
The Interaction between hERG1 and β1 Integrins Modulates hERG1 Current in Different Pathological Cell Models
 , ,
, , 

{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Coatings
2.3. Patch Clamp Recording
2.4. Statistical Analysis
3. Results
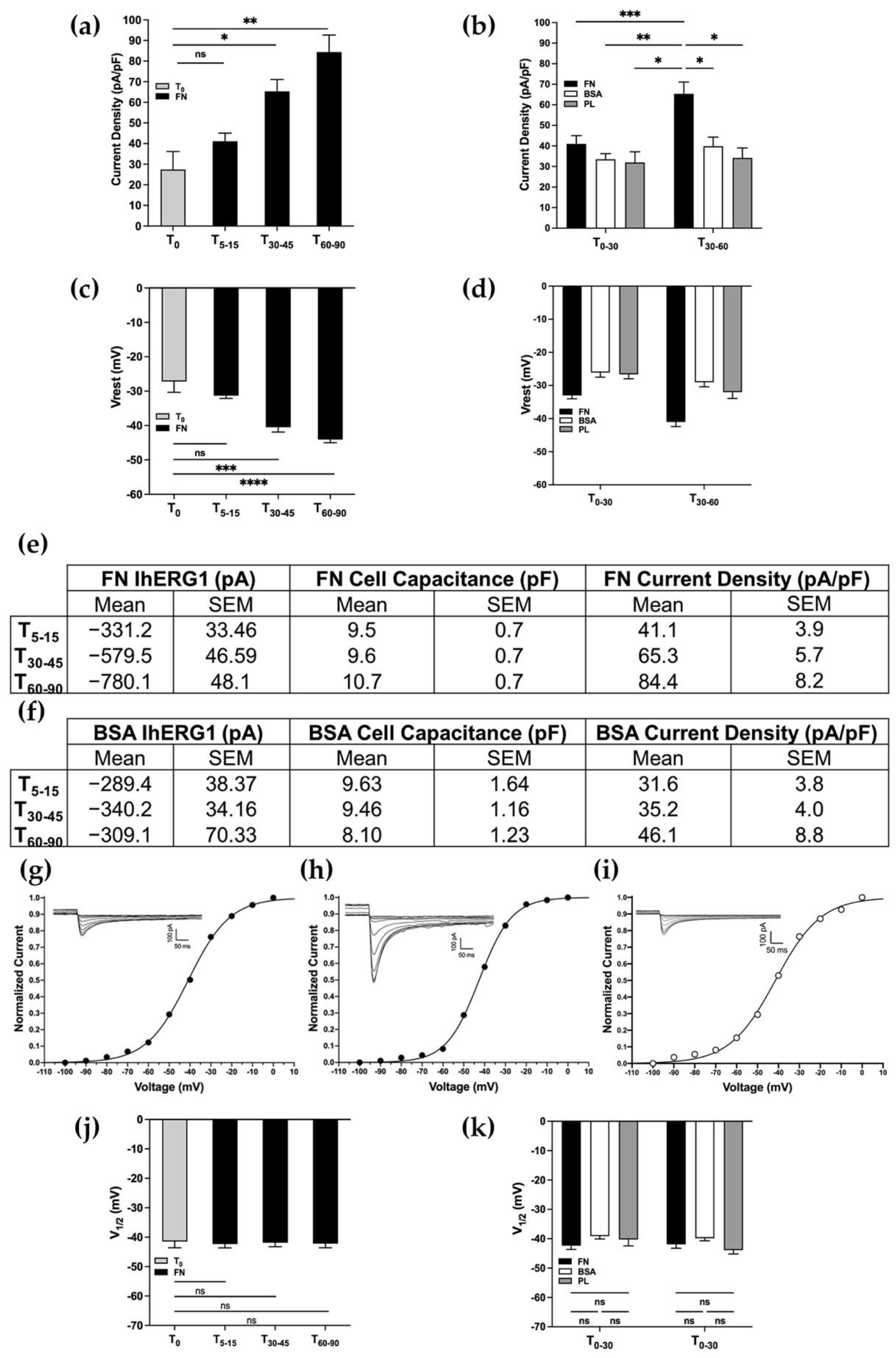
3.1. β1 Integrin Activation Leads to hERG1 Current Density Increase and Resting Membrane Potential Hyperpolarization: A Kinetic Alteration?
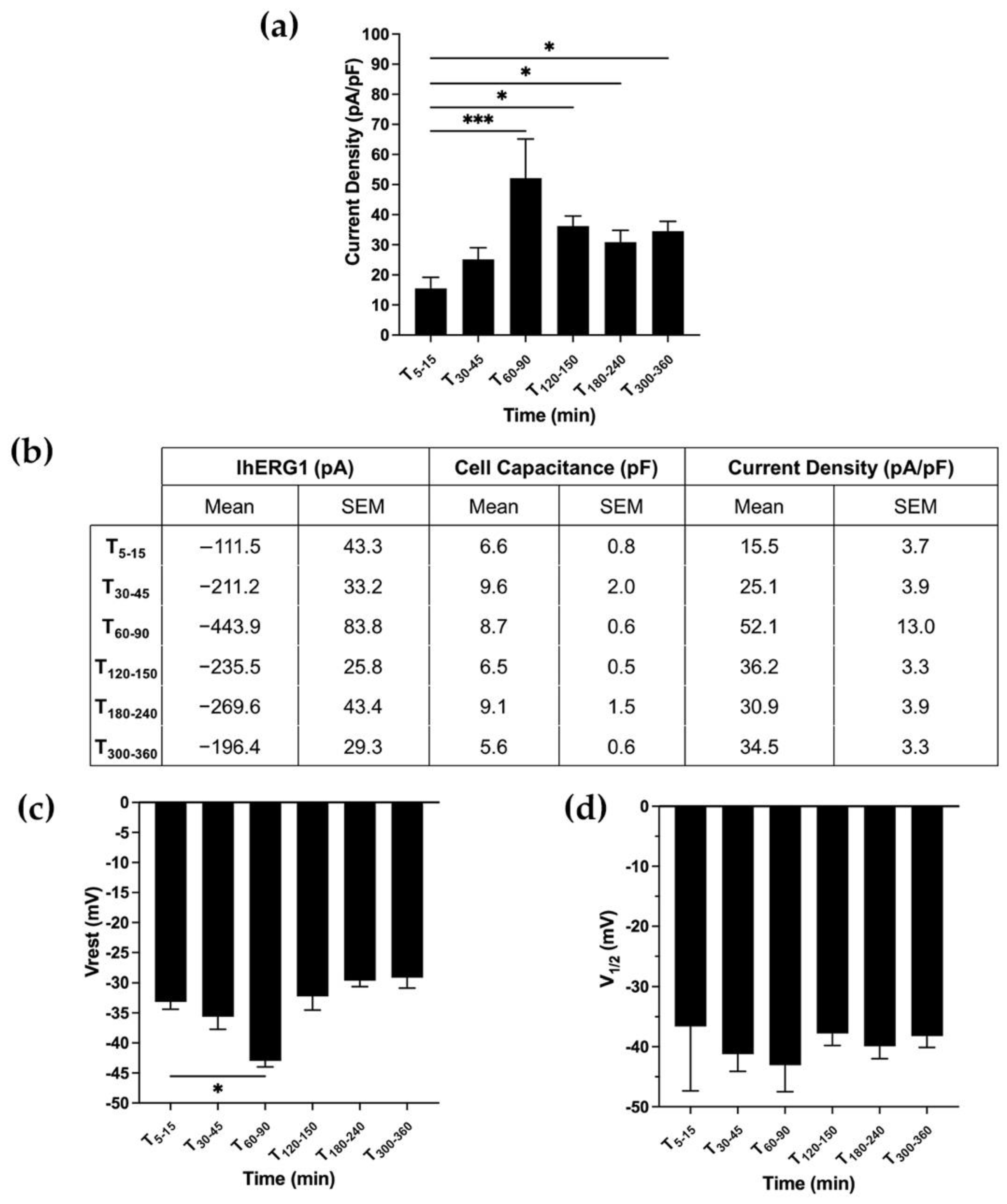
3.2. hERG1-β1 Interaction in a Neuroblastoma Cell Line
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Djamgoz, M.B.A.; Coombes, R.C.; Schwab, A. Ion Transport and Cancer: From Initiation to Metastasis. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130092. [Google Scholar] [CrossRef] [PubMed]
- Prevarskaya, N.; Skryma, R.; Shuba, Y. Ion Channels in Cancer: Are Cancer Hallmarks Oncochannelopathies? Physiol. Rev. 2018, 98, 559–621. [Google Scholar] [CrossRef] [PubMed]
- Becchetti, A.; Crescioli, S.; Zanieri, F.; Petroni, G.; Mercatelli, R.; Coppola, S.; Gasparoli, L.; D’Amico, M.; Pillozzi, S.; Crociani, O.; et al. The Conformational State of HERG1 Channels Determines Integrin Association, Downstream Signaling, and Cancer Progression. Sci. Signal. 2017, 10, eaaf3236. [Google Scholar] [CrossRef]
- Becchetti, A.; Petroni, G.; Arcangeli, A. Ion Channel Conformations Regulate Integrin-Dependent Signaling. Trends Cell Biol. 2019, 29, 298–307. [Google Scholar] [CrossRef]
- Arcangeli, A.; Becchetti, A. Complex Functional Interaction between Integrin Receptors and Ion Channels. Trends Cell Biol. 2006, 16, 631–639. [Google Scholar] [CrossRef]
- Duranti, C.; Arcangeli, A. Ion Channel Targeting with Antibodies and Antibody Fragments for Cancer Diagnosis. Antibodies 2019, 8, 33. [Google Scholar] [CrossRef] [PubMed]
- Arcangeli, A.; Bianchi, L.; Becchetti, A.; Faravelli, L.; Coronnello, M.; Mini, E.; Olivotto, M.; Wanke, E. A Novel Inward-Rectifying K+ Current with a Cell-Cycle Dependence Governs the Resting Potential of Mammalian Neuroblastoma Cells. J. Physiol. 1995, 489 Pt 2, 455–471. [Google Scholar] [CrossRef]
- Arcangeli, A. Expression and Role of HERG Channels in Cancer Cells. Novartis Found. Symp. 2005, 266, 225–232, discussion 232–234. [Google Scholar]
- Huang, X.; Jan, L.Y. Targeting Potassium Channels in Cancer. J. Cell Biol. 2014, 206, 151–162. [Google Scholar] [CrossRef]
- Feng, J.; Yu, J.; Pan, X.; Li, Z.; Chen, Z.; Zhang, W.; Wang, B.; Yang, L.; Xu, H.; Zhang, G.; et al. HERG1 Functions as an Oncogene in Pancreatic Cancer and Is Downregulated by MiR-96. Oncotarget 2014, 5, 5832–5844. [Google Scholar] [CrossRef]
- Arcangeli, A.; Rosati, B.; Crociani, O.; Cherubini, A.; Fontana, L.; Passani, B.; Wanke, E.; Olivotto, M. Modulation of HERG Current and Herg Gene Expression during Retinoic Acid Treatment of Human Neuroblastoma Cells: Potentiating Effects of BDNF. J. Neurobiol. 1999, 40, 214–225. [Google Scholar] [CrossRef]
- Arcangeli, A.; Becchetti, A. HERG Channels: From Antitargets to Novel Targets for Cancer Therapy. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2017, 23, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Sanguinetti, M.C.; Jiang, C.; Curran, M.E.; Keating, M.T. A Mechanistic Link between an Inherited and an Acquired Cardiac Arrhythmia: HERG Encodes the IKr Potassium Channel. Cell 1995, 81, 299–307. [Google Scholar] [CrossRef]
- Arcangeli, A.; Becchetti, A. Novel Perspectives in Cancer Therapy: Targeting Ion Channels. Drug Resist. Updates 2015, 21–22, 11–19. [Google Scholar] [CrossRef]
- Manoli, S.; Coppola, S.; Duranti, C.; Lulli, M.; Magni, L.; Kuppalu, N.; Nielsen, N.; Schmidt, T.; Schwab, A.; Becchetti, A.; et al. The Activity of Kv 11.1 Potassium Channel Modulates F-Actin Organization During Cell Migration of Pancreatic Ductal Adenocarcinoma Cells. Cancers 2019, 11, 135. [Google Scholar] [CrossRef]
- Maneshi, P.; Mason, J.; Dongre, M.; Öhlund, D. Targeting Tumor-Stromal Interactions in Pancreatic Cancer: Impact of Collagens and Mechanical Traits. Front. Cell Dev. Biol. 2021, 9, 787485. [Google Scholar] [CrossRef]
- Lastraioli, E.; Perrone, G.; Sette, A.; Fiore, A.; Crociani, O.; Manoli, S.; D’Amico, M.; Masselli, M.; Iorio, J.; Callea, M.; et al. HERG1 Channels Drive Tumour Malignancy and May Serve as Prognostic Factor in Pancreatic Ductal Adenocarcinoma. Br. J. Cancer 2015, 112, 1076–1087. [Google Scholar] [CrossRef]
- Arcangeli, A.; Becchetti, A.; Mannini, A.; Mugnai, G.; Fifippi, P.D.; Tarone, G. Integrin-Mediated Neurite Outgrowth in Neuroblastoma Cells Depends on the Activation of Potassium Channels. J. Cell Biol. 1993, 122, 13. [Google Scholar] [CrossRef]
- Arcangeli, A.; Faravelli, L.; Bianchi, L.; Rosati, B.; Gritti, A.; Vescovi, A.; Wanke, E.; Olivotto, M. Soluble or Bound Laminin Elicit in Human Neuroblastoma Cells Short- or Long-Term Potentiation of a K+ Inwardly Rectifying Current: Relevance to Neuritogenesis. Cell Adhes. Commun. 1996, 4, 369–385. [Google Scholar] [CrossRef]
- Cherubini, A.; Hofmann, G.; Pillozzi, S.; Guasti, L.; Crociani, O.; Cilia, E.; Di Stefano, P.; Degani, S.; Balzi, M.; Olivotto, M.; et al. Human Ether-a-Go-Go -Related Gene 1 Channels Are Physically Linked to β1 Integrins and Modulate Adhesion-Dependent Signaling. Mol. Biol. Cell 2005, 16, 2972–2983. [Google Scholar] [CrossRef]
- Cherubini, A.; Pillozzi, S.; Hofmann, G.; Crociani, O.; Guasti, L.; Lastraioli, E.; Polvani, S.; Masi, A.; Becchetti, A.; Wanke, E.; et al. HERG K+ Channels and Beta1 Integrins Interact through the Assembly of a Macromolecular Complex. Ann. N. Y. Acad. Sci. 2002, 973, 559–561. [Google Scholar] [CrossRef] [PubMed]
- Cohen-Kupiec, R.; Weinstein, S.; Kantor, G.; Peer, D.; Weil, M. IKAP/HELP1 down-Regulation in Neuroblastoma Cells Causes Enhanced Cell Adhesion Mediated by Contactin Overexpression. Cell Adhes. Migr. 2010, 4, 541–550. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Leventhal, P.S.; Feldman, E.L. Tyrosine Phosphorylation and Enhanced Expression of Paxillin during Neuronal Differentiation in Vitro. J. Biol. Chem. 1996, 271, 5957–5960. [Google Scholar] [CrossRef] [PubMed]
- Becchetti, A.; Duranti, C.; Arcangeli, A. Dynamics and Physiological Meaning of Complexes between Ion Channels and Integrin Receptors: The Case of Kv11.1. Am. J. Physiol. Cell Physiol. 2022, 322, C1138–C1150. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.B.; Dransfield, I. Electric Fields and Inflammation: May the Force Be with You. Sci. World J. 2008, 8, 1280–1294. [Google Scholar] [CrossRef][Green Version]
- Olivotto, M.; Arcangeli, A.; Carlà, M.; Wanke, E. Electric Fields at the Plasma Membrane Level: A Neglected Element in the Mechanisms of Cell Signalling. BioEssays News Rev. Mol. Cell. Dev. Biol. 1996, 18, 495–504. [Google Scholar] [CrossRef]
- Brown, G.C. Electrostatic Coupling between Membrane Proteins. FEBS Lett. 1990, 260, 1–5. [Google Scholar] [CrossRef]
- Yang, M.; Brackenbury, W.J. Membrane Potential and Cancer Progression. Front. Physiol. 2013, 4, 185. [Google Scholar] [CrossRef]
- Canady, K.S.; Ali-Osman, F.; Rubel, E.W. Extracellular Potassium Influences DNA and Protein Syntheses and Glial Fibrillary Acidic Protein Expression in Cultured Glial Cells. Glia 1990, 3, 368–374. [Google Scholar] [CrossRef]
- Boonstra, J.; Mummery, C.L.; Tertoolen, L.G.J.; Van Der Saag, P.T.; De Laat, S.W. Cation Transport and Growth Regulation in Neuroblastoma Cells. Modulations of K+ Transport and Electrical Membrane Properties during the Cell Cycle. J. Cell. Physiol. 1981, 107, 75–83. [Google Scholar] [CrossRef]
- Wonderlin, W.F.; Woodfork, K.A.; Strobl, J.S. Changes in Membrane Potential during the Progression of MCF-7 Human Mammary Tumor Cells through the Cell Cycle. J. Cell. Physiol. 1995, 165, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Price, M.; Lee, S.C.; Deutsch, C. Charybdotoxin Inhibits Proliferation and Interleukin 2 Production in Human Peripheral Blood Lymphocytes. Proc. Natl. Acad. Sci. USA 1989, 86, 10171–10175. [Google Scholar] [CrossRef] [PubMed]
- Freedman, B.D.; Price, M.A.; Deutsch, C.J. Evidence for Voltage Modulation of IL-2 Production in Mitogen-Stimulated Human Peripheral Blood Lymphocytes. J. Immunol. 1992, 149, 3784–3794. [Google Scholar] [PubMed]
- Wang, Y.F.; Jia, H.; Walker, A.M.; Cukierman, S. K-Current Mediation of Prolactin-Induced Proliferation of Malignant (Nb2) Lymphocytes. J. Cell. Physiol. 1992, 152, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Mills, B.; Tupper, J.T. Cell Cycle Dependent Changes in Potassium Transport. J. Cell. Physiol. 1976, 89, 123–132. [Google Scholar] [CrossRef]
- Arcangeli, A.; Rosati, B.; Cherubini, A.; Crociani, O.; Fontana, L.; Ziller, C.; Wanke, E.; Olivotto, M. HERG- and IRK-like Inward Rectifier Currents Are Sequentially Expressed during Neuronal Development of Neural Crest Cells and Their Derivatives. Eur. J. Neurosci. 1997, 9, 2596–2604. [Google Scholar] [CrossRef] [PubMed]
- Crociani, O.; Cherubini, A.; Piccini, E.; Polvani, S.; Costa, L.; Fontana, L.; Hofmann, G.; Rosati, B.; Wanke, E.; Olivotto, M.; et al. Erg Gene(s) Expression during Development of the Nervous and Muscular System of Quail Embryos. Mech. Dev. 2000, 95, 239–243. [Google Scholar] [CrossRef]
- Bianchi, L.; Wible, B.; Arcangeli, A.; Taglialatela, M.; Morra, F.; Castaldo, P.; Crociani, O.; Rosati, B.; Faravelli, L.; Olivotto, M.; et al. Herg Encodes a K+ Current Highly Conserved in Tumors of Different Histogenesis: A Selective Advantage for Cancer Cells? Cancer Res. 1998, 58, 815–822. [Google Scholar]
- Wang, H.; Zhang, Y.; Cao, L.; Han, H.; Wang, J.; Yang, B.; Nattel, S.; Wang, Z. HERG K+ Channel, a Regulator of Tumor Cell Apoptosis and Proliferation. Cancer Res. 2002, 62, 4843–4848. [Google Scholar]
- Duranti, C.; Carraresi, L.; Sette, A.; Stefanini, M.; Lottini, T.; Crescioli, S.; Crociani, O.; Iamele, L.; De Jonge, H.; Gherardi, E.; et al. Generation and Characterization of Novel Recombinant Anti-HERG1 ScFv Antibodies for Cancer Molecular Imaging. Oncotarget 2018, 9, 34972–34989. [Google Scholar] [CrossRef]
- Iorio, J.; Duranti, C.; Lottini, T.; Lastraioli, E.; Bagni, G.; Becchetti, A.; Arcangeli, A. KV11.1 Potassium Channel and the Na+/H+ Antiporter NHE1 Modulate Adhesion-Dependent Intracellular PH in Colorectal Cancer Cells. Front. Pharmacol. 2020, 11, 848. [Google Scholar] [CrossRef] [PubMed]
- Duranti, C.; Iorio, J.; Lottini, T.; Lastraioli, E.; Crescioli, S.; Bagni, G.; Lulli, M.; Capitani, C.; Bouazzi, R.; Stefanini, M.; et al. Harnessing the HERG1/Β1 Integrin Complex via a Novel Bispecific Single-Chain Antibody: An Effective Strategy against Solid Cancers. Mol. Cancer Ther. 2021, 20, 1338–1349. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chioccioli Altadonna, G.; Montalbano, A.; Iorio, J.; Becchetti, A.; Arcangeli, A.; Duranti, C. The Interaction between hERG1 and β1 Integrins Modulates hERG1 Current in Different Pathological Cell Models. Membranes 2022, 12, 1162. https://doi.org/10.3390/membranes12111162
Chioccioli Altadonna G, Montalbano A, Iorio J, Becchetti A, Arcangeli A, Duranti C. The Interaction between hERG1 and β1 Integrins Modulates hERG1 Current in Different Pathological Cell Models. Membranes. 2022; 12(11):1162. https://doi.org/10.3390/membranes12111162
Chicago/Turabian StyleChioccioli Altadonna, Ginevra, Alberto Montalbano, Jessica Iorio, Andrea Becchetti, Annarosa Arcangeli, and Claudia Duranti. 2022. "The Interaction between hERG1 and β1 Integrins Modulates hERG1 Current in Different Pathological Cell Models" Membranes 12, no. 11: 1162. https://doi.org/10.3390/membranes12111162
APA StyleChioccioli Altadonna, G., Montalbano, A., Iorio, J., Becchetti, A., Arcangeli, A., & Duranti, C. (2022). The Interaction between hERG1 and β1 Integrins Modulates hERG1 Current in Different Pathological Cell Models. Membranes, 12(11), 1162. https://doi.org/10.3390/membranes12111162