
Ethnic Differences in Serum Levels of microRNAs Potentially Regulating Alcohol Dehydrogenase 1B and Aldehyde Dehydrogenase 2


 ,
, 

Abstract
:1. Introduction
2. Subjects and Methods
2.1. Subjects
2.2. Collection of Blood Sample
2.3. RNA Extraction and miRNA Expression Profiling
2.4. Bioinformatics Survey on miRNAs
2.5. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Crabb, D.W.; Edenberg, H.J.; Bosron, W.F.; Li, T.K. Genotypes for aldehyde dehydrogenase deficiency and alcohol sensitivity. The inactive ALDH2(2) allele is dominant. J. Clin. Investig. 1989, 83, 314–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, A.W. Racial differences in alcohol sensitivity. Alcohol Alcohol. 1986, 21, 93–104. [Google Scholar] [PubMed]
- Brooks, P.J.; Enoch, M.; Goldman, D.; Li, T.; Yokoyama, A. The alcohol flushing response: An unrecognized risk factor for esophageal cancer from alcohol consumption. PLoS Med. 2009, 6, e50. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.H.; Ferreira, J.C.B.; Mochly-Rosen, D. ALDH2 and Cardiovascular Disease. Adv. Exp. Med. Biol. 2019, 1193, 53–67. [Google Scholar] [PubMed]
- Voulgaridou, G.P.; Anestopoulos, I.; Franco, R.; Panayiotidis, M.I.; Pappa, A. DNA damage induced by endogenous aldehydes: Current state of knowledge. Mutat. Res. 2011, 711, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Edenberg, H.J.; McClintick, J.N. Alcohol dehydrogenases, aldehyde dehydrogenases, and alcohol use disorders: A critical review. Alcohol Clin. Exp. Res. 2018, 42, 2281–2297. [Google Scholar] [CrossRef] [PubMed]
- Takeshita, T.; Yang, X.; Morimoto, K. Association of the ADH2 genotypes with skin responses after ethanol exposure in Japanese male university students. Alcohol Clin. Exp. Res. 2001, 25, 1264–1269. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.; Zuo, M.; Hu, G.; Duan, X.; Yang, Z. mir-193 targets ALDH2 and contributes to toxic aldehyde accumulation and tyrosine hydroxylase dysfunction in cerebral ischemia/reperfusion injury. Oncotarget 2017, 8, 99681–99692. [Google Scholar] [CrossRef] [Green Version]
- Shen, B.; Pan, Q.; Yang, Y.; Gao, Y.; Liu, X.; Li, W.; Han, Y.; Yuan, X.; Qu, Y.; Zhao, Z. miR-224 affects mammary epithelial cell apoptosis and triglyceride production by downregulating ACADM and ALDH2 genes. DNA Cell Biol. 2017, 36, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Li, S.P.; Liu, B.; Song, B.; Wang, C.X.; Zhou, Y.C. miR-28 promotes cardiac ischemia by targeting mitochondrial aldehyde dehydrogenase 2 (ALDH2) in mus musculus cardiac myocytes. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 752–758. [Google Scholar] [PubMed]
- Wang, Z.; Song, J.; Zhang, L.; Huang, S.; Bao, L.; Chen, F.; Zhao, X. Increased expression of microRNA-378a-5p in acute ethanol exposure of rat cardiomyocytes. Cell Stress Chaperones 2017, 22, 245–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, J.; Zhou, L.; Xie, Q.F.; Xie, H.Y.; Wei, X.Y.; Gao, F.; Xing, C.Y.; Xu, X.; Li, L.J.; Zheng, S.S. The impact of miR-34a on protein output in hepatocellular carcinoma HepG2 cells. Proteomics 2010, 10, 1557–1572. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, Y.; Luo, J.; Xu, L.; Li, X.; Jin, Y.; Li, C.; Feng, M.; Wang, Y.; Chen, J.; et al. MicroRNA hsa-miR-1301-3p regulates human ADH6, ALDH5A1 and ALDH8A1 in the ethanol-acetaldehyde-acetate metabolic pathway. Mol. Pharmacol. 2020, 98, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Hou, Y.; Ma, W.; Xie, M.; Jin, Y.; Xu, L.; Li, C.; Wang, Y.; Chen, J.; Chen, W.; et al. A novel mechanism underlying alcohol dehydrogenase expression: Hsa-miR-148a-3p promotes ADH4 expression via an AGO1-dependent manner in control and ethanol-exposed hepatic cells. Biochem. Pharmacol. 2021, 189, 114458. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, I.; Eguchi, R.; Sotoda, Y.; von Lewinski, D.; Sourij, H.; Daimon, T.; Groschner, K.; Rainer, P.P. Blood levels of microRNAs associated with ischemic heart disease differ between Austrians and Japanese: A pilot study. Sci. Rep. 2020, 10, 13628. [Google Scholar] [CrossRef] [PubMed]
- Storey, J.D.; Taylor, J.E.; Siegmund, D. Strong control, conservative point estimation and simultaneous conservative consistency of false discovery rates: A unified approach. J. R. Statist. Soc. Ser. 2004, B 66, 187–205. [Google Scholar] [CrossRef]

{kind=link}
| miRNA | Regulation * | Material | Species | Reference | Comparison of Serum Level |
|---|---|---|---|---|---|
| miR-193 | down | PC-12 cell, brain tissue | rat | Mao et al., 2017 | q = 0.003 (miR-193a-5p) ** q = 0.100 (miR-193b-5p) n.d. (miR-193a-3p, miR-193b-3p) |
| miR-224 | down | mammary epithelial cell | bovine | Shen et al., 2017 | n.d. (miR-224-3p, miR-224-5p) |
| miR-28 | down | cardiomyocyte | mouse | Li et al., 2015 | n.d. (miR-28-3p, miR-28-5p) |
| miR-34a | down | HepG2 cell | human | Cheng et al., 2010 | n.d. (miR-34a-3p, miR-34a-5p) |
| miR-378a-5p | down | cardiomyocyte | rat | Wang et al., 2017 | n.d. |
| miRNA | Target Score | Context Score | Fold Change * | q Value |
|---|---|---|---|---|
| (miRDB) | (TargetScan) | |||
| ALDH2 | ||||
| 30a-5p | 78 | −0.39 | n.d. | n.d. |
| 30b-5p | 78 | −0.40 | n.d. | n.d. |
| 30c-5p | 78 | −0.40 | n.d. | n.d. |
| 30d-5p | 78 | −0.39 | 1.21 | 0.111 |
| 30e-5p | 78 | −0.41 | n.d. | n.d. |
| 3942-3p | 90 | −0.39 | n.d. | n.d. |
| 3689a-5p | 65 | −0.30 | n.d. | n.d. |
| 3689e-5p | 65 ** | −0.30 | n.d. | n.d. |
| 4510 | 66 | −0.41 | n.d. | n.d. |
| 4753−3p | 80 | −0.34 | n.d. | n.d. |
| 4793-5p | 75 | −0.59 | n.d. | n.d. |
| 6127 | 66 | −0.41 | −1.04 | 0.272 |
| 6129 | 66 | −0.41 | n.d. | n.d. |
| 6130 | 66 | −0.43 | −1.11 | 0.197 |
| 6133 | 66 | −0.43 | −1.01 | 0.412 |
| 6809-3p | 94 | −0.37 | n.d. | n.d. |
| 6854-3p | 73 | −0.47 | n.d. | n.d. |
| 8065 | 80 | −0.60 | n.d. | n.d. |
| ADH1B | ||||
| 135b-3p | 68 | −0.41 | n.d. | n.d. |
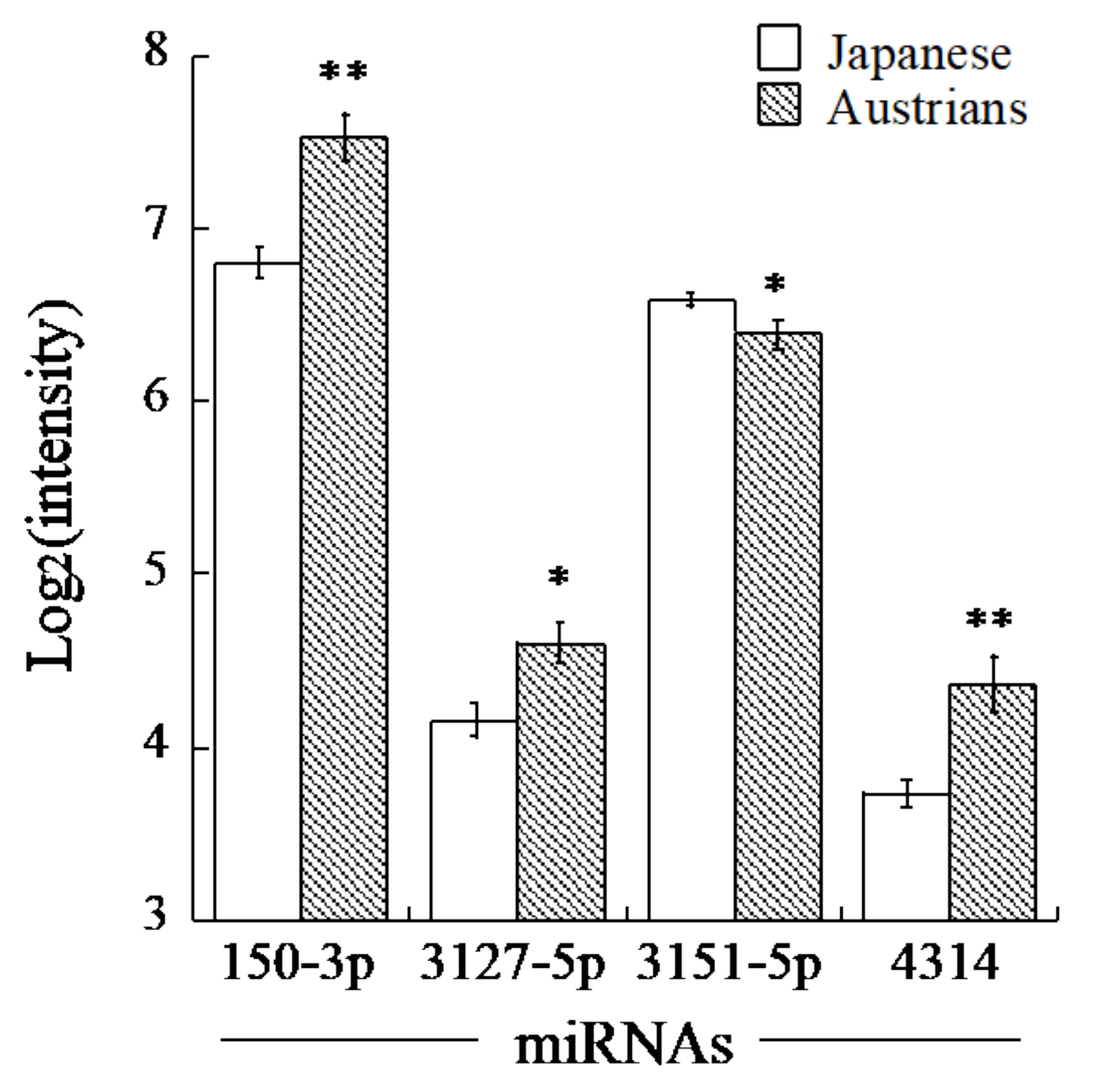
| 150-3p | 72 | −0.35 | 1.65 | <0.001 |
| 3127-5p | 71 | −0.32 | 1.36 | <0.005 |
| 3151-5p | 85 | −0.42 | −1.16 | <0.003 |
| 3943 | 88 | −0.34 | n.d. | n.d. |
| 4314 | 93 | −0.52 | 1.55 | <0.001 |
| 449b-3p | 60 | −0.34 | −1.02 | 0.278 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wakabayashi, I.; Sourij, H.; Sotoda, Y.; Daimon, T.; Groschner, K.; Rainer, P.P. Ethnic Differences in Serum Levels of microRNAs Potentially Regulating Alcohol Dehydrogenase 1B and Aldehyde Dehydrogenase 2. J. Clin. Med. 2021, 10, 3678. https://doi.org/10.3390/jcm10163678
Wakabayashi I, Sourij H, Sotoda Y, Daimon T, Groschner K, Rainer PP. Ethnic Differences in Serum Levels of microRNAs Potentially Regulating Alcohol Dehydrogenase 1B and Aldehyde Dehydrogenase 2. Journal of Clinical Medicine. 2021; 10(16):3678. https://doi.org/10.3390/jcm10163678
Chicago/Turabian StyleWakabayashi, Ichiro, Harald Sourij, Yoko Sotoda, Takashi Daimon, Klaus Groschner, and Peter P. Rainer. 2021. "Ethnic Differences in Serum Levels of microRNAs Potentially Regulating Alcohol Dehydrogenase 1B and Aldehyde Dehydrogenase 2" Journal of Clinical Medicine 10, no. 16: 3678. https://doi.org/10.3390/jcm10163678
APA StyleWakabayashi, I., Sourij, H., Sotoda, Y., Daimon, T., Groschner, K., & Rainer, P. P. (2021). Ethnic Differences in Serum Levels of microRNAs Potentially Regulating Alcohol Dehydrogenase 1B and Aldehyde Dehydrogenase 2. Journal of Clinical Medicine, 10(16), 3678. https://doi.org/10.3390/jcm10163678