

Discovery of the Potentiator of the Pore-Forming Ability of Lantibiotic Nisin: Perspectives for Anticancer Therapy


Abstract
1. Introduction
2. Materials and Methods
2.1. Chemical Reagents
2.2. Reconstitution of Nisin into Planar Lipid Bilayers
2.3. Measurements of the Changes in the Membrane Boundary and Its Component Dipole Potential
2.4. Calcein Assay
2.5. Cell Culture
2.6. Mitochondrial Potential
2.7. Statistical Analysis
3. Results and Discussion
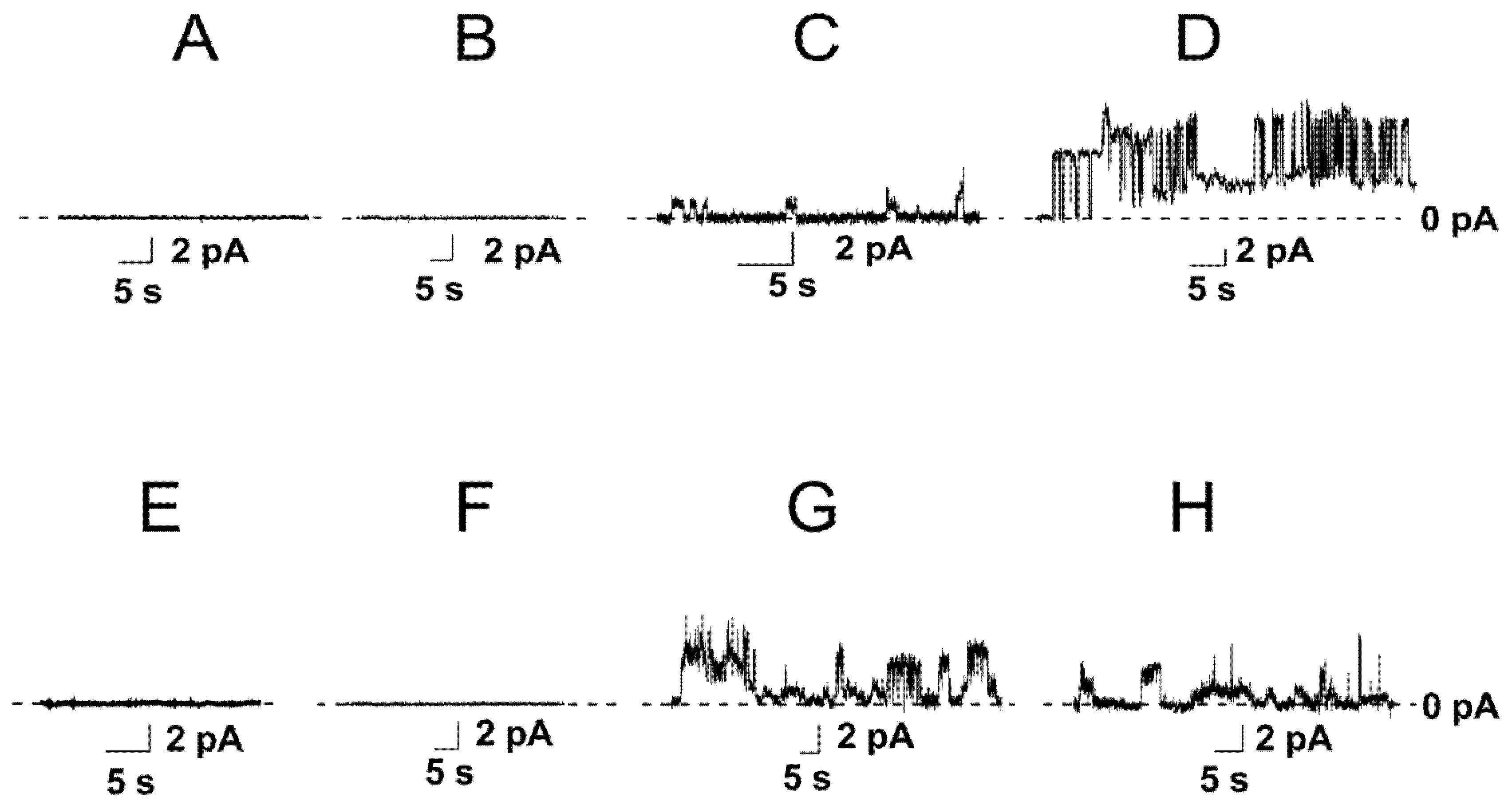
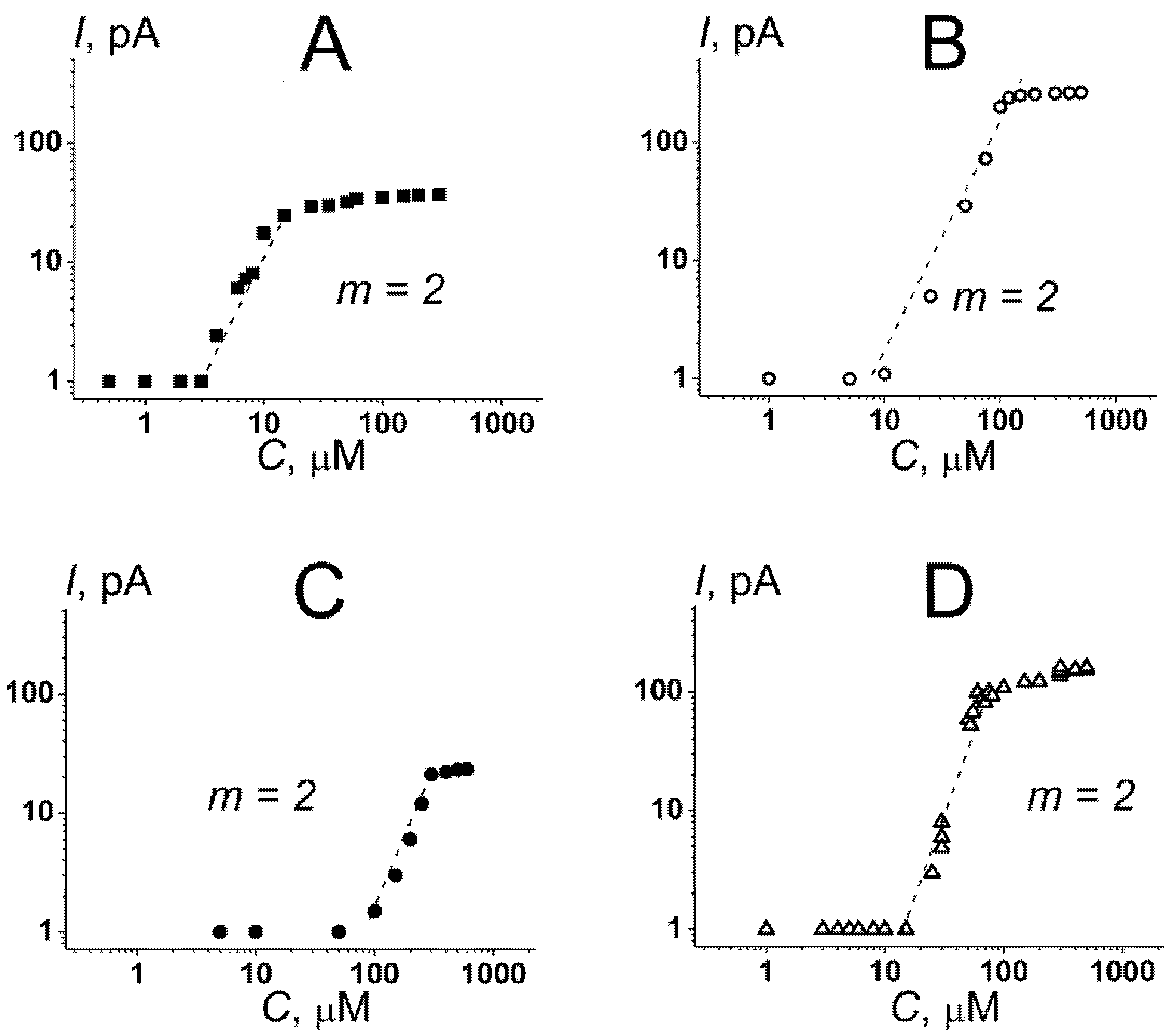
3.1. The Role of Negatively Charged Phospholipids in the Pore-Forming Ability of Nisin
- (1)
- The maximum leakage of the marker from vesicles composed of DOPC/DOPG (50/50 mol%) and DOPC/TOCL (50/50 mol%) was significantly higher than of DOPC and DOPC/DOPS (50/50 mol%) (Supplementary Materials, Figure S1 and Table S1). The results were consistent with the data of electrophysiological measurements (Figure 1).
- (2)
- The time dependences in DOPC/DOPG (50/50 mol%) and DOPC/TOCL (50/50 mol%) bilayers were described by double-exponential functions instead of single-exponential functions used to fit the dependences in DOPC and DOPC/DOPS (50/50 mol%) membranes (Supplementary Materials, Figure S1 and Table S1). A biphasic nature of the curves was consistent with two different mechanisms of nisin action on phosphatidylglycerol- and cardiolipin-enriched membranes: in addition to the detergent action observed for all tested systems, nisin was able to form the pores exceptionally in DOPC/DOPG (50/50 mol%) and DOPC/TOCL (50/50 mol%) bilayers (Figure 1C,D).
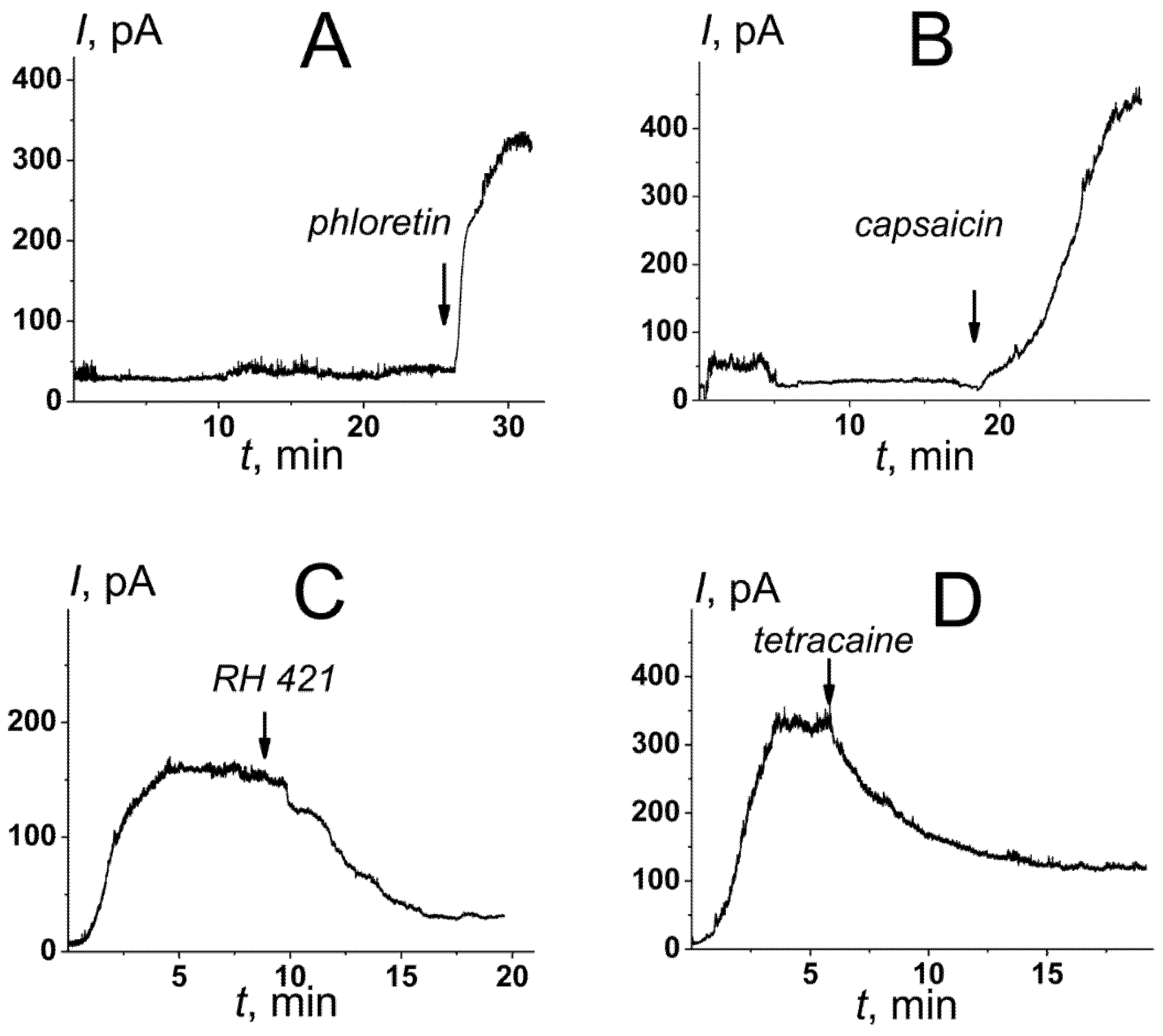
3.2. Small Molecules Potentiating the Activity of Nisin in Cardiolipin-Containing Bilayers
4. Conclusions
- (i)
- The observed decrease in the threshold concentration of nisin in the DOPS-containing bilayers compared to a membrane composed of DOPC might indicate that the antitumor action of the lantibiotic might involve the nisin-induced increase in the permeability of the plasma membrane of malignant cells at phosphatidylserine externalization.
- (ii)
- The pore-formation by nisin in DOPG- and TOCL-enriched membranes might partially explain lantibiotic antimicrobial effects.
- (iii)
- The pronounced pore-forming ability of nisin in TOCL-containing membranes might be a cause of its apoptotic action through mitochondrial pathway.
- (iv)
- The ability of nisin to form pores depends on the distribution of the electrical potential on the membrane-solution interface: a decrease in the membrane boundary potential is accompanied by an increase in the pore-forming activity of the lantibiotic.
- (v)
- Phloretin and capsaicin potentiate the pore-forming ability of nisin in TOCL-enriched membranes. The compound diminishing membrane dipole potential might be potential agonists of the antitumor action of nisin that creates the perspectives to develop innovative lantibiotic formulations.
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Minko, T.; Rodriguez-Rodriguez, L.; Pozharov, V. Nanotechnology approaches for personalized treatment of multidrug resistant cancers. Adv. Drug Deliv. Rev. 2013, 65, 1880–1895. [Google Scholar] [CrossRef]
- Kartal-Yandim, M.; Adan-Gokbulut, A.; Baran, Y. Molecular mechanisms of drug resistance and its reversal in cancer. Crit. Rev. Biotechnol. 2016, 36, 716–726. [Google Scholar] [CrossRef]
- Dagogo-Jack, I.; Shaw, A.T. Tumour heterogeneity and resistance to cancer therapies. Nat. Rev. Clin. Oncol. 2018, 15, 81–94. [Google Scholar] [CrossRef]
- Utsugi, T.; Schroit, A.J.; Connor, J.; Bucana, C.D.; Fidler, I.J. Elevated expression of phosphatidylserine in the outer membrane leaflet of human tumor cells and recognition by activated human blood monocytes. Cancer Res. 1991, 51, 3062–3066. [Google Scholar]
- Hoskin, D.W.; Ramamoorthy, A. Studies on anticancer activities of antimicrobial peptides. Biochim. Biophys. Acta 2008, 1778, 357–375. [Google Scholar] [CrossRef] [PubMed]
- Schweizer, F. Cationic amphiphilic peptides with cancer-selective toxicity. Eur. J. Pharmacol. 2009, 625, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Riedl, S.; Rinner, B.; Asslaber, M.; Schaider, H.; Walzer, S.; Novak, A.; Lohner, K.; Zweytick, D. In search of a novel target–phosphatidylserine exposed by non-apoptotic tumor cells and metastases of malignancies with poor treatment efficacy. Biochim. Biophys. Acta 2011, 1808, 2638–2645. [Google Scholar] [CrossRef] [PubMed]
- Joo, N.E.; Ritchie, K.; Kamarajan, P.; Miao, D.; Kapila, Y.L. Nisin, an apoptogenic bacteriocin and food preservative, attenuates HNSCC tumorigenesis via CHAC1. Cancer Med. 2012, 1, 295–305. [Google Scholar] [CrossRef]
- Gross, E.; Morell, J.L. The structure of nisin. J. Am. Chem. Soc. 1971, 93, 4634–4635. [Google Scholar] [CrossRef]
- Paiva, A.D.; de Oliveira, M.D.; de Paula, S.O.; Baracat-Pereira, M.C.; Breukink, E.; Mantovani, H.C. Toxicity of bovicin HC5 against mammalian cell lines and the role of cholesterol in bacteriocin activity. Microbiology 2012, 158 Pt 11, 2851–2858. [Google Scholar] [CrossRef]
- El-Sayed Ibrahim, N.; Morsy, H.; Abdelgwad, M. The comparative effect of nisin and thioridazine as potential anticancer agents on hepatocellular carcinoma. Rep. Biochem. Mol. Biol. 2021, 9, 452–462. [Google Scholar] [CrossRef] [PubMed]
- Maher, S.; McClean, S. Investigation of the cytotoxicity of eukaryotic and prokaryotic antimicrobial peptides in intestinal epithelial cells in vitro. Biochem. Pharmacol. 2006, 71, 1289–1298. [Google Scholar] [CrossRef] [PubMed]
- Kamarajan, P.; Hayami, T.; Matte, B.; Liu, Y.; Danciu, T.; Ramamoorthy, A.; Worden, F.; Kapila, S.; Kapila, Y. Nisin ZP, a Bacteriocin and Food Preservative, Inhibits Head and Neck Cancer Tumorigenesis and Prolongs Survival. PLoS ONE 2015, 10, e0131008. [Google Scholar] [CrossRef] [PubMed]
- Lewies, A.; Wentzel, J.F.; Miller, H.C.; Du Plessis, L.H. The antimicrobial peptide nisin Z induces selective toxicity and apoptotic cell death in cultured melanoma cells. Biochimie 2018, 144, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Prince, A.; Tiwari, A.; Ror, P.; Sandhu, P.; Roy, J.; Jha, S.; Mallick, B.; Akhter, Y.; Saleem, M. Attenuation of neuroblastoma cell growth by nisin is mediated by modulation of phase behavior and enhanced cell membrane fluidity. Phys. Chem. Chem. Phys. 2019, 21, 1980–1987. [Google Scholar] [CrossRef]
- Beloribi-Djefaflia, S.; Vasseur, S.; Guillaumond, F. Lipid metabolic reprogramming in cancer cells. Oncogenesis 2016, 5, e189. [Google Scholar] [CrossRef]
- Paradies, G.; Paradies, V.; De Benedictis, V.; Ruggiero, F.M.; Petrosillo, G. Functional role of cardiolipin in mitochondrial bioenergetics. Biochim. Biophys. Acta 2014, 1837, 408–417. [Google Scholar] [CrossRef]
- Scherer, K.M.; Spille, J.H.; Sahl, H.G.; Grein, F.; Kubitscheck, U. The lantibiotic nisin induces lipid II aggregation, causing membrane instability and vesicle budding. Biophys. J. 2015, 108, 1114–1124. [Google Scholar] [CrossRef]
- Wiedemann, I.; Breukink, E.; van Kraaij, C.; Kuipers, O.P.; Bierbaum, G.; de Kruijff, B.; Sahl, H.G. Specific binding of nisin to the peptidoglycan precursor lipid II combines pore formation and inhibition of cell wall biosynthesis for potent antibiotic activity. J. Biol. Chem. 2001, 276, 1772–1779. [Google Scholar] [CrossRef]
- Brötz, H.; Josten, M.; Wiedemann, I.; Schneider, U.; Götz, F.; Bierbaum, G.; Sahl, H.G. Role of lipid-bound peptidoglycan precursors in the formation of pores by nisin, epidermin and other lantibiotics. Mol. Microbiol. 1998, 30, 317–327. [Google Scholar] [CrossRef]
- Breukink, E.; Wiedemann, I.; van Kraaij, C.; Kuipers, O.P.; Sahl, H.G.; de Kruijff, B. Use of the cell wall precursor lipid II by a pore-forming peptide antibiotic. Science 1999, 286, 2361–2364. [Google Scholar] [CrossRef] [PubMed]
- Hasper, H.E.; de Kruijff, B.; Breukink, E. Assembly and stability of nisin-lipid II pores. Biochemistry 2004, 43, 11567–11575. [Google Scholar] [CrossRef] [PubMed]
- Wiedemann, I.; Benz, R.; Sahl, H.G. Lipid II-mediated pore formation by the peptide antibiotic nisin: A black lipid membrane study. J. Bacteriol. 2004, 186, 3259–3261. [Google Scholar] [CrossRef] [PubMed]
- Breukink, E.; van Kraaij, C.; Demel, R.A.; Siezen, R.J.; Kuipers, O.P.; de Kruijff, B. The C-terminal region of nisin is responsible for the initial interaction of nisin with the target membrane. Biochemistry 1997, 36, 6968–6976. [Google Scholar] [CrossRef]
- el-Jastimi, R.; Lafleur, M. Structural characterization of free and membrane-bound nisin by infrared spectroscopy. Biochim. Biophys. Acta 1997, 1324, 151–158. [Google Scholar] [CrossRef]
- Giffard, C.J.; Dodd, H.M.; Horn, N.; Ladha, S.; Mackie, A.R.; Parr, A.; Gasson, M.J.; Sanders, D. Structure-function relations of variant and fragment nisins studied with model membrane systems. Biochemistry 1997, 36, 3802–3810. [Google Scholar] [CrossRef] [PubMed]
- Sahl, H.G.; Kordel, M.; Benz, R. Voltage-dependent depolarization of bacterial membranes and artificial lipid bilayers by the peptide antibiotic nisin. Arch. Microbiol. 1987, 149, 120–124. [Google Scholar] [CrossRef]
- Lins, L.; Ducarme, P.; Breukink, E.; Brasseur, R. Computational study of nisin interaction with model membrane. Biochim. Biophys. Acta 1999, 1420, 111–120. [Google Scholar] [CrossRef]
- Montal, M.; Muller, P. Formation of bimolecular membranes from lipid monolayers and study of their electrical properties. Proc. Nat. Acad. Sci. USA 1972, 65, 3561–3566. [Google Scholar] [CrossRef]
- Gawrisch, K.; Ruston, D.; Zimmerberg, J.; Parsegian, V.A.; Rand, R.P.; Fuller, N. Membrane dipole potentials, hydration forces, and the ordering of water at membrane surfaces. Biophys. J. 1992, 61, 1213–1223. [Google Scholar] [CrossRef]
- Ermakov, Y.A.; Sokolov, V.S. Boundary potentials of bilayer lipid membranes: Methods and interpretations. In The Planar Lipid Bilayers (BLMs) and Their Applications; Tien, H.T., Ottova-Leitmannova, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2003; pp. 109–141. [Google Scholar]
- Ermakov, Y.A.; Nesterenko, A.M. Boundary potential of lipid bilayers: Methods and interpretations. J. Phys. Conf. Ser. 2017, 780, 012002. [Google Scholar] [CrossRef]
- Andersen, O.S.; Finkelstein, A.; Katz, I.; Cass, A. Effect of phloretin on the permeability of thin lipid membranes. J. Gen. Physiol. 1976, 67, 749–771. [Google Scholar] [CrossRef] [PubMed]
- Gross, E.; Bedlack, R.S.; Loew, L.M. Dual-wavelength ratiometric fluorescence measurement of the membrane dipole potential. Biophys. J. 1994, 67, 208–216. [Google Scholar] [CrossRef]
- Starke-Peterkovic, T.; Turner, N.; Else, P.L.; Clarke, R.J. Electric field strength of membrane lipids from vertebrate species: Membrane lipid composition and Na+-K+-ATPase molecular activity. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R663–R670. [Google Scholar] [CrossRef] [PubMed]
- Clarke, R.J.; Kane, D.J. Optical detection of membrane dipole potential: Avoidance of fluidity and dye-induced effects. Biochim. Biophys. Acta 1997, 1323, 223–239. [Google Scholar] [CrossRef]
- Clarke, R.J. The dipole potential of phospholipid membranes and methods for its detection. Adv. Colloid Interface Sci. 2001, 89–90, 263–281. [Google Scholar] [CrossRef]
- Fiske, C.H.; Subbarow, Y. The colorimetric determination of phosphorus. J. Biol. Chem. 1925, 66, 375–400. [Google Scholar] [CrossRef]
- Chulkov, E.G.; Ostroumova, O.S. Phloretin modulates the rate of channel formation by polyenes. Biochim. Biophys. Acta Biomembranes 2016, 1858, 289–294. [Google Scholar] [CrossRef]
- Harayama, T.; Riezman, H. Understanding the diversity of membrane lipid composition. Nat. Rev. Mol. Cell Biol. 2018, 19, 281–296. [Google Scholar] [CrossRef]
- Driessen, A.J.; van den Hooven, H.W.; Kuiper, W.; van de Kamp, M.; Sahl, H.G.; Konings, R.N.; Konings, W.N. Mechanistic studies of lantibiotic-induced permeabilization of phospholipid vesicles. Biochemistry 1995, 34, 1606–1614. [Google Scholar] [CrossRef]
- Martin, I.; Ruysschaert, J.M.; Sanders, D.; Giffard, C.J. Interaction of the lantibiotic nisin with membranes revealed by fluorescence quenching of an introduced tryptophan. Eur. J. Biochem. 1996, 239, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wu, S.; Cen, Z.; Zhang, Y.; Chen, Y.; Huang, Y.; Cillo, A.R.; Prokopec, J.S.; Quarato, G.; Vignali, D.A.A.; et al. Mobilizing phospholipids on tumor plasma membrane implicates phosphatidylserine externalization blockade for cancer immunotherapy. Cell Rep. 2022, 41, 111582. [Google Scholar] [CrossRef] [PubMed]
- Blanco, V.M.; Chu, Z.; Vallabhapurapu, S.D.; Sulaiman, M.K.; Kendler, A.; Rixe, O.; Warnick, R.E.; Franco, R.S.; Qi, X. Phosphatidylserine-selective targeting and anticancer effects of SapC-DOPS nanovesicles on brain tumors. Oncotarget 2014, 5, 7105–7118. [Google Scholar] [CrossRef] [PubMed]
- Birge, R.B.; Boeltz, S.; Kumar, S.; Carlson, J.; Wanderley, J.; Calianese, D.; Barcinski, M.; Brekken, R.A.; Huang, X.; Hutchins, J.T.; et al. Phosphatidylserine is a global immunosuppressive signal in efferocytosis, infectious disease, and cancer. Cell Death. Differ. 2016, 23, 962–978. [Google Scholar] [CrossRef] [PubMed]
- Nagata, S.; Suzuki, J.; Segawa, K.; Fujii, T. Exposure of phosphatidylserine on the cell surface. Cell Death Differ. 2016, 23, 952–961. [Google Scholar] [CrossRef] [PubMed]
- Parsons, J.B.; Rock, C.O. Bacterial lipids: Metabolism and membrane homeostasis. Prog. Lipid Res. 2013, 52, 249–276. [Google Scholar] [CrossRef]
- Sohlenkamp, C.; Geiger, O. Bacterial membrane lipids: Diversity in structures and pathways. FEMS Microbiol. Rev. 2016, 40, 133–159. [Google Scholar] [CrossRef]
- Schlame, M. Cardiolipin synthesis for the assembly of bacterial and mitochondrial membranes. J. Lipid Res. 2008, 49, 1607–1620. [Google Scholar] [CrossRef]
- Verheul, A.; Russell, N.J.; Van’T Hof, R.; Rombouts, F.M.; Abee, T. Modifications of membrane phospholipid composition in nisin-resistant Listeria monocytogenes Scott A. Appl. Environ. Microbiol. 1997, 63, 3451–3457. [Google Scholar] [CrossRef]
- Crandall, A.D.; Montville, T.J. Nisin resistance in Listeria monocytogenes ATCC 700302 is a complex phenotype. Appl. Environ. Microbiol. 1998, 64, 231–237. [Google Scholar] [CrossRef]
- Sorice, M.; Circella, A.; Misasi, R.; Pittoni, V.; Garofalo, T.; Cirelli, A.; Pavan, A.; Pontieri, G.M.; Valesini, G. Cardiolipin on the surface of apoptotic cells as a possible trigger for antiphospholipids antibodies. Clin. Exp. Immunol. 2000, 122, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Sorice, M.; Circella, A.; Cristea, I.M.; Garofalo, T.; Di Renzo, L.; Alessandri, C.; Valesini, G.; Esposti, M.D. Cardiolipin and its metabolites move from mitochondria to other cellular membranes during death receptor-mediated apoptosis. Cell Death Differ. 2004, 11, 1133–1145. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.P.; Yoon, J.H.; Long, N.P.; Koo, G.B.; Noh, H.J.; Oh, S.J.; Lee, S.B.; Kim, H.M.; Hong, J.Y.; Lee, W.J.; et al. Spheroid-Induced Epithelial-Mesenchymal Transition Provokes Global Alterations of Breast Cancer Lipidome: A Multi-Layered Omics Analysis. Front. Oncol. 2019, 9, 145. [Google Scholar] [CrossRef]
- Stafford, J.H.; Thorpe, P.E. Increased exposure of phosphatidylethanolamine on the surface of tumor vascular endothelium. Neoplasia 2011, 13, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Broughton, L.J.; Crow, C.; Maraveyas, A.; Madden, L.A. Duramycin-induced calcium release in cancer cells. Anticancer Drugs 2016, 27, 173–182. [Google Scholar] [CrossRef]
- Lewis, R.N.; Zhang, Y.P.; McElhaney, R.N. Calorimetric and spectroscopic studies of the phase behavior and organization of lipid bilayer model membranes composed of binary mixtures of dimyristoylphosphatidylcholine and dimyristoylphosphatidylglycerol. Biochim. Biophys. Acta 2005, 1668, 203–214. [Google Scholar] [CrossRef]
- Frias, M.; Benesch, M.G.; Lewis, R.N.; McElhaney, R.N. On the miscibility of cardiolipin with 1,2-diacyl phosphoglycerides: Binary mixtures of dimyristoylphosphatidylethanolamine and tetramyristoylcardiolipin. Biochim. Biophys. Acta 2011, 1808, 774–783. [Google Scholar] [CrossRef]
- Sennato, S.; Bordi, F.; Cametti, C.; Coluzza, C.; Desideri, A.; Rufini, S. Evidence of domain formation in cardiolipin-glycerophospholipid mixed monolayers. A thermodynamic and AFM study. J. Phys. Chem. B 2005, 109, 15950–15957. [Google Scholar] [CrossRef]
- Lupi, S.; Perla, A.; Maselli, P.; Bordi, F.; Sennato, S. Infrared spectra of phosphatidylethanolamine-cardiolipin binary system. Colloids Surf. B Biointerfaces 2008, 64, 56–64. [Google Scholar] [CrossRef]
- Boscia, A.L.; Treece, B.W.; Mohammadyani, D.; Klein-Seetharaman, J.; Braun, A.R.; Wassenaar, T.A.; Klösgen, B.; Tristram-Nagle, S. X-ray structure, thermodynamics, elastic properties and MD simulations of cardiolipin/dimyristoylphosphatidylcholine mixed membranes. Chem. Phys. Lipids 2014, 178, 1–10. [Google Scholar] [CrossRef]
- Epand, R.M. Anionic Lipid Clustering Model. Adv. Exp. Med. Biol. 2019, 1117, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Efimova, S.S.; Ostroumova, O.S. Effect of dipole modifiers on the magnitude of the dipole potential of sterol-containing bilayers. Langmuir 2012, 28, 9908–9914. [Google Scholar] [CrossRef] [PubMed]
- Efimova, S.S.; Zakharova, A.A.; Ostroumova, O.S. Alkaloids modulate the functioning of ion channels produced by antimicrobial agents via an influence on the lipid host. Front. Cell Dev. Biol. 2020, 8, 537. [Google Scholar] [CrossRef] [PubMed]
- Efimova, S.S.; Zakharova, A.A.; Schagina, L.V.; Ostroumova, O.S. Local anesthetics affect gramicidin a channels via membrane electrostatic potentials. J. Membr. Biol. 2016, 249, 781–787. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Lipid Composition | Pore-Forming Ability | Detergent Activity | |||
|---|---|---|---|---|---|
| isc, pA | Ctr, μM | C|Imax, μM | Imax, pA | Cdet, μM | |
| DOPC | – | – | – | – | 2000 ± 50 |
| DOPC/DOPS (50/50 mol%) | – | – | – | – | 700 ± 10 |
| DOPC/DOPG (50/50 mol%) | 0.5 ÷ 1.0 | 5 ± 2 | 25 ± 3 | 30 ± 8 | 750 ± 20 |
| DOPC/TOCL (50/50 mol%) | 2 ÷ 29 | 10 ± 3 | 100 ± 5 | 250 ± 20 | 610 ± 10 |
| DOPE | – | – | – | – | 2050 ± 50 |
| DOPE/DOPS (50/50 mol%) | – | – | – | – | 600 ± 5 |
| DOPE/DOPG (50/50 mol%) | 1 ÷ 18 | 65 ± 5 | 310 ±15 | 30 ± 5 | 650 ± 10 |
| DOPE/TOCL (50/50 mol%) | 1 ÷ 27 | 12 ± 3 | 80 ± 15 | 195 ± 10 | 550 ± 15 |
| C, μM | Small Molecules | pKa # | ρ, % # | Δφb, mV | Δφd, mV | Iagent/I0controla |
|---|---|---|---|---|---|---|
| 20 | phloretin | 7.96 | −23 | −86 ± 5 | −99 ± 11 | 5.3 ± 1.3 |
| 400 | capsaicin | 4.25 | 0 | −95 ± 11 | −80 ± 12 | 11.3 ± 1.9 |
| 5 | RH 421 | 5.96 | −3 | 106 ± 11 | –* | 0.4 ± 0.1 |
| 500 | tetracaine | 8.42 | +91 | 51 ± 6 | 4 ± 2 | 0.6 ± 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chernyshova, D.N.; Tyulin, A.A.; Ostroumova, O.S.; Efimova, S.S. Discovery of the Potentiator of the Pore-Forming Ability of Lantibiotic Nisin: Perspectives for Anticancer Therapy. Membranes 2022, 12, 1166. https://doi.org/10.3390/membranes12111166
Chernyshova DN, Tyulin AA, Ostroumova OS, Efimova SS. Discovery of the Potentiator of the Pore-Forming Ability of Lantibiotic Nisin: Perspectives for Anticancer Therapy. Membranes. 2022; 12(11):1166. https://doi.org/10.3390/membranes12111166
Chicago/Turabian StyleChernyshova, Dayana N., Alexander A. Tyulin, Olga S. Ostroumova, and Svetlana S. Efimova. 2022. "Discovery of the Potentiator of the Pore-Forming Ability of Lantibiotic Nisin: Perspectives for Anticancer Therapy" Membranes 12, no. 11: 1166. https://doi.org/10.3390/membranes12111166
APA StyleChernyshova, D. N., Tyulin, A. A., Ostroumova, O. S., & Efimova, S. S. (2022). Discovery of the Potentiator of the Pore-Forming Ability of Lantibiotic Nisin: Perspectives for Anticancer Therapy. Membranes, 12(11), 1166. https://doi.org/10.3390/membranes12111166