchestrator of cutaneous scar formation: A review of the literature. Plast. Aesthet. Res. 2020, 7, 54. [Google Scholar] [CrossRef] [PubMed]
- Curaj, A.; Schumacher, D.; Rusu, M.; Staudt, M.; Li, X.; Simsekyilmaz, S.; Jankowski, V.; Jankowski, J.; Dumitraşcu, A.R.; Hausenloy, D.J. Neutrophils Modulate Fibroblast Function and Promote Healing and Scar Formation after Murine Myocardial Infarction. Int. J. Mol. Sci. 2020, 21, 3685. [Google Scholar] [CrossRef]
- Lucas, T.; Waisman, A.; Ranjan, R.; Roes, J.; Krieg, T.; Müller, W.; Roers, A.; Eming, S.A. Differential roles of macrophages in diverse phases of skin repair. J. Immunol. 2010, 184, 3964–3977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oishi, Y.; Manabe, I. Macrophages in inflammation, repair and regeneration. Int. Immunol. 2018, 30, 511–528. [Google Scholar] [CrossRef] [PubMed]
- Ha, D.H.; Kim, H.K.; Lee, J.; Kwon, H.H.; Park, G.H.; Yang, S.H.; Jung, J.Y.; Choi, H.; Lee, J.H.; Sung, S.; et al. Mesenchymal Stem/Stromal Cell-Derived Exosomes for Immunomodulatory Therapeutics and Skin Regeneration. Cells 2020, 9, 1157. [Google Scholar] [CrossRef]
- Antsiferova, M.; Martin, C.; Huber, M.; Feyerabend, T.B.; Förster, A.; Hartmann, K.; Rodewald, H.R.; Hohl, D.; Werner, S. Mast cells are dispensable for normal and activin-promoted wound healing and skin carcinogenesis. J. Immunol. 2013, 191, 6147–6155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willenborg, S.; Eckes, B.; Brinckmann, J.; Krieg, T.; Waisman, A.; Hartmann, K.; Roers, A.; Eming, S.A. Genetic ablation of mast cells redefines the role of mast cells in skin wound healing and bleomycin-induced fibrosis. J. Investig. Dermatol. 2014, 134, 2005–2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, M.; Kosaric, N.; Bonham, C.A.; Gurtner, G.C. Wound Healing: A Cellular Perspective. Physiol. Rev. 2019, 99, 665–706. [Google Scholar] [CrossRef] [PubMed]
- Wilgus, T.A.; Wulff, B.C. The Importance of Mast Cells in Dermal Scarring. Adv. Wound Care 2014, 3, 356–365. [Google Scholar] [CrossRef] [Green Version]
- Casado-Díaz, A.; Quesada-Gómez, J.M.; Dorado, G. Extracellular Vesicles Derived From Mesenchymal Stem Cells (MSC) in Regenerative Medicine: Applications in Skin Wound Healing. Front. Bioeng. Biotechnol. 2020, 8, 146. [Google Scholar] [CrossRef] [Green Version]
- El Ayadi, A.; Jay, J.W.; Prasai, A. Current Approaches Targeting the Wound Healing Phases to Attenuate Fibrosis and Scarring. Int. J. Mol. Sci. 2020, 21, 1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karppinen, S.M.; Heljasvaara, R.; Gullberg, D.; Tasanen, K.; Pihlajaniemi, T. Toward understanding scarless skin wound healing and pathological scarring. F1000Research 2019, 8, F1000. [Google Scholar] [CrossRef] [Green Version]
- Jourdan, M.; Madfes, D.C.; Lima, E.; Tian, Y.; Seité, S. Skin Care Management For Medical And Aesthetic Procedures To Prevent Scarring. Clin. Cosmet. Investig. Dermatol. 2019, 12, 799–804. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, G.V.; Hawkins, H.K.; Chinkes, D.; Burke, A.; Tavares, A.L.; Ramos-e-Silva, M.; Albrecht, T.B.; Kitten, G.T.; Herndon, D.N. Hypertrophic versus non hypertrophic scars compared by immunohistochemistry and laser confocal microscopy: Type I and III collagens. Int. Wound J. 2009, 6, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Tottoli, E.M.; Dorati, R.; Genta, I.; Chiesa, E.; Pisani, S.; Conti, B. Skin Wound Healing Process and New Emerging Technologies for Skin Wound Care and Regeneration. Pharmaceutics 2020, 12, 735. [Google Scholar] [CrossRef]
- Xue, M.; Jackson, C.J. Extracellular Matrix Reorganization During Wound Healing and Its Impact on Abnormal Scarring. Adv. Wound Care 2015, 4, 119–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, S.L.; Sheppard, D.; Duffield, J.S.; Violette, S. Therapy for fibrotic diseases: Nearing the starting line. Sci. Transl. Med. 2013, 5, 167sr161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gauglitz, G.G.; Korting, H.C.; Pavicic, T.; Ruzicka, T.; Jeschke, M.G. Hypertrophic scarring and keloids: Pathomechanisms and current and emerging treatment strategies. Mol. Med. 2011, 17, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Shirakami, E.; Yamakawa, S.; Hayashida, K. Strategies to prevent hypertrophic scar formation: A review of therapeutic interventions based on molecular evidence. Burn. Trauma 2020, 8, tkz003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoica, A.E.; Grumezescu, A.M.; Hermenean, A.O.; Andronescu, E.; Vasile, B.S. Scar-Free Healing: Current Concepts and Future Perspectives. Nanomaterials 2020, 10, 2179. [Google Scholar] [CrossRef]
- Ogawa, R. Surgery for scar revision and reduction: From primary closure to flap surgery. Burn. Trauma 2019, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Pastar, I.; Khan, A.A.; Stojadinovic, O.; Lebrun, E.A.; Medina, M.C.; Brem, H.; Kirsner, R.S.; Jimenez, J.J.; Leslie, C.; Tomic-Canic, M. Induction of specific microRNAs inhibits cutaneous wound healing. J. Biol. Chem. 2012, 287, 29324–29335. [Google Scholar] [CrossRef] [Green Version]
- Yates, C.C.; Bodnar, R.; Wells, A. Matrix control of scarring. Cell. Mol. Life Sci. 2011, 68, 1871–1881. [Google Scholar] [CrossRef] [Green Version]
- Liarte, S.; Bernabé-García, Á.; Nicolás, F.J. Role of TGF-β in Skin Chronic Wounds: A Keratinocyte Perspective. Cells 2020, 9, 306. [Google Scholar] [CrossRef] [Green Version]
- Bateman, J.F.; Boot-Handford, R.P.; Lamandé, S.R. Genetic diseases of connective tissues: Cellular and extracellular effects of ECM mutations. Nat. Rev. Genet. 2009, 10, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Tracy, L.E.; Minasian, R.A.; Caterson, E.J. Extracellular Matrix and Dermal Fibroblast Function in the Healing Wound. Adv. Wound Care 2016, 5, 119–136. [Google Scholar] [CrossRef]
- Schnittert, J.; Bansal, R.; Storm, G.; Prakash, J. Integrins in wound healing, fibrosis and tumor stroma: High potential targets for therapeutics and drug delivery. Adv. Drug Deliv. Rev. 2018, 129, 37–53. [Google Scholar] [CrossRef]
- Ferguson, M.W.; O’Kane, S. Scar-free healing: From embryonic mechanisms to adult therapeutic intervention. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2004, 359, 839–850. [Google Scholar] [CrossRef] [Green Version]
- Monavarian, M.; Kader, S.; Moeinzadeh, S.; Jabbari, E. Regenerative Scar-Free Skin Wound Healing. Tissue Eng. Part B Rev. 2019, 25, 294–311. [Google Scholar] [CrossRef] [PubMed]
- Usansky, I.; Jaworska, P.; Asti, L.; Kenny, F.N.; Hobbs, C.; Sofra, V.; Song, H.; Logan, M.; Graham, A.; Shaw, T.J. A developmental basis for the anatomical diversity of dermis in homeostasis and wound repair. J. Pathol. 2021, 253, 315–325. [Google Scholar] [CrossRef]
- Arseni, L.; Lombardi, A.; Orioli, D. From Structure to Phenotype: Impact of Collagen Alterations on Human Health. Int. J. Mol. Sci. 2018, 19, 1407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prodromidou, A.; Frountzas, M.; Vlachos, D.E.; Vlachos, G.D.; Bakoyiannis, I.; Perrea, D.; Pergialiotis, V. Botulinum toxin for the prevention and healing of wound scars: A systematic review of the literature. Plast. Surg. 2015, 23, 260–264. [Google Scholar] [CrossRef] [Green Version]
- Cramer, M.C.; Badylak, S.F. Extracellular Matrix-Based Biomaterials and Their Influence Upon Cell Behavior. Ann. Biomed. Eng. 2020, 48, 2132–2153. [Google Scholar] [CrossRef]
- Manka, S.W.; Bihan, D.; Farndale, R.W. Structural studies of the MMP-3 interaction with triple-helical collagen introduce new roles for the enzyme in tissue remodelling. Sci. Rep. 2019, 9, 18785. [Google Scholar] [CrossRef] [Green Version]
- Jansen, K.A.; Licup, A.J.; Sharma, A.; Rens, R.; MacKintosh, F.C.; Koenderink, G.H. The Role of Network Architecture in Collagen Mechanics. Biophys. J. 2018, 114, 2665–2678. [Google Scholar] [CrossRef] [Green Version]
- Potekaev, N.N.; Borzykh, O.B.; Medvedev, G.V.; Petrova, M.M.; Gavrilyuk, O.A.; Karpova, E.I.; Trefilova, V.V.; Demina, O.M.; Popova, T.E.; Shnayder, N.A. Genetic and Epigenetic Aspects of Skin Collagen Fiber Turnover and Functioning. Cosmetics 2021, 8, 92. [Google Scholar] [CrossRef]
- Raghunathan, R.; Sethi, M.K.; Klein, J.A.; Zaia, J. Proteomics, Glycomics, and Glycoproteomics of Matrisome Molecules. Mol. Cell Proteom. 2019, 18, 2138–2148. [Google Scholar] [CrossRef]
- Bayer, I.S. Hyaluronic Acid and Controlled Release: A Review. Molecules 2020, 25, 2649. [Google Scholar] [CrossRef] [PubMed]
- Shnayder, N.A.; Dyuzhakova, A.V.; Vaiman, E.E.; Nikitina, E.I.; Borzykh, O.B.; Nasyrova, R.F. The role of genetic factors of endogenous hyaluronic acid metabolism in maintaining skin homeostasis. Vestn. Dermatol. Venerol. 2021, 97, 24–38. [Google Scholar]
- Olmos-Zuñiga, J.R.; Baltazares-Lipp, M.; Hernández-Jiménez, C.; Jasso-Victoria, R.; Gaxiola-Gaxiola, M.; Silva-Martínez, M.; Iñiguez-García, M.A.; González-González, A.I.; Vázquez-Minero, J.C.; Luna-Flores, A.; et al. Treatment with Hyaluronic Acid and Collagen-Polyvinylpyrrolidone Improves Extracellular Matrix Assembly for Scarring after Tracheal Resection. Biomed. Res. Int. 2020, 2020, 3964518. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Naranjo, J.D.; Londono, R.; Badylak, S.F. Biologic Scaffolds. Cold Spring Harb. Perspect. Med. 2017, 7, a025676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartman, C.D.; Isenberg, B.C.; Chua, S.G.; Wong, J.Y. Extracellular matrix type modulates cell migration on mechanical gradients. Exp. Cell Res. 2017, 359, 361–366. [Google Scholar] [CrossRef]
- Rippa, A.L.; Kalabusheva, E.P.; Vorotelyak, E.A. Regeneration of Dermis: Scarring and Cells Involved. Cells 2019, 8, 607. [Google Scholar] [CrossRef] [Green Version]
- Govindaraju, P.; Todd, L.; Shetye, S.; Monslow, J.; Puré, E. CD44-dependent inflammation, fibrogenesis, and collagenolysis regulates extracellular matrix remodeling and tensile strength during cutaneous wound healing. Matrix Biol. 2019, 75–76, 314–330. [Google Scholar] [CrossRef] [PubMed]
- Rose, K.W.J.; Taye, N.; Karoulias, S.Z.; Hubmacher, D. Regulation of ADAMTS Proteases. Front. Mol. Biosci. 2021, 8, 701959. [Google Scholar] [CrossRef] [PubMed]
- Cui, N.; Hu, M.; Khalil, R.A. Biochemical and Biological Attributes of Matrix Metalloproteinases. Prog. Mol. Biol. Transl. Sci. 2017, 147, 1–73. [Google Scholar]
- Wells, A.; Nuschke, A.; Yates, C.C. Skin tissue repair: Matrix microenvironmental influences. Matrix Biol. 2016, 49, 25–36. [Google Scholar] [CrossRef] [Green Version]
- Weisel, J.W.; Litvinov, R.I. Fibrin Formation, Structure and Properties. Subcell. Biochem. 2017, 82, 405–456. [Google Scholar]
- Graham, J.; Raghunath, M.; Vogel, V. Fibrillar fibronectin plays a key role as nucleator of collagen I polymerization during macromolecular crowding-enhanced matrix assembly. Biomater. Sci. 2019, 7, 4519–4535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, M.; Ji, Y.; Luo, Y.; Li, R.; Fay, W.P.; Wu, J. Plasminogen activator inhibitor-1 regulates the vascular expression of vitronectin. J. Thromb. Haemost. 2017, 15, 2451–2460. [Google Scholar] [CrossRef] [Green Version]
- Seifert, A.W.; Monaghan, J.R.; Voss, S.R.; Maden, M. Skin regeneration in adult axolotls: A blueprint for scar-free healing in vertebrates. PLoS ONE 2012, 7, e32875. [Google Scholar] [CrossRef] [Green Version]
- Kavasi, R.M.; Berdiaki, A.; Spyridaki, I. HA metabolism in skin homeostasis and inflammatory disease. Food Chem. Toxicol. 2017, 101, 128. [Google Scholar] [CrossRef]
- Leung, A.; Crombleholme, T.M.; Keswani, S.G. Fetal wound healing: Implications for minimal scar formation. Curr. Opin. Pediatr. 2012, 24, 371. [Google Scholar] [CrossRef] [Green Version]
- Vitulo, N.; Dalla Valle, L.; Skobo, T.; Valle, G.; Alibardi, L. Transcriptome analysis of the regenerating tail vs. the scarring limb in lizard reveals pathways leading to successful vs unsuccessful organ regeneration in amniotes. Dev. Dyn. 2017, 246, 116. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, X.H.; Panetta, N.J.; Talbott, M.D. Hyaluronic acid synthesis is required for zebrafish tail fin regeneration. PLoS ONE 2017, 12, e0171898. [Google Scholar] [CrossRef]
- Abdelaziz Mohamed, I.; Gadeau, A.P.; Hasan, A.; Abdulrahman, N.; Mraiche, F. Osteopontin: A Promising Therapeutic Target in Cardiac Fibrosis. Cells 2019, 8, 1558. [Google Scholar] [CrossRef] [Green Version]
- Kaasbøll, O.J.; Gadicherla, A.K.; Wang, J.H.; Monsen, V.T.; Hagelin, E.M.V.; Dong, M.Q.; Attramadal, H. Connective tissue growth factor (CCN2) is a matricellular preproprotein controlled by proteolytic activation. J. Biol. Chem. 2018, 293, 17953–17970. [Google Scholar] [CrossRef] [Green Version]
- Spada, S.; Tocci, A.; Di Modugno, F.; Nisticò, P. Fibronectin as a multiregulatory molecule crucial in tumor matrisome: From structural and functional features to clinical practice in oncology. J. Exp. Clin. Cancer Res. 2021, 40, 102. [Google Scholar] [CrossRef]
- Musiime, M.; Chang, J.; Hansen, U.; Kadler, K.E.; Zeltz, C.; Gullberg, D. Collagen Assembly at the Cell Surface: Dogmas Revisited. Cells 2021, 10, 662. [Google Scholar] [CrossRef] [PubMed]
- Karna, S.R.; D’Arpa, P.; Chen, T.; Qian, L.W.; Fourcaudot, A.B.; Yamane, K.; Chen, P.; Abercrombie, J.J.; You, T.; Leung, K.P. RNA-Seq transcriptomic responses of full-thickness dermal excision wounds to Pseudomonas aeruginosa acute and biofilm infection. PLoS ONE 2016, 11, e016531. [Google Scholar] [CrossRef] [Green Version]
- Akbas, F.; Ozdemir, B.; Bahtiyar, N.; Arkan, H.; Onaran, I. Platelet-rich plasma and platelet-derived lipid factors induce different and similar gene expression responses for selected genes related to wound healing in rat dermal wound environment. Mol. Biol. Res. Commun. 2020, 9, 145–153. [Google Scholar]
- Eming, S.A.; Martin, P.; Tomic-Canic, M. Wound repair and regeneration: Mechanisms, signaling, and translation. Sci. Transl. Med. 2014, 6, 265sr6. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Wu, H.; Byrne, M.; Krane, S.; Jaenisch, R. Type III collagen is crucial for collagen I fibrillogenesis and for normal cardiovascular development. Proc. Natl. Acad. Sci. USA 1997, 94, 1852–1856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Germain, D.P. Ehlers-Danlos syndrome type IV. Orphanet J. Rare Dis. 2007, 2, 32. [Google Scholar] [CrossRef]
- Volk, S.W.; Wang, Y.; Mauldin, E.A.; Liechty, K.W.; Adams, S.L. Diminished type III collagen promotes myofibroblast differentiation and increases scar deposition in cutaneous wound healing. Cells Tissues Organs 2011, 194, 25–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuttle, L.; Nataatmadja, M.; Fraser, J.F.; Kempf, M.; Kimble, R.M.; Hayes, M.T. Collagen in the scarless fetal skin wound: Detection with Picrosirius-polarization. Wound Repair. Regener. 2005, 13, 198. [Google Scholar] [CrossRef] [Green Version]
- Stone, R.C.; Chen, V.; Burgess, J.; Pannu, S.; Tomic-Canic, M. Genomics of Human Fibrotic Diseases: Disordered Wound Healing Response. Int. J. Mol. Sci. 2020, 21, 8590. [Google Scholar] [CrossRef]
- Habashi, J.P.; Judge, D.P.; Holm, T.M.; Cohn, R.D.; Loeys, B.L.; Cooper, T.K.; Myers, L.; Klein, E.C.; Liu, G.; Calvi, C.; et al. Losartan, an AT1 antagonist, prevents aortic aneurysm in a mouse model of Marfan syndrome. Science 2006, 312, 117–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martino, M.M.; Tortelli, F.; Mochizuki, M.; Traub, S.; Ben-David, D.; Kuhn, G.A.; Müller, R.; Livne, E.; Eming, S.A.; Hubbell, J.A. Engineering the growth factor microenvironment with fibronectin domains to promote wound and bone tissue healing. Sci. Transl. Med. 2011, 3, 100ra189. [Google Scholar] [CrossRef] [Green Version]
- Polat, E.; Aksöz, İ.; Arkan, H.; Coşkunpınar, E.; Akbaş, F.; Onaran, İ. Gene expression profiling of Lucilia sericata larvae extraction/secretion-treated skin wounds. Gene 2014, 550, 223–229. [Google Scholar] [CrossRef]
- Baumann, L.; Bernstein, E.F.; Weiss, A.S.; Bates, D.; Humphrey, S.; Silberberg, M.; Daniels, R. Clinical Relevance of Elastin in the Structure and Function of Skin. Aesthet. Surg. J. Open Forum 2021, 3, ojab019. [Google Scholar] [CrossRef]
- Cohen, B.E.; Geronemus, R.G.; McDaniel, D.H.; Brauer, J.A. The role of elastic fibers in scar formation and treatment. Dermatol. Surg. 2017, 43 (Suppl. 1), 19–24. [Google Scholar] [CrossRef]
- Bi, M.; Sun, P.; Li, D.; Dong, Z.; Chen, Z. Intralesional Injection of Botulinum Toxin Type A Compared with Intralesional Injection of Corticosteroid for the Treatment of Hypertrophic Scar and Keloid: A Systematic Review and Meta-Analysis. Med. Sci. Monit. 2019, 25, 2950–2958. [Google Scholar] [CrossRef]
- Murdock, J.; Sayed, M.S.; Tavakoli, M.; Portaliou, D.M.; Lee, W.W. Safety and efficacy of a growth factor and cytokine-containing topical product in wound healing and incision scar management after upper eyelid blepharoplasty: A prospective split-face study. Clin. Ophthalmol. 2016, 10, 1223–1228. [Google Scholar]
- Wang, J.; Liao, Y.; Xia, J.; Wang, Z.; Mo, X.; Feng, J.; He, Y.; Chen, X.; Li, Y.; Lu, F.; et al. Mechanical micronization of lipoaspirates for the treatment of hypertrophic scars. Stem Cell Res. Ther. 2019, 10, 42. [Google Scholar] [CrossRef] [PubMed]
- Urciuolo, F.; Casale, C.; Imparato, G.; Netti, P.A. Bioengineered Skin Substitutes: The Role of Extracellular Matrix and Vascularization in the Healing of Deep Wounds. J. Clin. Med. 2019, 8, 2083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, S.S.; Dotor, J.; Hontanilla, B. Effect of P144® (Anti-TGF-β) in an “In Vivo” Human Hypertrophic Scar Model in Nude Mice. PLoS ONE 2015, 10, e0144489. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Zhao, T.; Xie, X.; Yu, D.; Wu, L.; Yu, W.; Sun, W. MicroRNA-663 regulates the proliferation of fibroblasts in hypertrophic scars via transforming growth factor-β1. Exp. Ther. Med. 2018, 16, 1311–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Potekaev, N.N.; Borzykh, O.B.; Medvedev, G.V.; Pushkin, D.V.; Petrova, M.M.; Petrov, A.V.; Dmitrenko, D.V.; Karpova, E.I.; Demina, O.M.; Shnayder, N.A. The Role of Extracellular Matrix in Skin Wound Healing. J. Clin. Med. 2021, 10, 5947. https://doi.org/10.3390/jcm10245947
Potekaev NN, Borzykh OB, Medvedev GV, Pushkin DV, Petrova MM, Petrov AV, Dmitrenko DV, Karpova EI, Demina OM, Shnayder NA. The Role of Extracellular Matrix in Skin Wound Healing. Journal of Clinical Medicine. 2021; 10(24):5947. https://doi.org/10.3390/jcm10245947
Chicago/Turabian StylePotekaev, Nikolai N., Olga B. Borzykh, German V. Medvedev, Denis V. Pushkin, Marina M. Petrova, Artem V. Petrov, Diana V. Dmitrenko, Elena I. Karpova, Olga M. Demina, and Natalia A. Shnayder. 2021. "The Role of Extracellular Matrix in Skin Wound Healing" Journal of Clinical Medicine 10, no. 24: 5947. https://doi.org/10.3390/jcm10245947
APA StylePotekaev, N. N., Borzykh, O. B., Medvedev, G. V., Pushkin, D. V., Petrova, M. M., Petrov, A. V., Dmitrenko, D. V., Karpova, E. I., Demina, O. M., & Shnayder, N. A. (2021). The Role of Extracellular Matrix in Skin Wound Healing. Journal of Clinical Medicine, 10(24), 5947. https://doi.org/10.3390/jcm10245947