
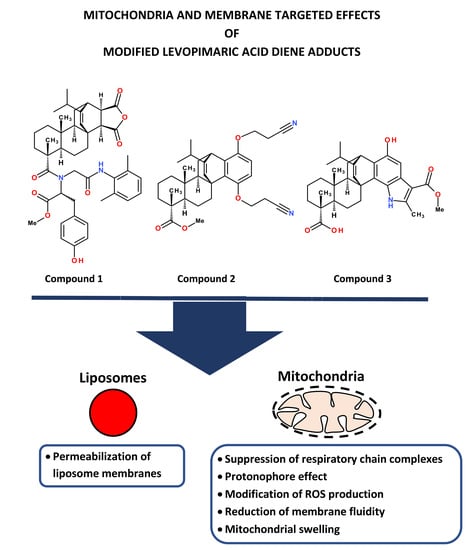
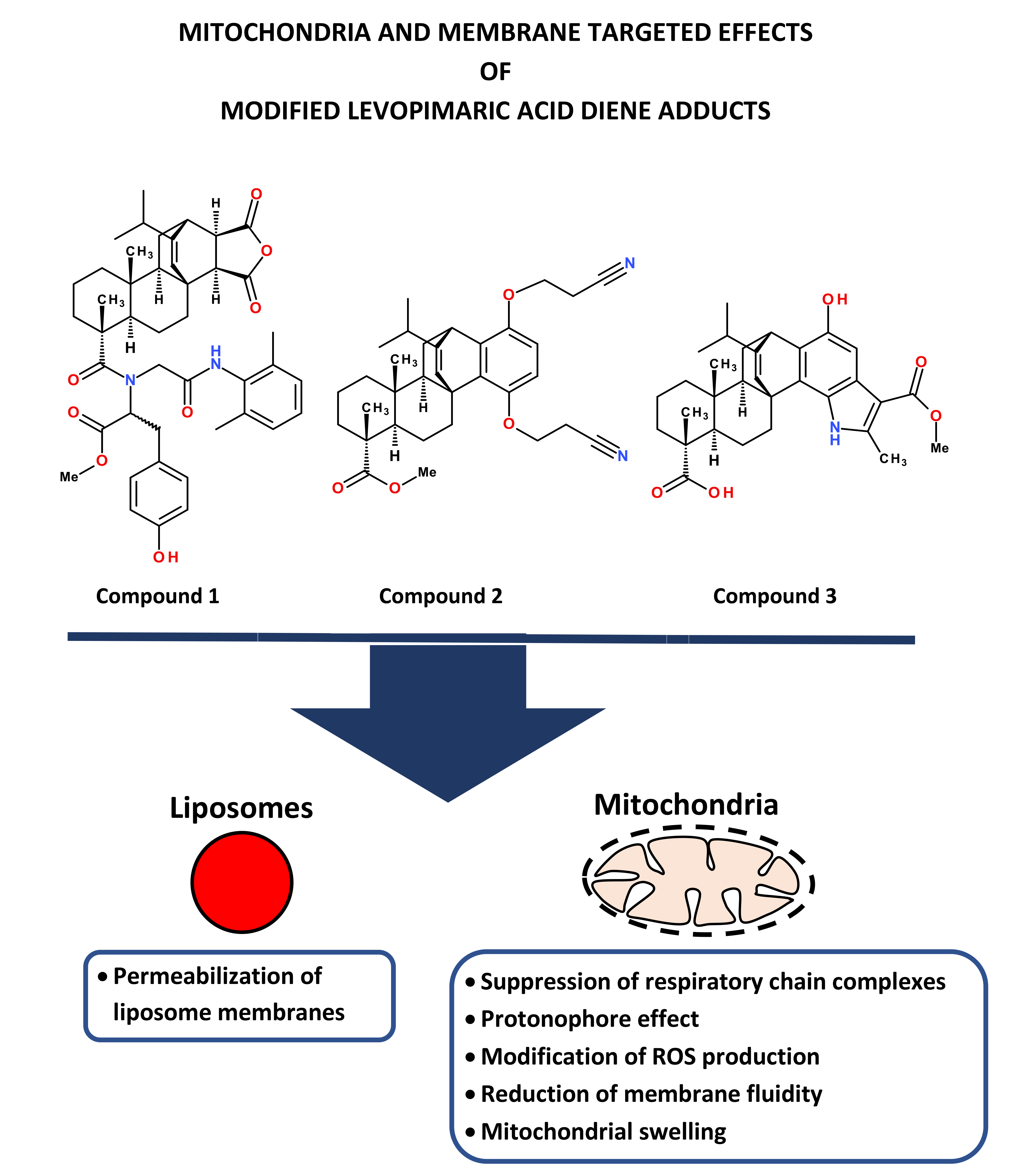
Effect of Modified Levopimaric Acid Diene Adducts on Mitochondrial and Liposome Membranes

 ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Reagents and Chemicals
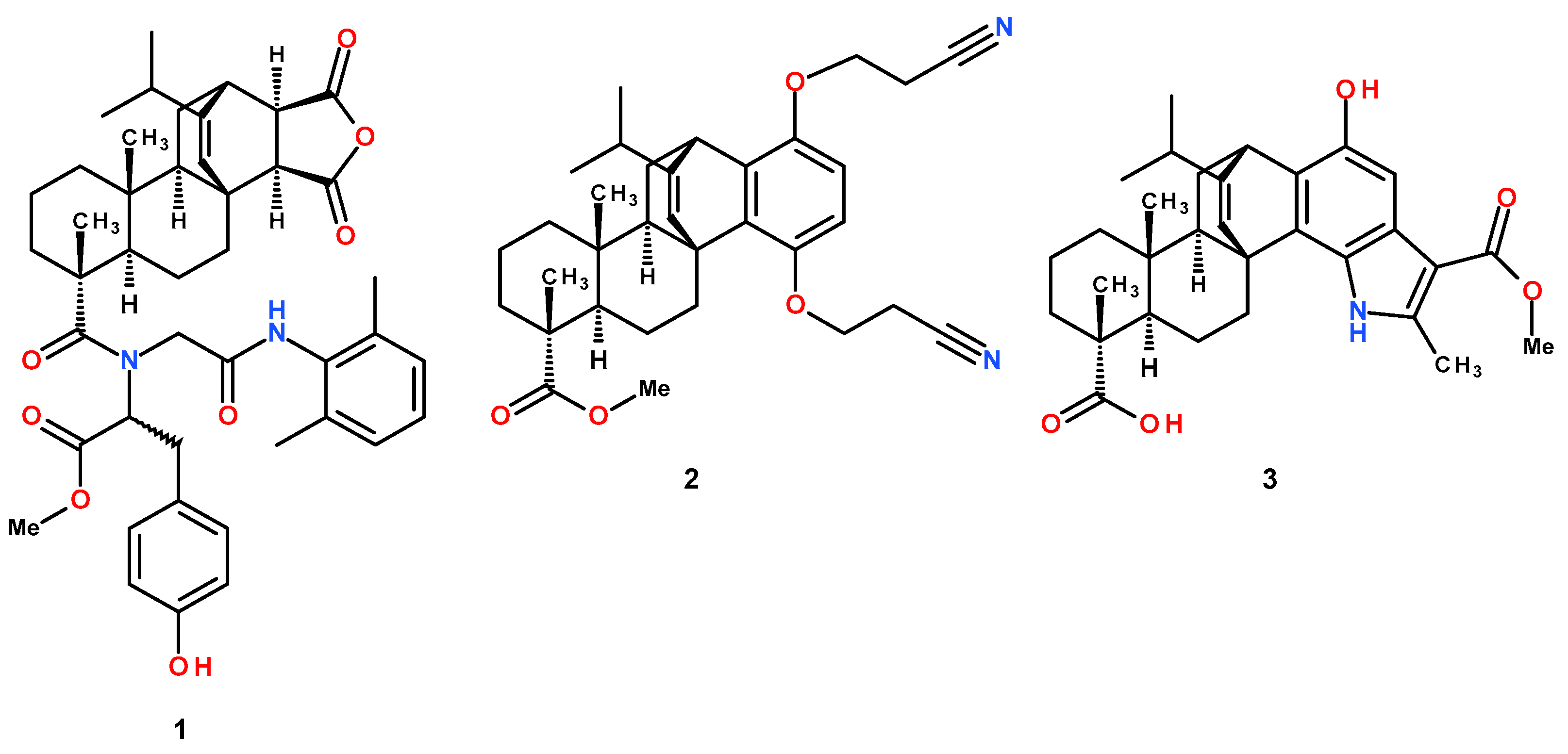
2.2. Synthesis of Modified Levopimaric Acid Diene Adducts
2.3. Isolation of Rat Liver Mitochondria
2.4. Determination of Mitochondrial Respiration and Oxidative Phosphorylation
2.5. Measuring Activity of Complexes of the Mitochondrial Electron Transport Chain (ETC)
2.6. Monitoring of Mitochondrial Membrane Potential
2.7. Production of H2O2 by Rat Liver Mitochondria
2.8. Monitoring Optical Density of Mitochondrial Suspension
2.9. Evaluation of Mitochondrial Membrane Fluidity by Generalized Polarization (GP) of Laurdan
2.10. Construction of Liposomes
2.11. Statistical Analysis
3. Results
3.1. Modified Levopimaric Acid Diene Adducts Reduce the Efficiency of Oxidative Phosphorylation in Rat Liver Mitochondria
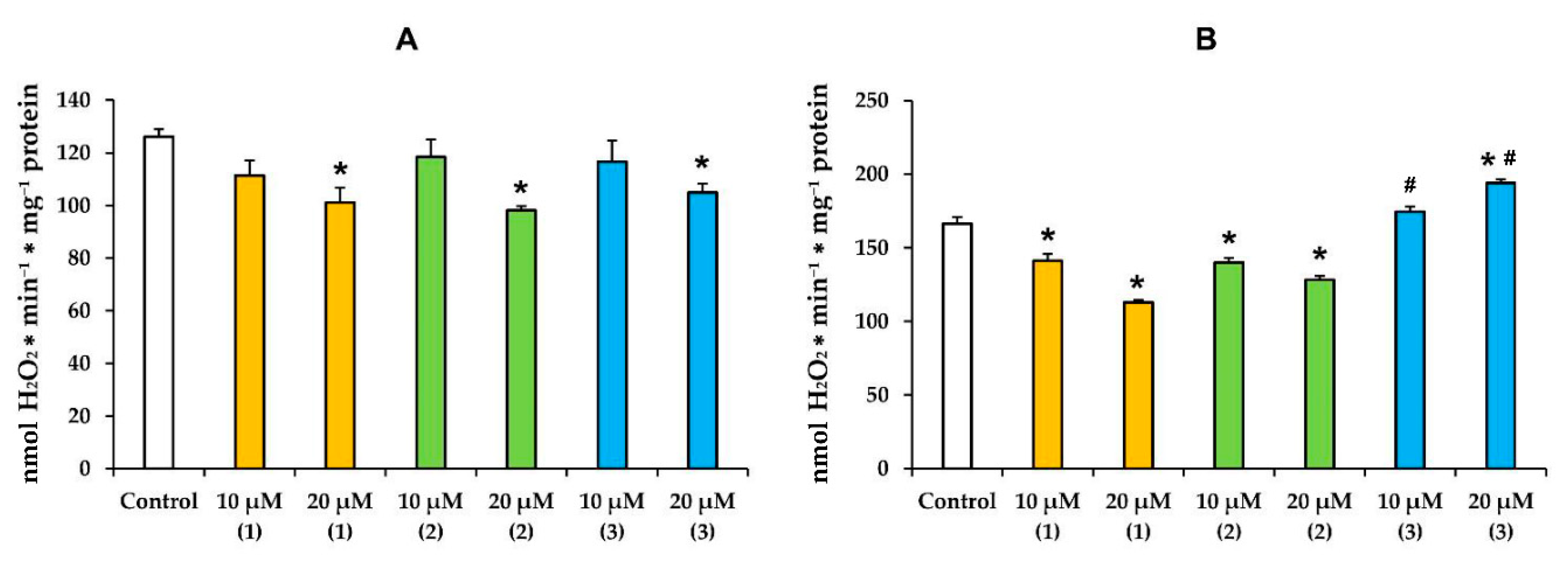
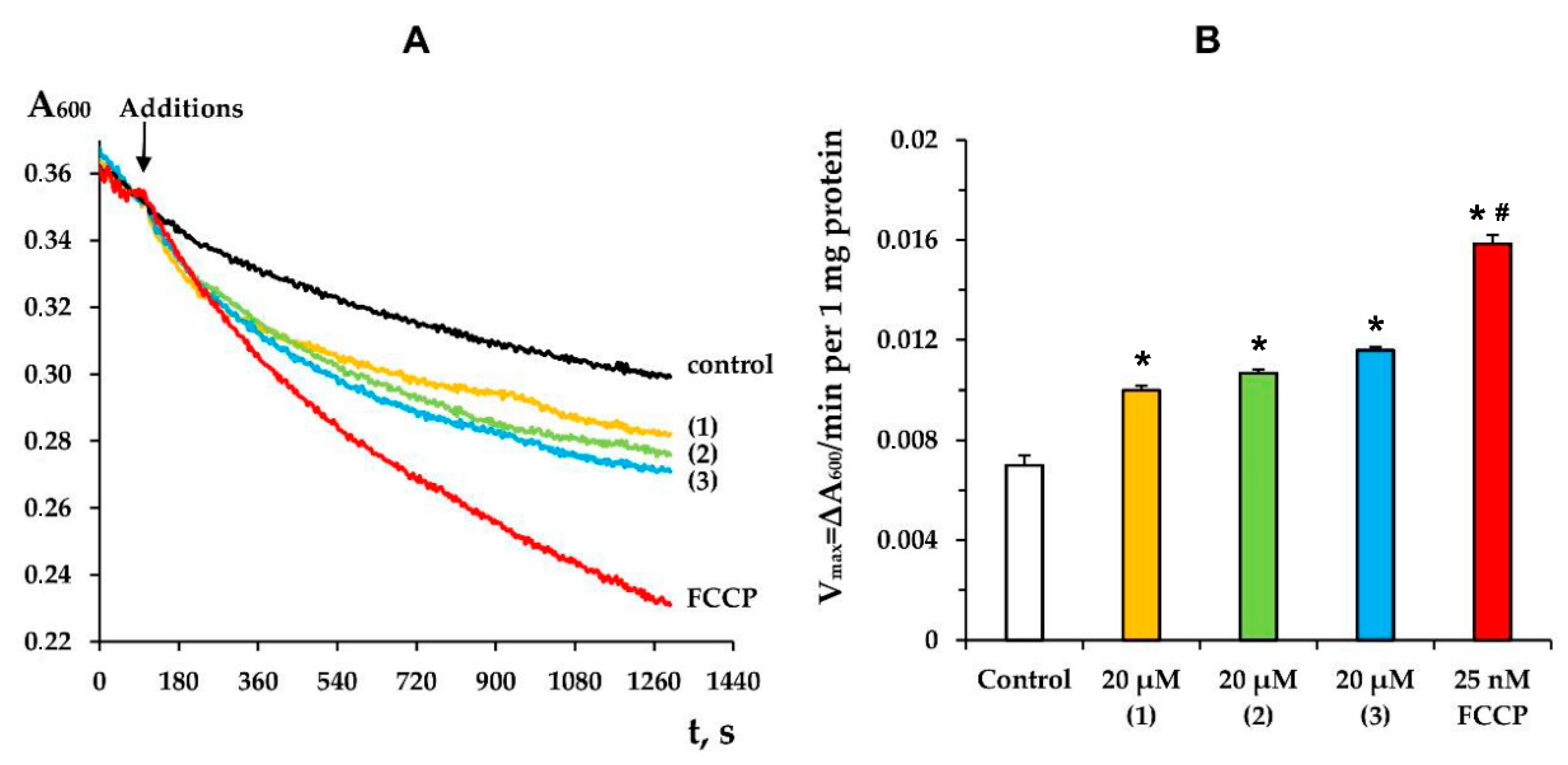
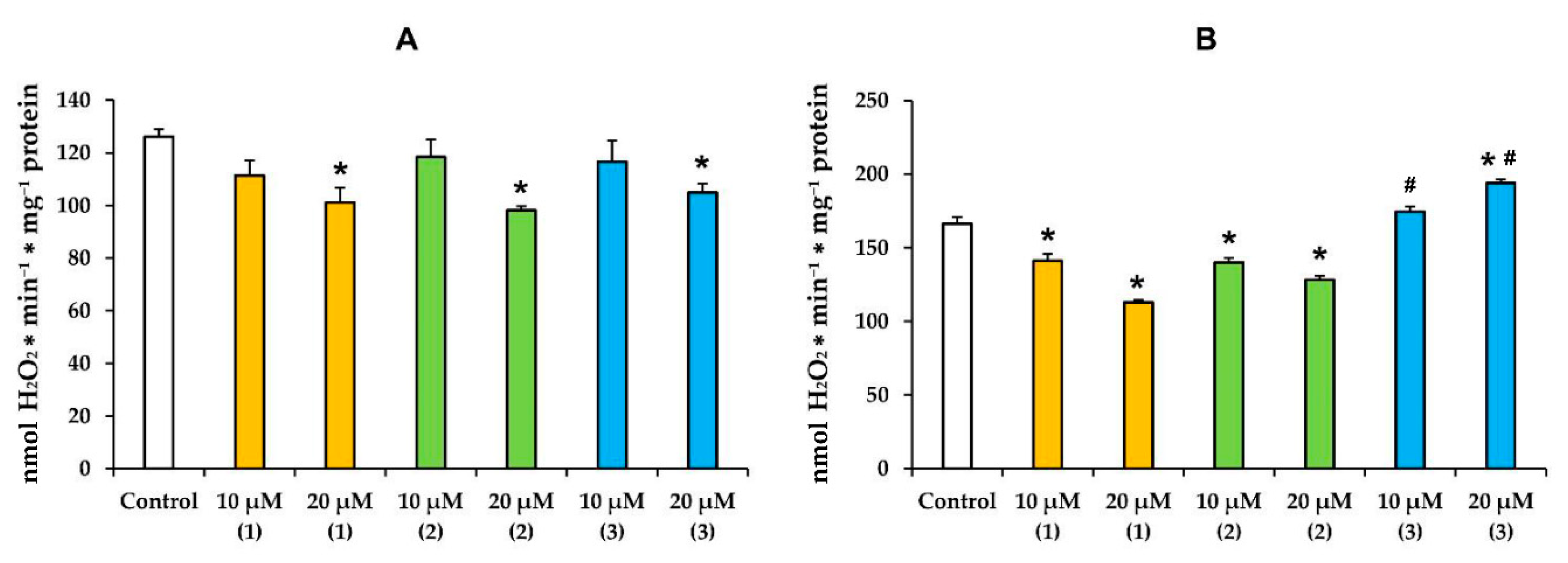
3.2. Modified Levopimaric Acid Diene Adducts Modulate Hydrogen Peroxide Production by Liver Mitochondria
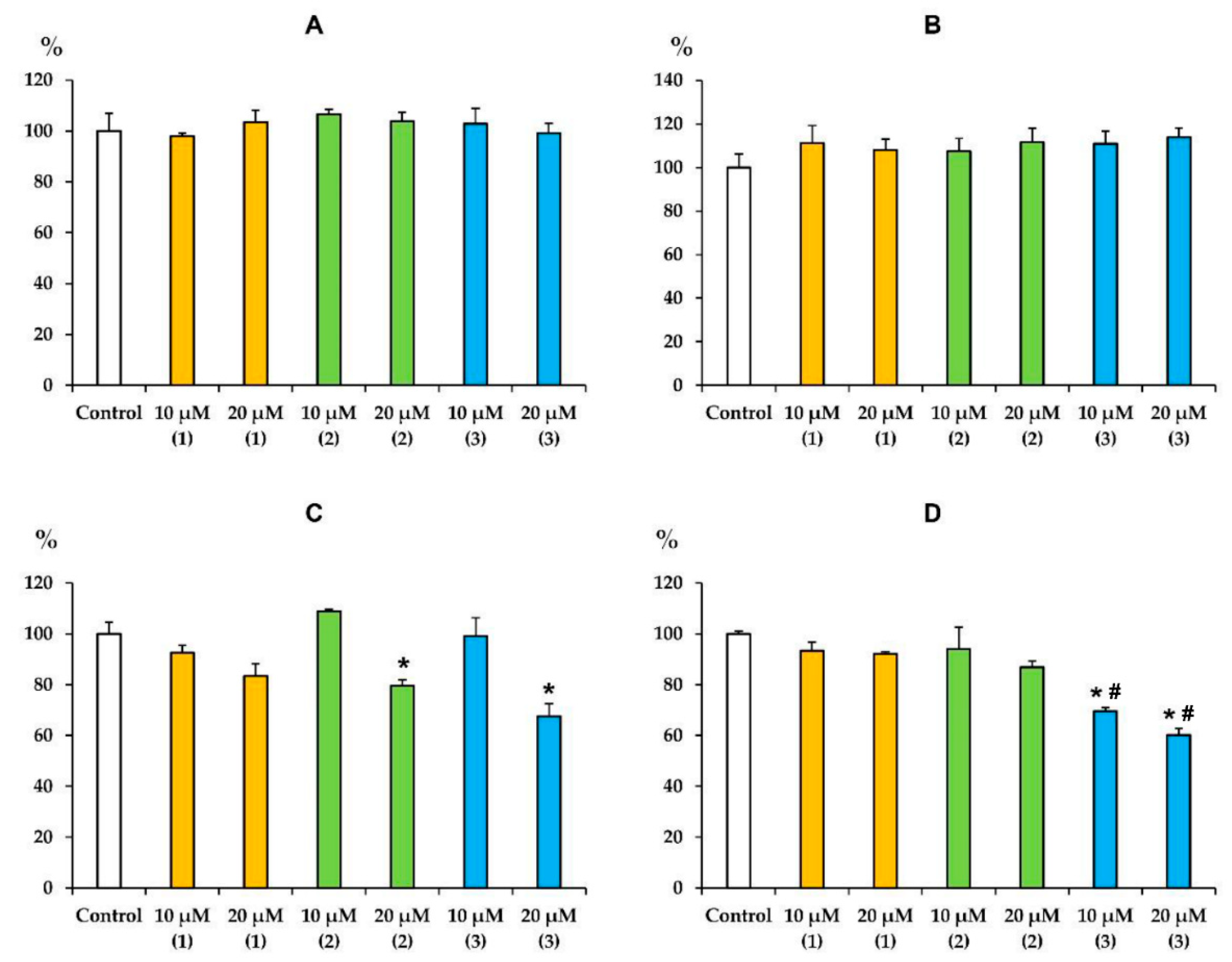
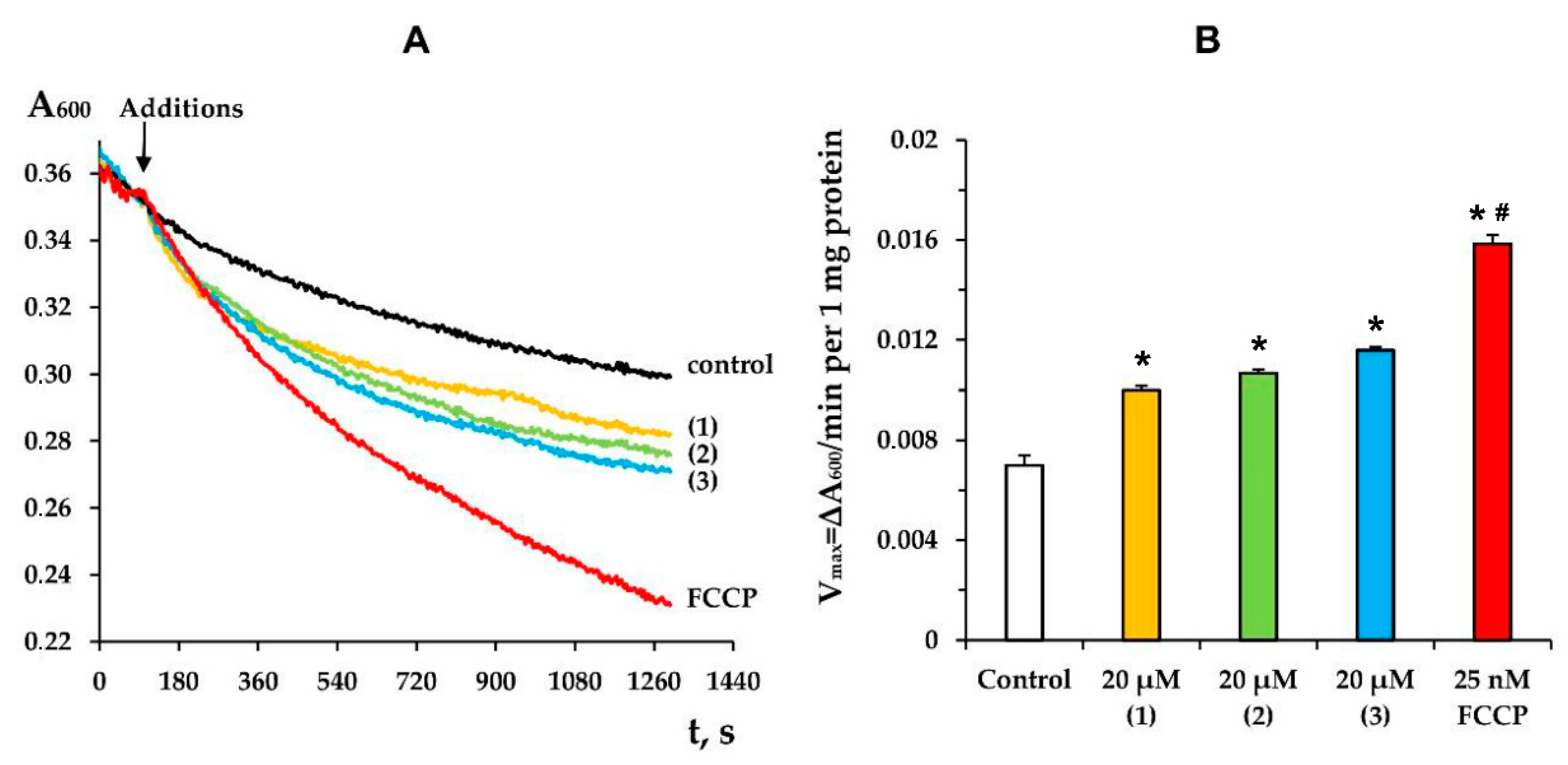
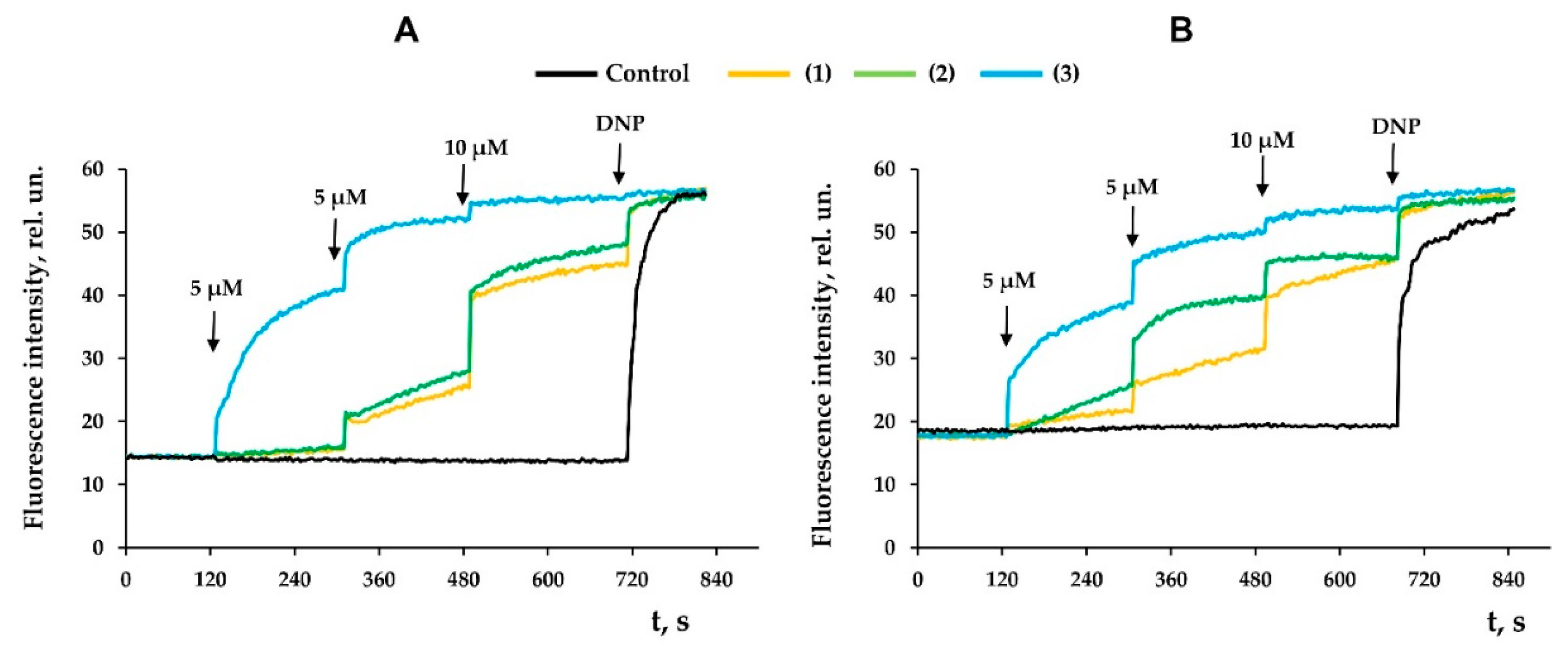
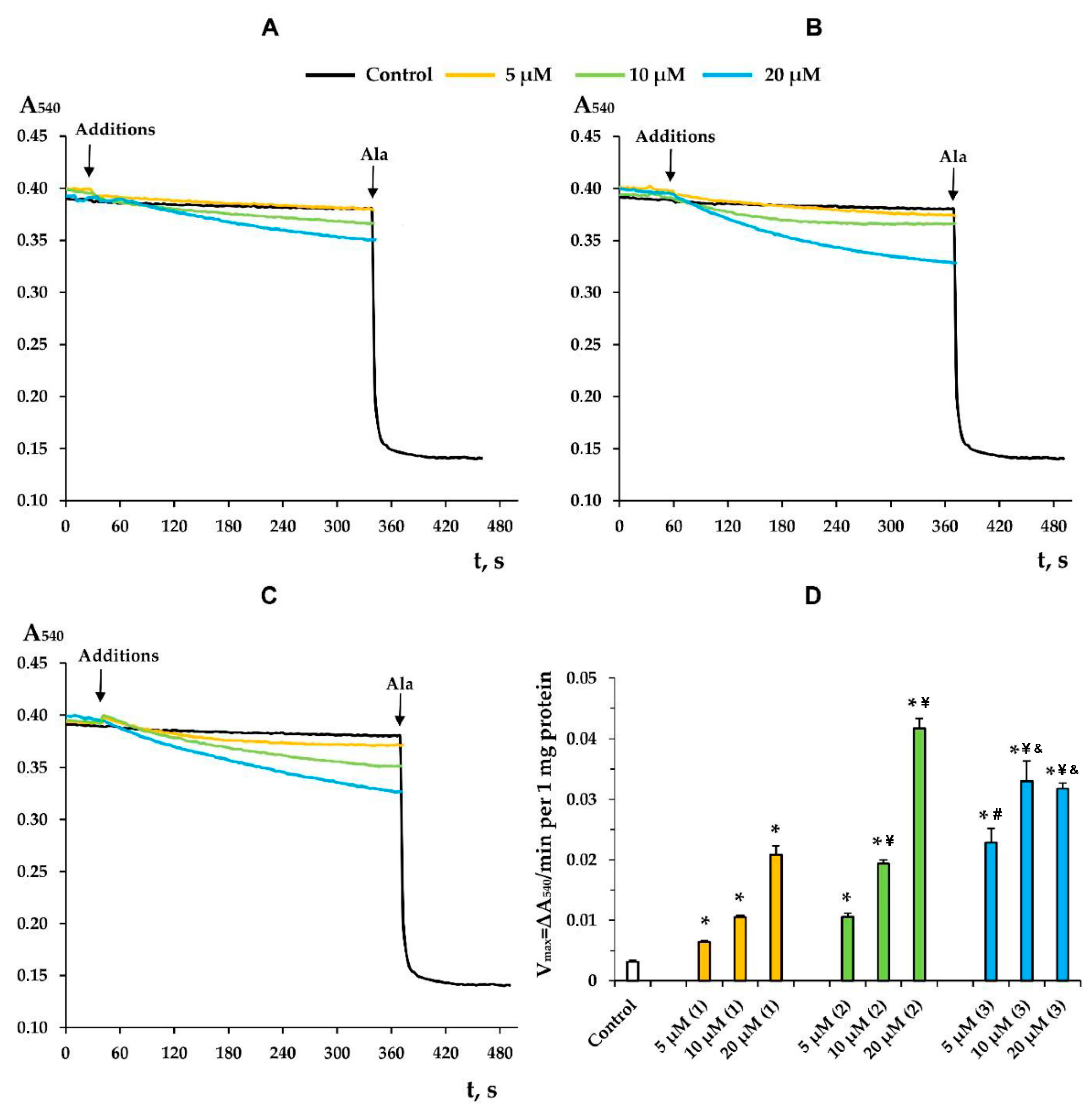
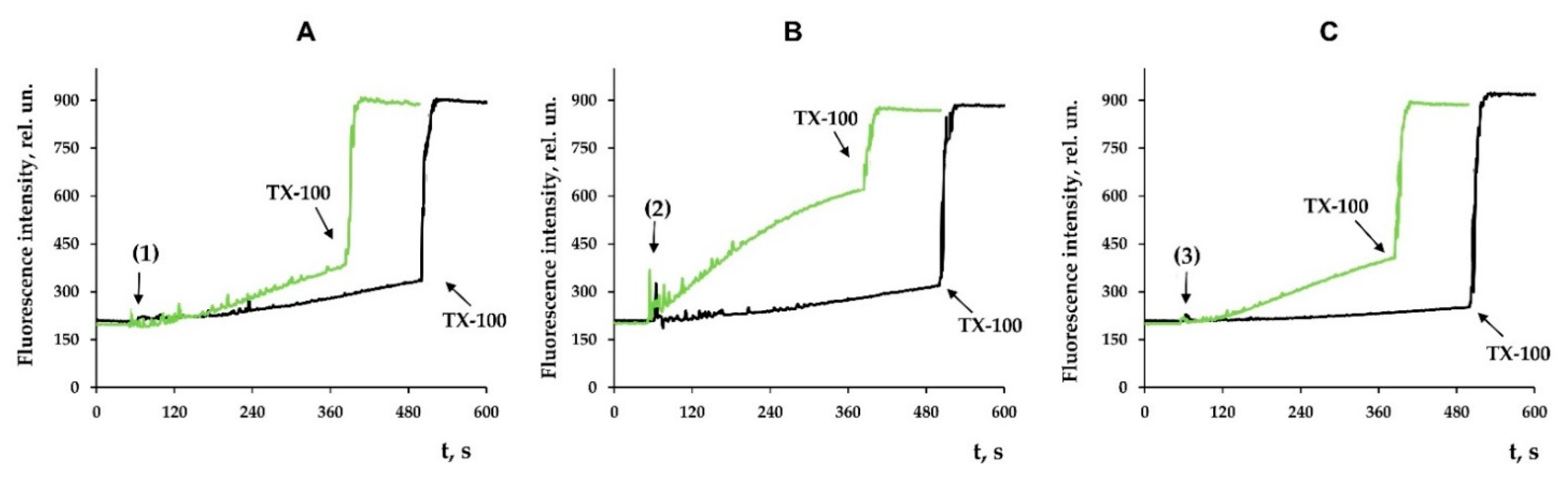
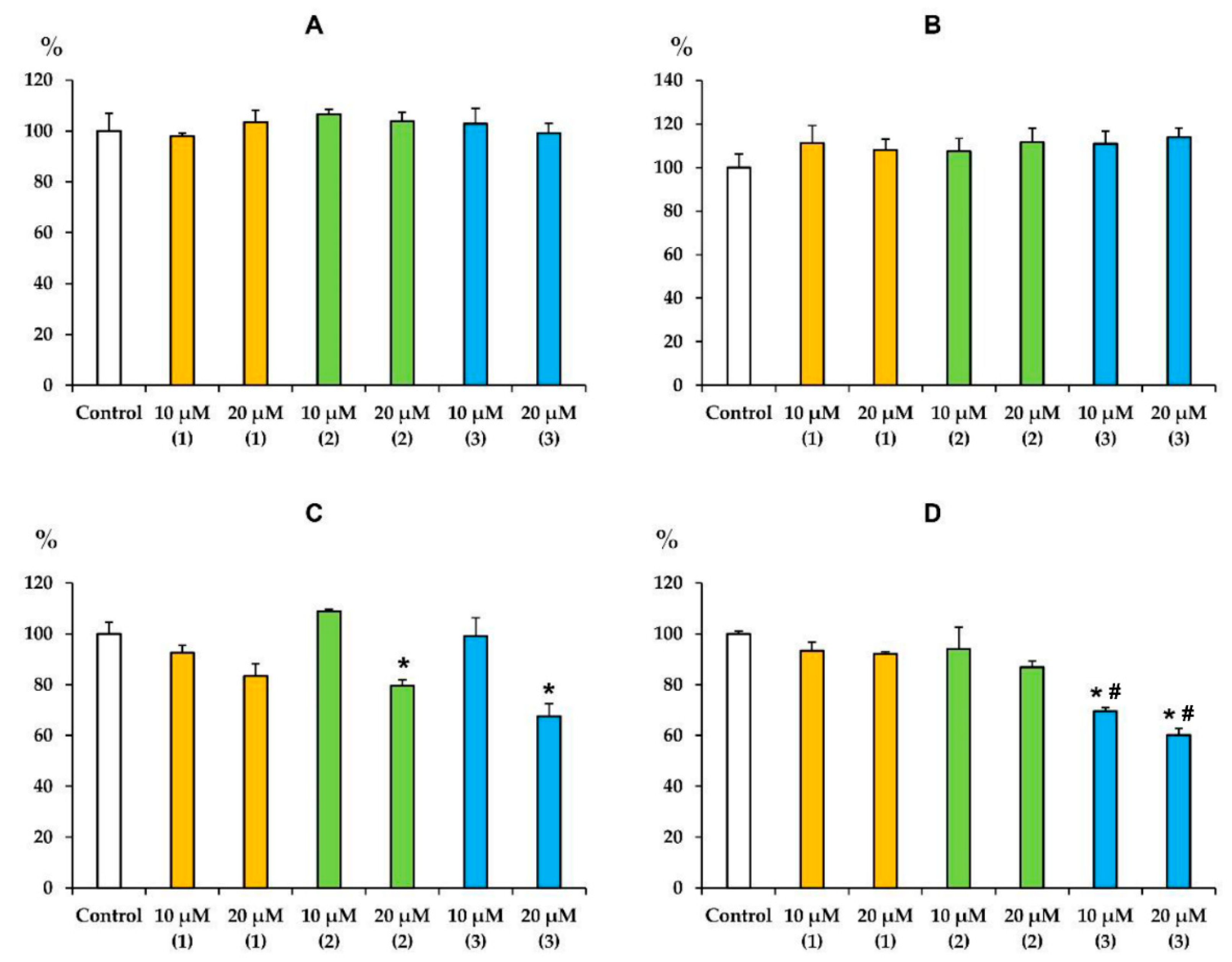
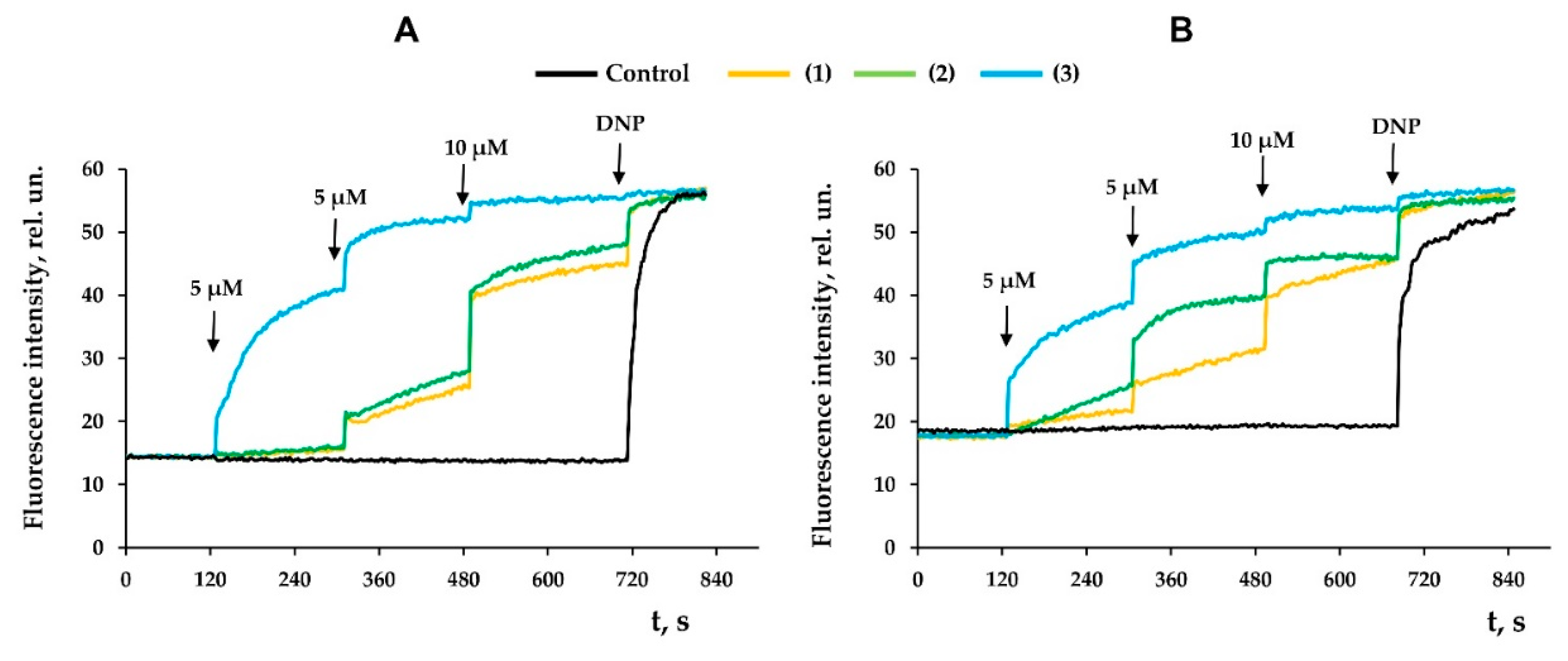
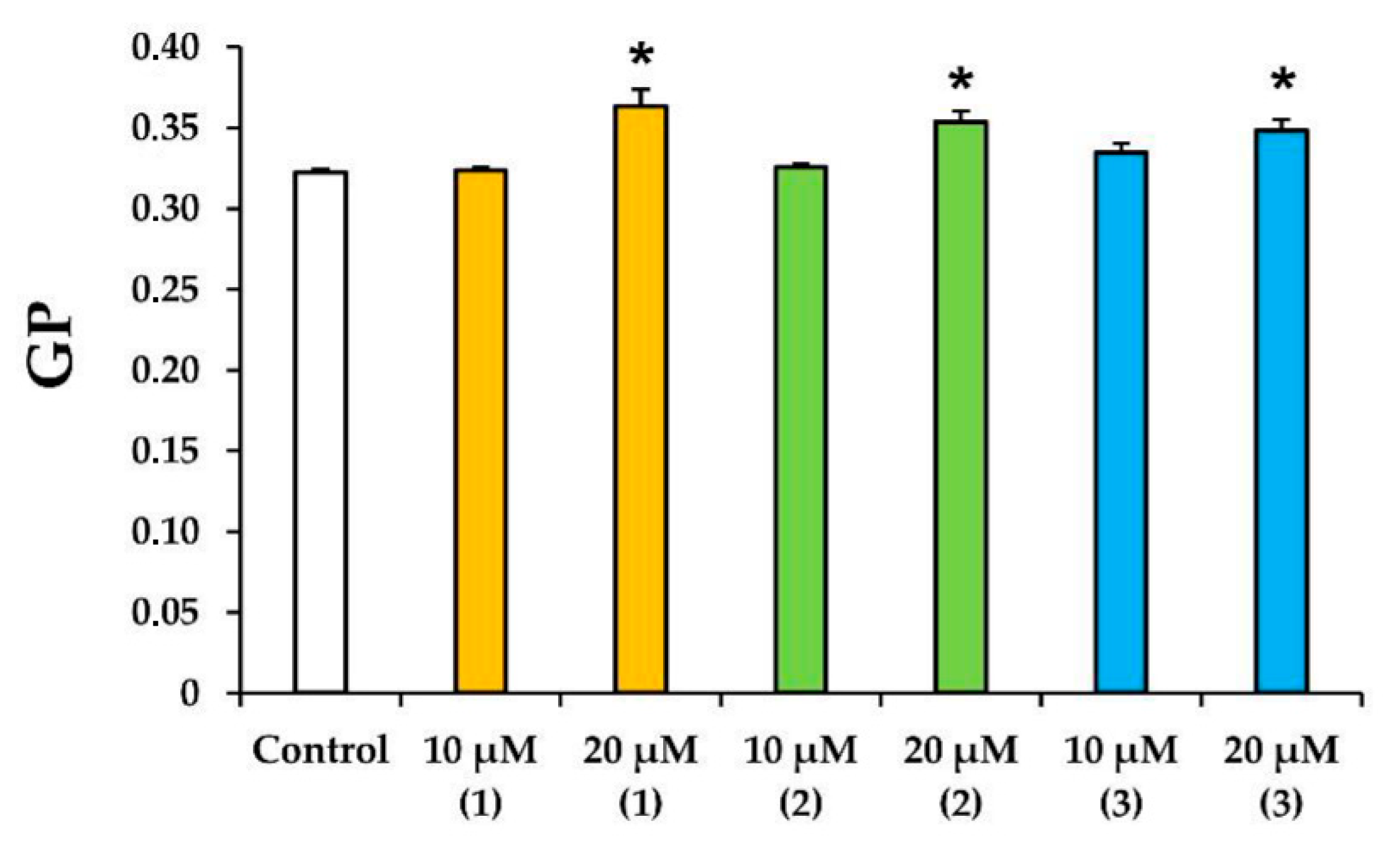
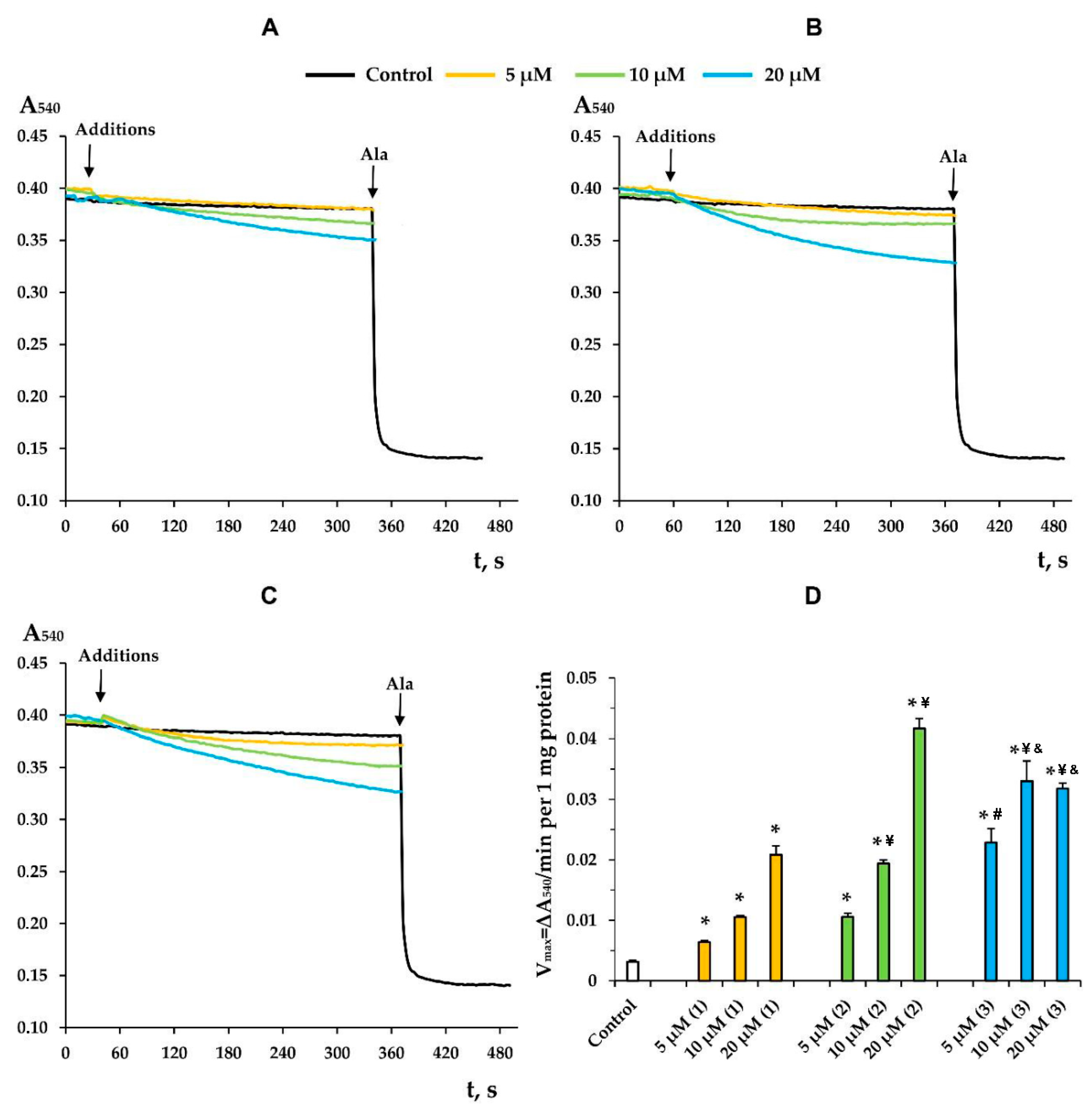
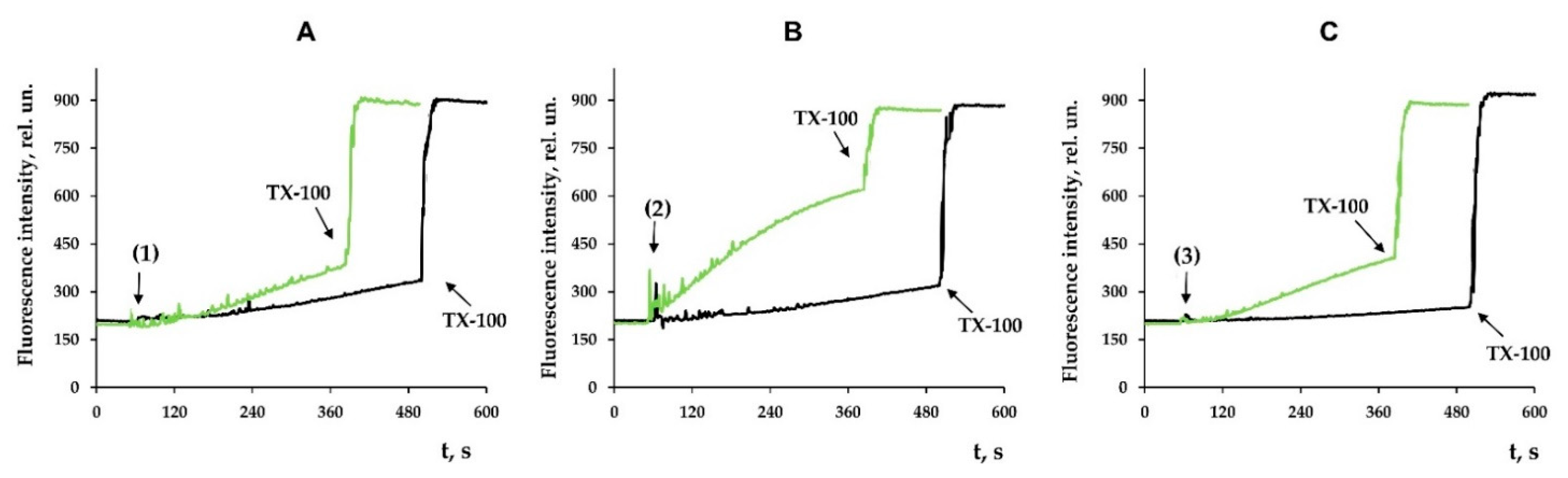
3.3. Modified Levopimaric Acid Diene Adducts Decrease the Overall Fluidity of the Mitochondrial Membranes and Induce Permeabilization of Mitochondrial Membranes as well as Lecithin Liposome Membranes
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hou, W.; Zhang, G.; Luo, Z.; Li, D.; Ruan, H.; Ruan, B.H.; Su, L.; Xu, H. Identification of a diverse synthetic abietane diterpenoid library and insight into the structure-activity relationships for antibacterial activity. Bioorg. Med. Chem. Lett. 2017, 27, 5382–5386. [Google Scholar] [PubMed]
- Zhang, G.-J.; Li, Y.-H.; Jiang, J.-D.; Yu, S.-S.; Qu, J.; Maa, S.-G.; Liu, Y.-B.; Yu, D.-Q. Anti-coxsackie virus B diterpenes from the rootsof Illicium jiadifengpi. Tetrahedron 2013, 69, 1017–1023. [Google Scholar] [CrossRef]
- Pirttimaa, M.; Nasereddin, A.; Kopelyanskiy, D.; Kaiser, M.; Yli-Kauhaluoma, J.; Oksman-Caldentey, K.-M.; Brun, R.; Jaffe, C.h.L.; Moreira, V.M.; Alakurtti, S. Abietane-type diterpenoid amides with highly potent and selective activity against Leishmania donovani and Trypanosoma cruzi. J. Nat. Prod. 2016, 79, 362–368. [Google Scholar] [PubMed]
- Kazakova, O.B.; Smirnova, I.E.; Tkhi Tkhu Do, H.; Nguen, T.T.; Apryshko, G.N.; Zhukova, O.S.; Medvedeva, N.I.; Nazyrov, T.I.; Tret’yakova, E.V.; Chudov, I.V.; et al. Synthesis, structure, and pharmacological activity of (7R,8S)-Epoxy-(13R,17R)-trioxolane abietic acid. Russ. J. Bioorg. Chem. 2013, 39, 202–210. [Google Scholar]
- Mohamed, M.A. Anti-inflammatory activity of heterocyclic systems using abietic acid as starting material. Monatsh. Chem. 2008, 139, 697–705. [Google Scholar]
- Justino, G.A.C.; Correia, C.F.; Mira, L.; Dos Santos, R.M.B.; Simones, J.A.M.; Silva, A.M.; Santos, C.L.; Gigante, B.R. Antioxidant activity of a catechol derived from abietic acid. J. Agric. Food Chem. 2006, 54, 342–348. [Google Scholar]
- Xu, H.; Liu, L.; Fan, X.; Zhang, G.; Li, Y.; Jiang, B. Identification of a diverse synthetic abietane diterpenoid library for anticancer activity. Bioorg. Med. Chem. Lett. 2017, 27, 505–510. [Google Scholar] [CrossRef]
- Akaberi, M.; Mehri, S.; Iranshahi, M. Multiple pro-apoptotic targets of abietane diterpenoids from Salvia species. Fitoterapia 2015, 100, 118–132. [Google Scholar]
- Jung, T.W.; Jeong, J.C.; Park, S.Y.; Cho, W.; Oh, H.; Lee, H.J.; Hacimuftuoglu, A.; Abd El-Aty, A.M.; Bang, J.S.; Jeong, J.H. Abietic acid alleviates endoplasmic reticulum stress and lipid accumulation in human primary hepatocytes through the AMPK/ORP150 signaling. Biochem. Biophys. Res. Commun. 2022, 608, 142–148. [Google Scholar]
- Faustino, C.; Neto, Í.; Fonte, P.; Macedo, A. Cytotoxicity and Chemotherapeutic Potential of Natural Rosin Abietane Diterpenoids and their Synthetic Derivatives. Curr. Pharm. Des. 2018, 24, 4362–4375. [Google Scholar] [CrossRef]
- Salminen, A.; Lehtonen, M.; Suuronen, T.; Kaarniranta, K.; Huuskonen, J. Terpenoids: Natural inhibitors of NF-κB signaling with anti-inflammatory and anticancer potential. Cell. Mol. Life Sci. 2008, 65, 2979–2999. [Google Scholar] [PubMed]
- De Oliveira, M.R.; da Costa Ferreira, G.; Peres, A.; Bosco, S.M.D. Carnosic Acid Suppresses the H2O2-Induced Mitochondria-Related Bioenergetics Disturbances and Redox Impairment in SH-SY5Y Cells: Role for Nrf2. Mol. Neurobiol. 2018, 55, 968–979. [Google Scholar] [PubMed]
- Sakamoto, K.; Ohya, S.; Muraki, K.; Imaizumi, Y. A Novel Opener of Large-Conductance Ca2+-Activated K+ (BK) Channel Reduces Ischemic Injury in Rat Cardiac Myocytes by Activating Mitochondrial KCa Channel. J. Pharmacol Sci. 2008, 108, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Qu, H.-E.; Huang, X.-C.; Pan, Y.-M.; Liang, D.; Chen, Z.-F.; Wang, H.-S.; Zhang, Y. Synthesis and Biological Evaluation of Novel Dehydroabietic Acid Derivatives Conjugated with Acyl-Thiourea Peptide Moiety as Antitumor Agents. Int. J. Mol. Sci. 2015, 16, 14571–14593. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; He, F.; Liu, H.; Zhu, Y.; Tian, W.; Gao, P.; He, H.; Yue, W.; Lei, X.; Ni, B.; et al. Natural diterpenoid compound elevates expression of Bim protein, which interacts with antiapoptotic protein Bcl-2, converting it to proapoptotic Bax-like molecule. J. Biol. Chem. 2012, 287, 1054–1065. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Ding, J. In vitro anticancer effects of levopimaric acid in cisplatinresistant human lung carcinoma are mediated via autophagy, ROS-mediated mitochondrial dysfunction, cell apoptosis and modulation of ERK/MAPK/JNK signalling pathway. J. BUON. 2020, 25, 248–254. [Google Scholar]
- Tretyakova, E.V.; Smirnova, I.E.; Kazakova, O.B.; Tolstikov, G.A.; Yavorskaya, N.P.; Golubeva, I.S.; Pugacheva, R.B.; Apryshko, G.N.; Poroikov, V.V. Synthesis and anticancer activity of quinopimaric and maleopimaric acids’ derivatives. Bioorganic Med. Chem. 2014, 22, 6481–6489. [Google Scholar]
- Tretyakova, E.V.; Salimova, E.V.; Parfenova, L.V.; Yunusbaeva, M.M.; Dzhemileva, L.U.; D’yakonov, V.V.; Dzhemilev, U.M. Synthesis of New Dihydroquinopimaric Acid Analogs with Nitrile Groups as Apoptosis-Inducing Anticancer Agents. Anticancer Agents Med. Chem. 2019, 19, 1172–1183. [Google Scholar] [CrossRef]
- Tretyakova, E.V.; Ma, X.; Kazakova, O.B.; Shtro, A.A.; Petukhova, G.D.; Smirnova, A.A.; Xu, H.; Xiao, S. Abietic, Maleopimaric and Quinopimaric Dipeptide Ugi-4CR Derivatives and Their Potency against Influenza A and SARS-CoV-2. Nat. Prod. Res. 2022, 9, 1–7. [Google Scholar] [CrossRef]
- Tretyakova, E.V.; Yarmukhametova, L.R.; Salimova, E.V.; Kukovinets, O.S.; Parfenova, L.V. The Nenitzescu reaction in the synthesis of new abietane diterpene indoles. Chem. Heterocycl. Compd. 2020, 56, 1366–1369. [Google Scholar] [CrossRef]
- Belosludtsev, K.N.; Belosludtseva, N.V.; Tenkov, K.S.; Penkov, N.V.; Agafonov, A.V.; Pavlik, L.L.; Yashin, V.A.; Samartsev, V.N.; Dubinin, M.V. Study of the mechanism of permeabilization of lecithin liposomes and rat liver mitochondria by the antimicrobial drug triclosan. Biochim. Biophys. Acta Biomembr. 2018, 1860, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Belosludtsev, K.N.; Belosludtseva, N.V.; Kosareva, E.A.; Talanov, E.Y.; Gudkov, S.V.; Dubinin, M.V. Itaconic acid impairs the mitochondrial function by the inhibition of complexes II and IV and induction of the permeability transition pore opening in rat liver mitochondria. Biochimie 2020, 176, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Spinazzi, M.; Casarin, A.; Pertegato, V.; Salviati, L.; Angelini, C. Assessment of mitochondrial respiratory chain enzymatic activities on tissues and cultured cells. Nat. Protoc. 2012, 7, 1235–1246. [Google Scholar] [CrossRef] [PubMed]
- Dubinin, M.V.; Sharapov, V.A.; Ilzorkina, A.I.; Efimov, S.V.; Klochkov, V.V.; Gudkov, S.V.; Belosludtsev, K.N. Comparison of structural properties of cyclosporin A and its analogue alisporivir and their effects on mitochondrial bioenergetics and membrane behavior. Biochim. Biophys. Acta Biomembr. 2022, 1864, 183972. [Google Scholar] [CrossRef] [PubMed]
- Dubinin, M.V.; Semenova, A.A.; Ilzorkina, A.I.; Penkov, N.V.; Nedopekina, D.A.; Sharapov, V.A.; Khoroshavina, E.I.; Davletshin, E.V.; Belosludtseva, N.V.; Spivak, A.Y.; et al. Mitochondria-targeted prooxidant effects of betulinic acid conjugated with delocalized lipophilic cation F16. Free Radic. Biol. Med. 2021, 168, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Parasassi, T.; Krasnowska, E.K.; Bagatolli, L.; Gratton, E. Laurdan and Prodan as polarity-sensitive fluorescent membrane probes. J. Fluoresc. 1998, 8, 365–373. [Google Scholar] [CrossRef]
- Parasassi, T.; Stasio, G.; d’Ubaldo, A.; Gratton, E. Phase fluctuation in phospholipid membranes revealed by Laurdan Fluorescence. Biophys. J. 1990, 57, 1179–1186. [Google Scholar] [CrossRef] [Green Version]
- Dubinin, M.V.; Semenova, A.A.; Ilzorkina, A.I.; Mikheeva, I.B.; Yashin, V.A.; Penkov, N.V.; Vydrina, V.A.; Ishmuratov, G.Y.; Sharapov, V.A.; Khoroshavina, E.I.; et al. Effect of betulin and betulonic acid on isolated rat liver mitochondria and liposomes. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183383. [Google Scholar] [CrossRef]
- Dubinin, M.V.; Semenova, A.A.; Nedopekina, D.A.; Davletshin, E.V.; Spivak, A.Y.; Belosludtsev, K.N. Effect of F16-Betulin Conjugate on Mitochondrial Membranes and Its Role in Cell Death Initiation. Membranes 2021, 11, 352. [Google Scholar] [CrossRef]
- Dubinin, M.V.; Semenova, A.A.; Ilzorkina, A.I.; Markelova, N.Y.; Penkov, N.V.; Shakurova, E.R.; Belosludtsev, K.N.; Parfenova, L.V. New quaternized pyridinium derivatives of betulin: Synthesis and evaluation of membranotropic properties on liposomes, pro- and eukaryotic cells, and isolated mitochondria. Chem Biol/Interact. 2021, 349, 109678. [Google Scholar] [CrossRef]
- Brierley, G.P.; Jurkowitz, M.; Scott, K.M.; Merola, A.J. Ion transport by heart mitochondria. XX. Factors affecting passive osmotic swelling of isolated mitochondria. J. Biol. Chem. 1970, 245, 5404–5411. [Google Scholar] [CrossRef]
- Kruszniewska-Rajs, C.; Strzałka-Mrozik, B.; Kimsa-Dudek, M.; Synowiec-Wojtarowicz, A.; Chrobak, E.; Bębenek, E.; Boryczka, S.; Głuszek, S.; Gola, J.M. The Influence of Betulin and Its Derivatives EB5 and ECH147 on the Antioxidant Status of Human Renal Proximal Tubule Epithelial Cells. Int. J. Mol. Sci. 2022, 23, 2524. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wu, J.; Han, Y.; Zhang, J.; Lin, Y.; Wang, H.; Wang, J.; Liu, J.; Bu, M. Design and Synthesis of Novel Betulin Derivatives Containing Thio-/Semicarbazone Moieties as Apoptotic Inducers through Mitochindria-Related Pathways. Molecules 2021, 26, 6356. [Google Scholar] [CrossRef]
- Kersten, P.J.; Kopper, B.J.; Raffa, K.F.; Illman, B.L. Rapid Analysis of Abietanes in Conifers. J. Chem. Ecol. 2003, 32, 2679–2685. [Google Scholar] [CrossRef] [PubMed]
- San Feliciano, A.; Gordaliza, M.; Salinero, M.A.; Miguel Del Corral, J.M. Abietane acids: Sources, biological activities, and therapeutic uses. Planta Medica 1993, 59, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Tretyakova, E.V.; Smirnova, I.E.; Salimova, E.V.; Odinokov, V.N. Synthesis and antiviral activity of maleopimaric and quinopimaric acids’ derivatives. Bioorg. Med. Chem. 2015, 23, 6543–6550. [Google Scholar] [CrossRef] [PubMed]
- Qi, Z.-W.; Jiang, L.-H.; Wang, Y.-M.; Zhang, S. Research advance in isolation and application of levopimaric acid. Chem. Ind. For. Prod. 2016, 36, 127–132. [Google Scholar]
- Malikova, R.; Sakhautdinov, I. Maleated rosin-derived advanced materials: Preparation, properties and application. J. Iran Chem. Soc. 2022, 19, 3229–3248. [Google Scholar] [CrossRef]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef]
- Paradies, G.; Paradies, V.; Ruggiero, F.M.; Petrosillo, G. Role of Cardiolipin in Mitochondrial Function and Dynamics in Health and Disease: Molecular and Pharmacological Aspects. Cells 2019, 8, 728. [Google Scholar] [CrossRef]
- Wang, Y.; Du, Y.; Huang, N. A survey of the role of nitrile groups in protein-ligand interactions. Future Med. Chem. 2018, 10, 2713–2728. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Additions | State 2 | State 3 | State 4 | State 3UDNP | RC | ADP/O |
|---|---|---|---|---|---|---|
| nmol O2 * min−1 * mg−1 Protein | rel. un. | |||||
| Control | 2.50 ± 0.22 | 21.46 ± 0.55 | 3.18 ± 0.04 | 22.59 ± 1.75 | 6.75 ± 0.19 | 2.93 ± 0.03 |
| Compound 1 | ||||||
| 10 μM | 2.42 ± 0.22 | 20.78 ± 0.58 | 2.99 ± 0.14 | 23.13 ± 0.97 | 6.97 ± 0.16 | 2.91 ± 0.05 |
| 20 μM | 2.58 ± 0.21 | 19.80 ± 0.48 | 3.13 ± 0.10 | 20.58 ± 0.26 | 6.28 ± 0.03 | 2.90 ± 0.03 |
| Compound 2 | ||||||
| 10 μM | 2.59 ± 0.18 | 20.14 ± 0.84 | 3.11 ± 0.20 | 24.02 ± 1.09 | 6.49 ± 0.15 | 2.94 ± 0.06 |
| 20 μM | 3.25 ± 0.31 | 18.15 ± 0.17 * | 3.77 ± 0.09 * | 24.05 ± 0.35 | 4.82 ± 0.04 * | 2.98 ± 0.02 |
| Compound 3 | ||||||
| 10 μM | 3.16 ± 0.23 | 19.28 ± 0.13 | 3.75 ± 0.11 * | 18.83 ± 0.23 | 5.15 ± 0.12 * | 2.68 ± 0.03 * |
| 20 μM | 3.56 ± 0.12 * | 16.19 ± 0.47 * | 4.61 ± 0.08 *# | 15.75 ± 0.68 * | 3.51 ± 0.04 *# | 2.60 ± 0.01 * |
| Additions | State 2 | State 3 | State 4 | State 3UDNP | RC | ADP/O |
|---|---|---|---|---|---|---|
| nmol O2 * min−1 * mg−1 protein | rel. un. | |||||
| Control | 7.78 ± 0.18 | 47.77 ± 0.25 | 8.65 ± 0.19 | 57.48 ± 0.52 | 5.53 ± 0.09 | 1.79 ± 0.05 |
| Compound 1 | ||||||
| 10 μM | 7.01 ± 0.23 | 43.45 ± 0.35 * | 7.83 ± 0.13 | 53.51 ± 1.00 | 5.55 ± 0.05 | 1.71 ± 0.05 |
| 20 μM | 8.22 ± 0.19 | 38.54 ± 0.16 * | 8.84 ± 0.11 | 53.39 ± 0.87 | 4.36 ± 0.04 * | 1.75 ± 0.07 |
| Compound 2 | ||||||
| 10 μM | 7.21 ± 0.50 | 42.32 ± 0.23 * | 8.29 ± 0.15 | 52.51 ± 0.51 | 5.11 ± 0.12 * | 1.83 ± 0.11 |
| 20 μM | 7.94 ± 0.10 | 33.92 ± 0.98 *# | 10.75 ± 0.49 * | 46.11 ± 2.23 * | 3.16 ± 0.06 *# | 1.89 ± 0.06 |
| Compound 3 | ||||||
| 10 μM | 7.48 ± 0.32 | 41.83 ± 1.32 * | 8.64 ± 0.53 | 52.29 ± 2.11 | 4.85 ± 0.15 * | 1.91 ± 0.08 |
| 20 μM | 9.42 ± 0.27 * | 37.53 ± 1.59 * | 12.16 ± 0.26 * | 47.73 ± 2.05 * | 3.09 ± 0.07 *# | 1.78 ± 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dubinin, M.V.; Sharapov, V.A.; Semenova, A.A.; Parfenova, L.V.; Ilzorkina, A.I.; Khoroshavina, E.I.; Belosludtseva, N.V.; Gudkov, S.V.; Belosludtsev, K.N. Effect of Modified Levopimaric Acid Diene Adducts on Mitochondrial and Liposome Membranes. Membranes 2022, 12, 866. https://doi.org/10.3390/membranes12090866
Dubinin MV, Sharapov VA, Semenova AA, Parfenova LV, Ilzorkina AI, Khoroshavina EI, Belosludtseva NV, Gudkov SV, Belosludtsev KN. Effect of Modified Levopimaric Acid Diene Adducts on Mitochondrial and Liposome Membranes. Membranes. 2022; 12(9):866. https://doi.org/10.3390/membranes12090866
Chicago/Turabian StyleDubinin, Mikhail V., Vyacheslav A. Sharapov, Alena A. Semenova, Lyudmila V. Parfenova, Anna I. Ilzorkina, Ekaterina I. Khoroshavina, Natalia V. Belosludtseva, Sergey V. Gudkov, and Konstantin N. Belosludtsev. 2022. "Effect of Modified Levopimaric Acid Diene Adducts on Mitochondrial and Liposome Membranes" Membranes 12, no. 9: 866. https://doi.org/10.3390/membranes12090866
APA StyleDubinin, M. V., Sharapov, V. A., Semenova, A. A., Parfenova, L. V., Ilzorkina, A. I., Khoroshavina, E. I., Belosludtseva, N. V., Gudkov, S. V., & Belosludtsev, K. N. (2022). Effect of Modified Levopimaric Acid Diene Adducts on Mitochondrial and Liposome Membranes. Membranes, 12(9), 866. https://doi.org/10.3390/membranes12090866