

Fast-Rising Electric Pulses by Reducing Membrane Tension for Efficient Membrane Electroporation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
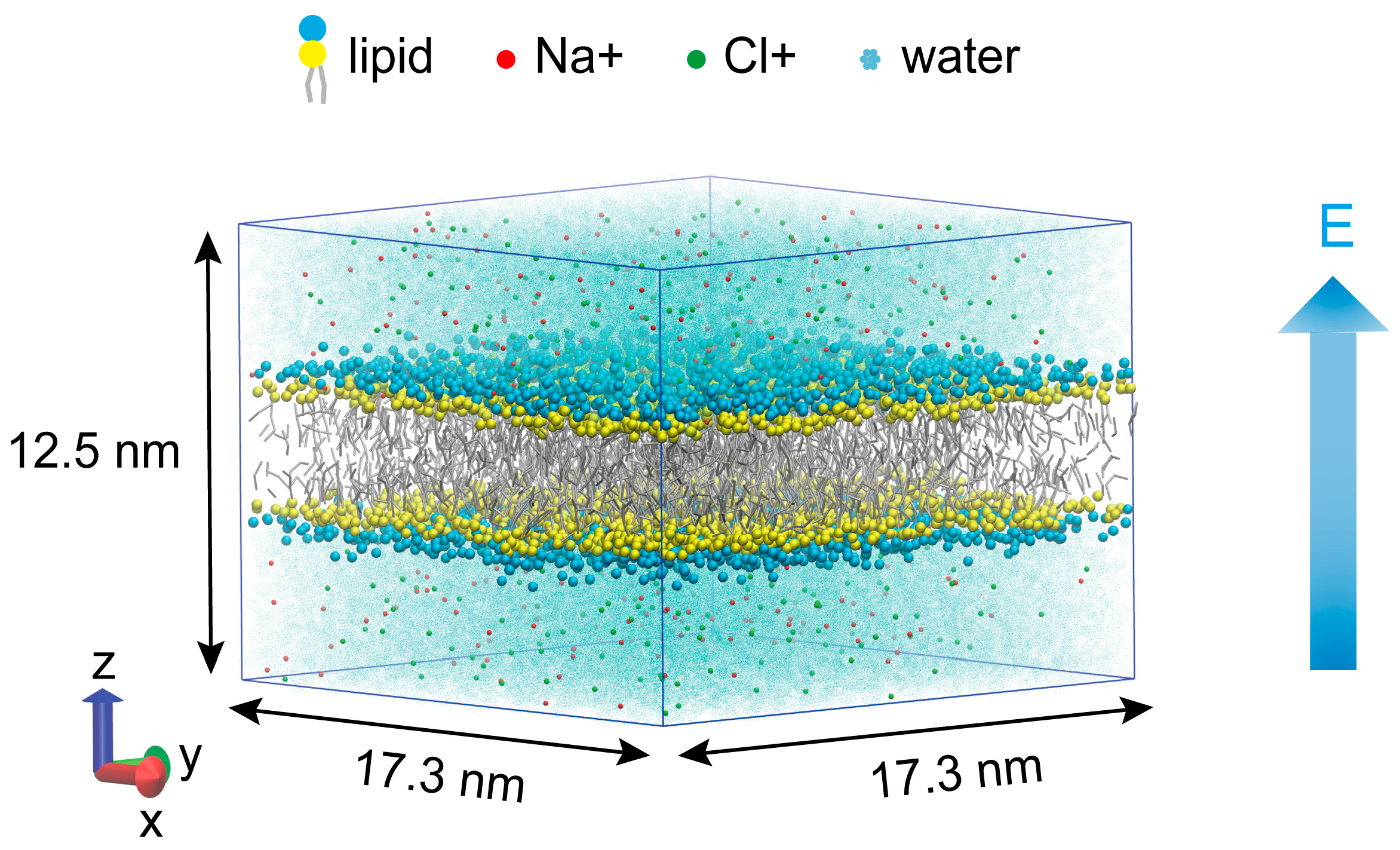
2. Materials and Methods
3. Results and Discussion
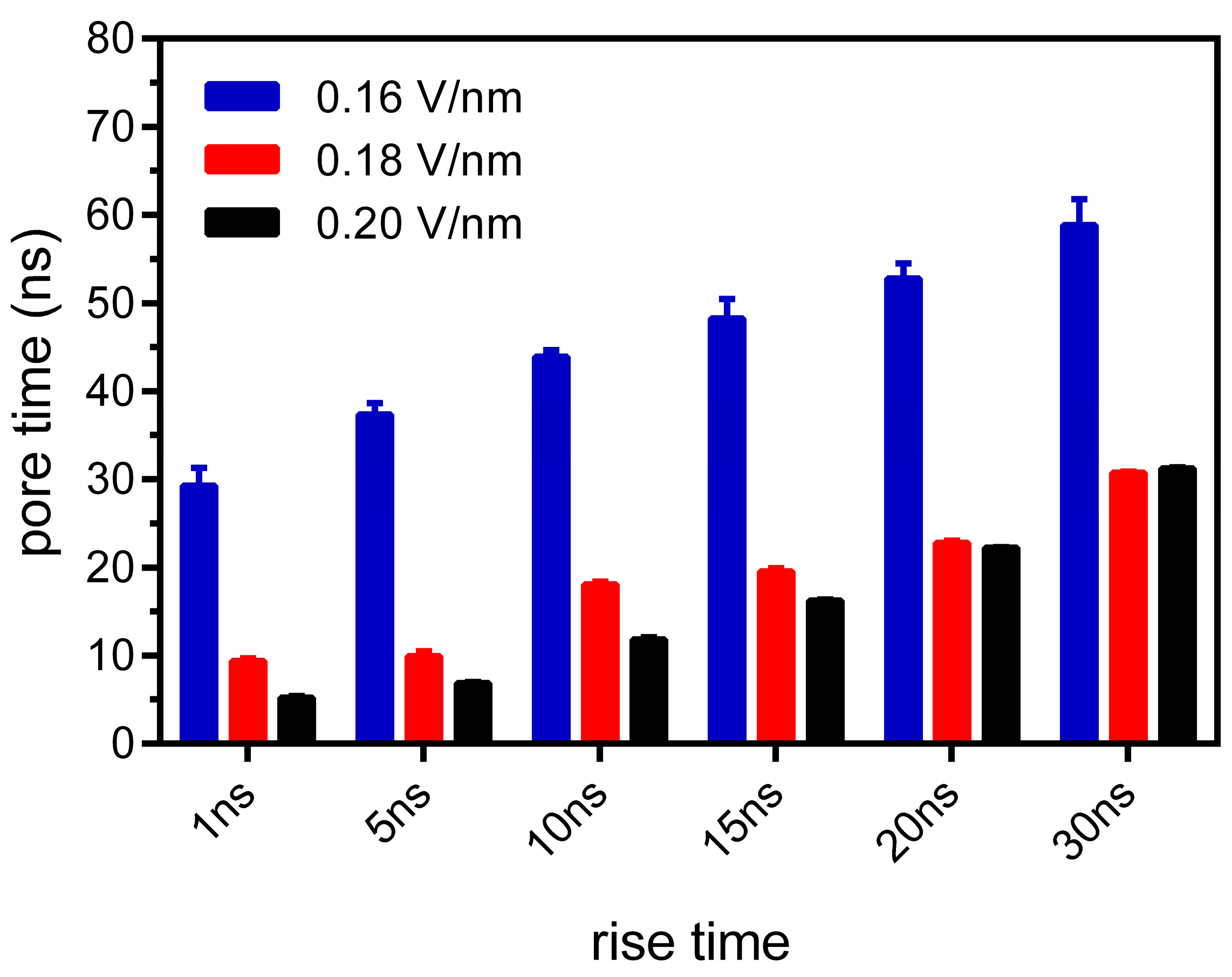
3.1. Fast-Rising Electric Pulses Promote Membrane Electroporation Effects
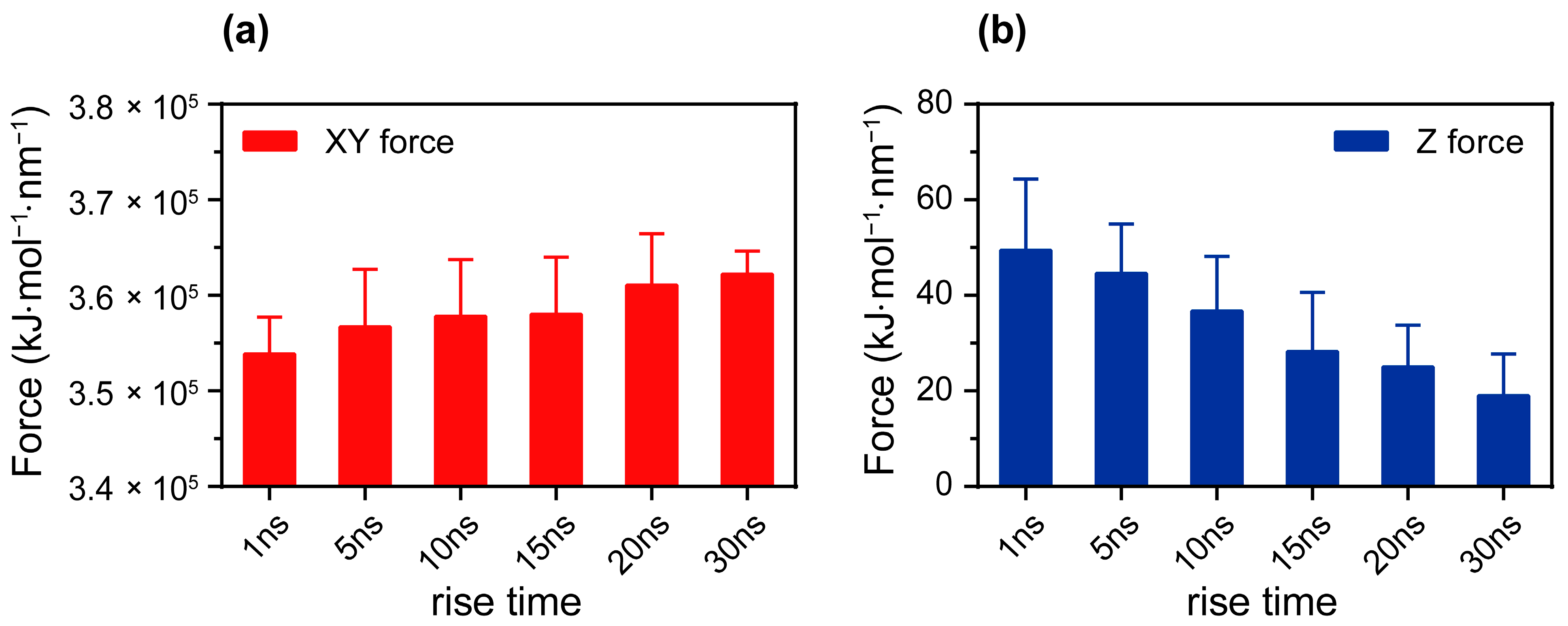
3.2. The Reverse Effect of Anisotropic Coulomb Forces Induced by Changing Electric Fields on Membranes
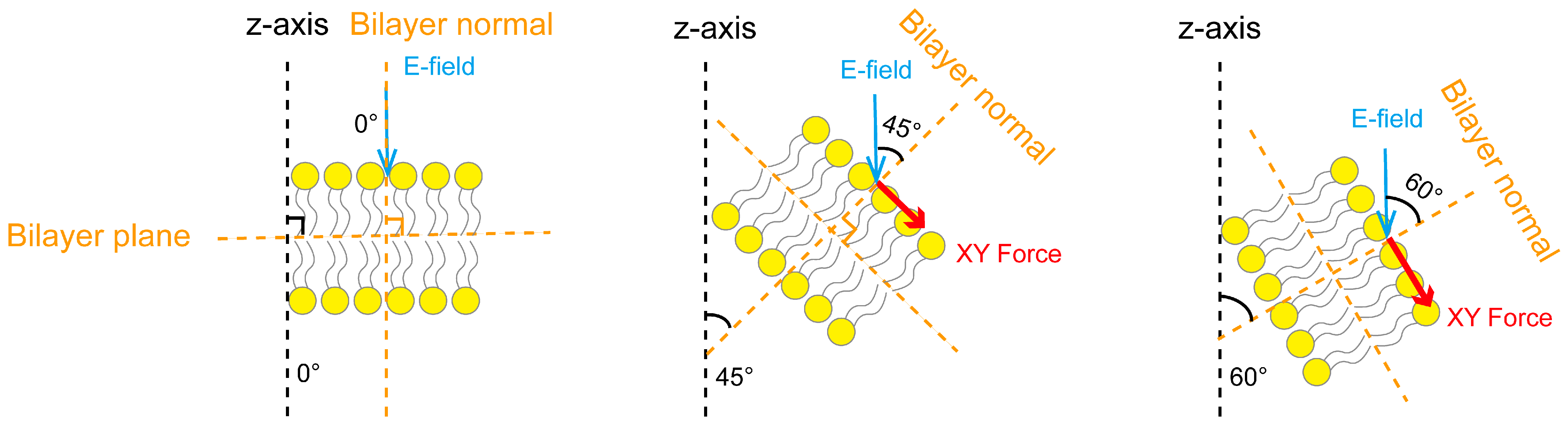
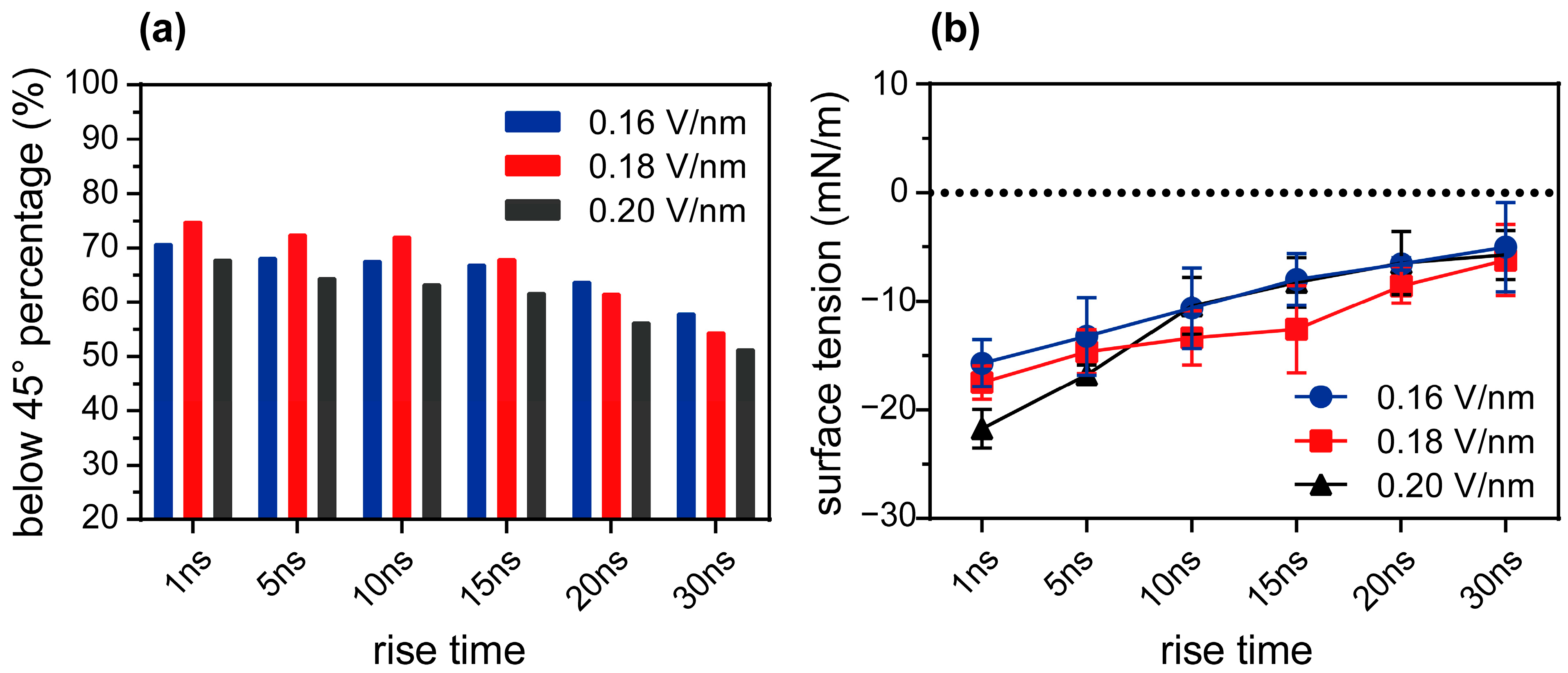
3.3. Dynamic Electric Field Regulation of Membrane Tension Related to the Electric Field Angle
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guo, S.; Jing, Y.; Burcus, N.I.; Lassiter, B.P.; Tanaz, R.; Heller, R.; Beebe, S.J. Nano-pulse stimulation induces potent immune responses, eradicating local breast cancer while reducing distant metastases. Int. J. Cancer 2018, 142, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Ringel-Scaia, V.M.; Beitel-White, N.; Lorenzo, M.F.; Brock, R.M.; Huie, K.E.; Coutermarsh-Ott, S.; Eden, K.; McDaniel, D.K.; Verbridge, S.S.; Rossmeisl, J.H., Jr.; et al. High-frequency irreversible electroporation is an effective tumor ablation strategy that induces immunologic cell death and promotes systemic anti-tumor immunity. EBioMedicine 2019, 44, 112–125. [Google Scholar] [CrossRef] [PubMed]
- Rossi, A.; Pakhomova, O.N.; Mollica, P.A.; Casciola, M.; Mangalanathan, U.; Pakhomov, A.G.; Muratori, C. Nanosecond Pulsed Electric Fields Induce Endoplasmic Reticulum Stress Accompanied by Immunogenic Cell Death in Murine Models of Lymphoma and Colorectal Cancer. Cancers 2019, 11, 2034. [Google Scholar] [CrossRef]
- Poompavai, S.; Sree, V.G.; Priyaa, A.K. Electrothermal Analysis of the Breast-Tumor Model During Electroporation. IEEE Trans. Radiat. Plasma Med. Sci. 2020, 4, 512–524. [Google Scholar] [CrossRef]
- Reddy, V.Y.; Dukkipati, S.R.; Neuzil, P.; Anic, A.; Petru, J.; Funasako, M.; Cochet, H.; Minami, K.; Breskovic, T.; Sikiric, I.; et al. Pulsed Field Ablation of Paroxysmal Atrial Fibrillation: 1-Year Outcomes of IMPULSE, PEFCAT, and PEFCAT II. JACC Clin. Electrophysiol. 2021, 7, 614–627. [Google Scholar] [CrossRef]
- Gudvangen, E.; Kim, V.; Novickij, V.; Battista, F.; Pakhomov, A.G. Electroporation and cell killing by milli- to nanosecond pulses and avoiding neuromuscular stimulation in cancer ablation. Sci. Rep. 2022, 12, 1763. [Google Scholar] [CrossRef]
- Miklavčič, D.; Verma, A.; Krahn, P.R.P.; Štublar, J.; Kos, B.; Escartin, T.; Lombergar, P.; Coulombe, N.; Terricabras, M.; Jarm, T.; et al. Biophysics and electrophysiology of pulsed field ablation in normal and infarcted porcine cardiac ventricular tissue. Sci. Rep. 2024, 14, 32063. [Google Scholar] [CrossRef]
- Kotnik, T.; Rems, L.; Tarek, M.; Miklavčič, D. Membrane Electroporation and Electropermeabilization: Mechanisms and Models. Annu. Rev. Biophys. 2019, 48, 63–91. [Google Scholar] [CrossRef]
- Peng, W.; Polajžer, T.; Yao, C.; Miklavčič, D. Dynamics of Cell Death Due to Electroporation Using Different Pulse Parameters as Revealed by Different Viability Assays. Ann. Biomed. Eng. 2024, 52, 22–35. [Google Scholar] [CrossRef]
- Polajžer, T.; Miklavčič, D. Immunogenic Cell Death in Electroporation-Based Therapies Depends on Pulse Waveform Characteristics. Vaccines 2023, 11, 1036. [Google Scholar] [CrossRef]
- Tang, J.; Yin, H.; Ma, J.; Bo, W.; Yang, Y.; Xu, J.; Liu, Y.; Gong, Y. Terahertz Electric Field-Induced Membrane Electroporation by Molecular Dynamics Simulations. J. Membr. Biol. 2018, 251, 681–693. [Google Scholar] [CrossRef] [PubMed]
- Akimov, S.A.; Volynsky, P.E.; Galimzyanov, T.R.; Kuzmin, P.I.; Pavlov, K.V.; Batishchev, O.V. Pore formation in lipid membrane II: Energy landscape under external stress. Sci. Rep. 2017, 7, 12509. [Google Scholar] [CrossRef] [PubMed]
- Akimov, S.A.; Volynsky, P.E.; Galimzyanov, T.R.; Kuzmin, P.I.; Pavlov, K.V.; Batishchev, O.V. Pore formation in lipid membrane I: Continuous reversible trajectory from intact bilayer through hydrophobic defect to transversal pore. Sci. Rep. 2017, 7, 12152. [Google Scholar] [CrossRef] [PubMed]
- Vasilkoski, Z.; Esser, A.T.; Gowrishankar, T.R.; Weaver, J.C. Membrane electroporation: The absolute rate equation and nanosecond time scale pore creation. Phys. Rev. E Stat. Nonlinear Soft Matter Phys. 2006, 74, 021904. [Google Scholar] [CrossRef]
- Schoenbach, K.H.; Beebe, S.J.; Buescher, E.S. Intracellular effect of ultrashort electrical pulses. Bioelectromagnetics 2001, 22, 440–448. [Google Scholar] [CrossRef]
- Beebe, S.J.; Chen, Y.J.; Sain, N.M.; Schoenbach, K.H.; Xiao, S. Transient features in nanosecond pulsed electric fields differentially modulate mitochondria and viability. PLoS ONE 2012, 7, e51349. [Google Scholar] [CrossRef]
- Jo, S.; Kim, T.; Iyer, V.G.; Im, W. CHARMM-GUI: A web-based graphical user interface for CHARMM. J. Comput. Chem. 2008, 29, 1859–1865. [Google Scholar] [CrossRef]
- Wu, E.L.; Cheng, X.; Jo, S.; Rui, H.; Song, K.C.; Dávila-Contreras, E.M.; Qi, Y.; Lee, J.; Monje-Galvan, V.; Venable, R.M.; et al. CHARMM-GUI Membrane Builder toward realistic biological membrane simulations. J. Comput. Chem. 2014, 35, 1997–2004. [Google Scholar] [CrossRef]
- Chen, L.; Chen, J.; Zhou, G.; Wang, Y.; Xu, C.; Wang, X. Molecular Dynamics Simulations of the Permeation of Bisphenol A and Pore Formation in a Lipid Membrane. Sci. Rep. 2016, 6, 33399. [Google Scholar] [CrossRef]
- Mou, Q.; Xu, M.; Deng, J.; Hu, N.; Yang, J. Studying the roles of salt ions in the pore initiation and closure stages in the biomembrane electroporation. APL Bioeng. 2023, 7, 026103. [Google Scholar] [CrossRef]
- Kasparyan, G.; Hub, J.S. Molecular simulations reveal the free energy landscape and transition state of membrane electroporation. Phys. Rev. Lett. 2024, 132, 148401. [Google Scholar] [CrossRef] [PubMed]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef]
- Marrink, S.J.; De Vries, A.H.; Mark, A.E. Coarse grained model for semiquantitative lipid simulations. J. Phys. Chem. B 2004, 108, 750–760. [Google Scholar] [CrossRef]
- Marrink, S.J.; Risselada, H.J.; Yefimov, S.; Tieleman, D.P.; De Vries, A.H. The MARTINI force field: Coarse grained model for biomolecular simulations. J. Phys. Chem. B 2007, 111, 7812–7824. [Google Scholar] [CrossRef] [PubMed]
- Wassenaar, T.A.; Ingólfsson, H.I.; Bockmann, R.A.; Tieleman, D.P.; Marrink, S.J. Computational lipidomics with insane: A versatile tool for generating custom membranes for molecular simulations. J. Chem. Theory Comput. 2015, 11, 2144–2155. [Google Scholar] [CrossRef]
- Berendsen, H.J.C. Transport Properties Computed by Linear Response through Weak Coupling to a Bath. In Computer Simulation in Materials Science: Interatomic Potentials, Simulation Techniques and Applications; Meyer, M., Pontikis, V., Eds.; Springer: Dordrecht, The Netherlands, 1991; pp. 139–155. [Google Scholar]
- Parrinello, M.; Rahman, A. Polymorphic transitions in single crystals: A new molecular dynamics method. J. Appl. Phys. 1981, 52, 7182–7190. [Google Scholar] [CrossRef]
- Darden, T.; York, D.; Pedersen, L. Particle mesh Ewald: An N log (N) method for Ewald sums in large systems. J. Chem. Phys. 1993, 98, 10089. [Google Scholar] [CrossRef]
- Stix, R.; Tan, X.F.; Bae, C.; Fernández-Mariño, A.I.; Swartz, K.J.; Faraldo-Gómez, J.D. Eukaryotic Kv channel Shaker inactivates through selectivity filter dilation rather than collapse. Sci. Adv. 2023, 9, eadj5539. [Google Scholar] [CrossRef]
- Guo, S.; Jackson, D.L.; Burcus, N.I.; Chen, Y.-J.; Xiao, S.; Heller, R. Gene electrotransfer enhanced by nanosecond pulsed electric fields. Mol. Ther. Methods Clin. Dev. 2014, 1, 14043. [Google Scholar] [CrossRef]
- Beebe, S.J.; Fox, P.M.; Rec, L.J.; Willis, E.L.; Schoenbach, K.H. Nanosecond, high-intensity pulsed electric fields induce apoptosis in human cells. Faseb J. 2003, 17, 1493–1495. [Google Scholar] [CrossRef]
- Marracino, P.; Caramazza, L.; Montagna, M.; Ghahri, R.; D’Abramo, M.; Liberti, M.; Apollonio, F. Electric-driven membrane poration: A rationale for water role in the kinetics of pore formation. Bioelectrochemistry 2022, 143, 107987. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Naikar, J.S.; Chan, S.S.Y.; Meivita, M.P.; Li, L.; Tan, Y.S.; Bajalovic, N.; Loke, D.K. Ultralong recovery time in nanosecond electroporation systems enabled by orientational-disordering processes. Nanoscale 2022, 14, 7934–7942. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Niu, Y.; Zhao, T.; Wang, X.; Wang, D.; Zhang, Y. Microscopic mechanistic study of the penetration distributions for plasma reactive oxygen and nitrogen species based on sialic acid targeting on the cell membrane surface. Free Radic. Biol. Med. 2024, 225, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38, 27-38. [Google Scholar] [CrossRef]
- Ho, M.C.; Levine, Z.A.; Vernier, P.T. Nanoscale, electric field-driven water bridges in vacuum gaps and lipid bilayers. J. Membr. Biol. 2013, 246, 793–801. [Google Scholar] [CrossRef]
- Chen, F.; Smith, P.E. Theory and computer simulation of solute effects on the surface tension of liquids. J. Phys. Chem. B 2008, 112, 8975–8984. [Google Scholar] [CrossRef]
- Maček Lebar, A.; Miklavčič, D.; Kotulska, M.; Kramar, P. Water Pores in Planar Lipid Bilayers at Fast and Slow Rise of Transmembrane Voltage. Membranes 2021, 11, 263. [Google Scholar] [CrossRef]
- Tang, J.; Ma, J.; Guo, L.; Wang, K.; Yang, Y.; Bo, W.; Yang, L.; Wang, Z.; Jiang, H.; Wu, Z.; et al. Interpretation of the molecular mechanism of the electroporation induced by symmetrical bipolar picosecond pulse trains. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183213. [Google Scholar] [CrossRef]
- Ye, P.; Huang, L.; Zhao, K. Bidirectional Modulation on Electroporation Induced by Membrane Tension Under the Electric Field. ACS Omega 2024, 9, 50458–50465. [Google Scholar] [CrossRef]
- Reddy, A.S.; Warshaviak, D.T.; Chachisvilis, M. Effect of membrane tension on the physical properties of DOPC lipid bilayer membrane. Biochim. Et Biophys. Acta Biomembr. 2012, 1818, 2271–2281. [Google Scholar] [CrossRef]
- Antila, H.S.; Wurl, A.; Ollila, O.H.S.; Miettinen, M.S.; Ferreira, T.M. Rotational decoupling between the hydrophilic and hydrophobic regions in lipid membranes. Biophys. J. 2022, 121, 68–78. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, P.; Huang, L.; Zhao, K. Fast-Rising Electric Pulses by Reducing Membrane Tension for Efficient Membrane Electroporation. Membranes 2025, 15, 151. https://doi.org/10.3390/membranes15050151
Ye P, Huang L, Zhao K. Fast-Rising Electric Pulses by Reducing Membrane Tension for Efficient Membrane Electroporation. Membranes. 2025; 15(5):151. https://doi.org/10.3390/membranes15050151
Chicago/Turabian StyleYe, Ping, Lulu Huang, and Kuiwen Zhao. 2025. "Fast-Rising Electric Pulses by Reducing Membrane Tension for Efficient Membrane Electroporation" Membranes 15, no. 5: 151. https://doi.org/10.3390/membranes15050151
APA StyleYe, P., Huang, L., & Zhao, K. (2025). Fast-Rising Electric Pulses by Reducing Membrane Tension for Efficient Membrane Electroporation. Membranes, 15(5), 151. https://doi.org/10.3390/membranes15050151