
The Structural Changes in the Membranes of Staphylococcus aureus Caused by Hydrolysable Tannins Witness Their Antibacterial Activity

 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Bacterial Strain and Growth Conditions
2.3. Antimicrobial Activity
2.3.1. Determination of Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC)
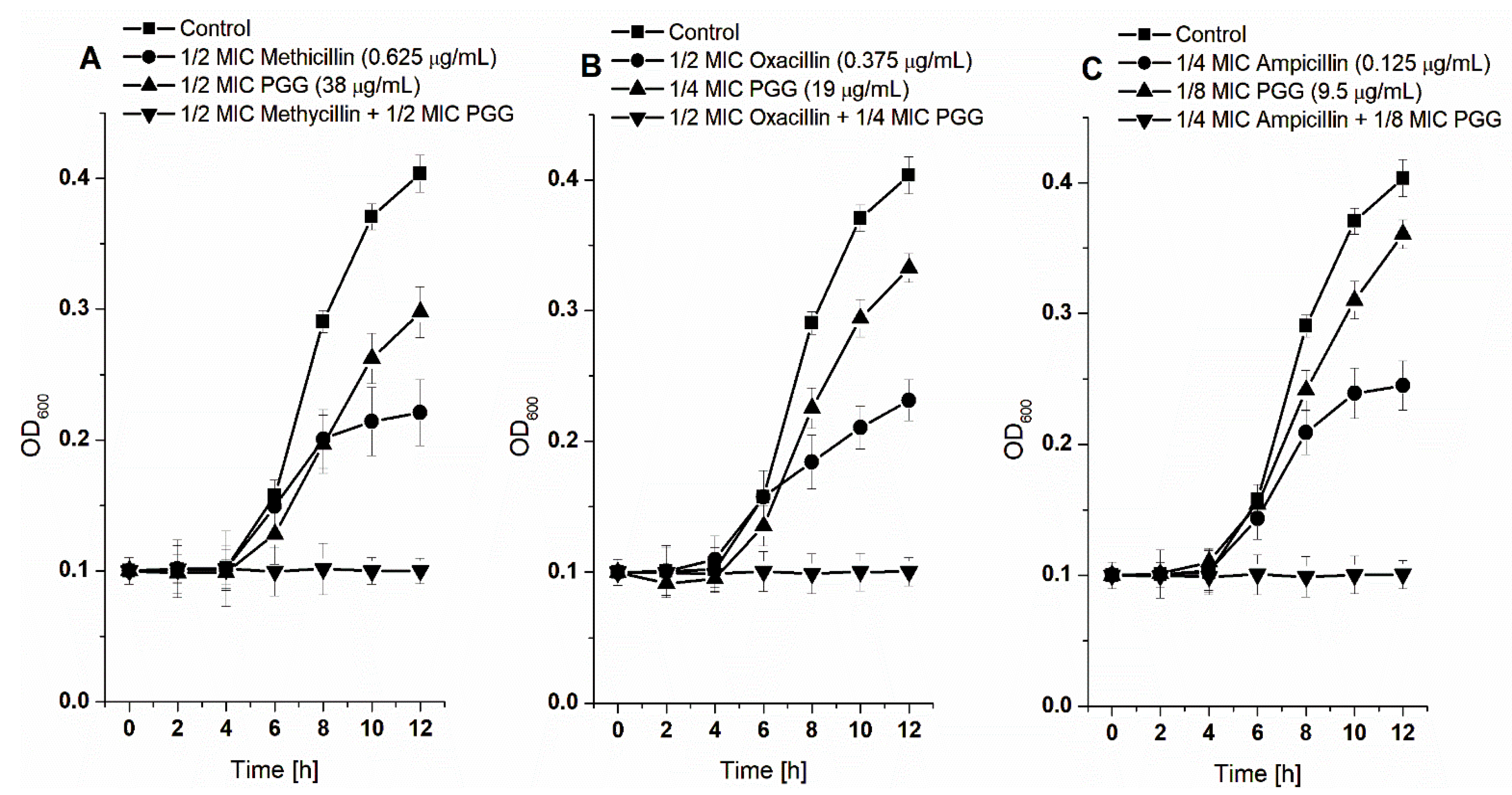
2.3.2. Synergic Effects between PGG and β-Lactam Antibiotics against S. aureus
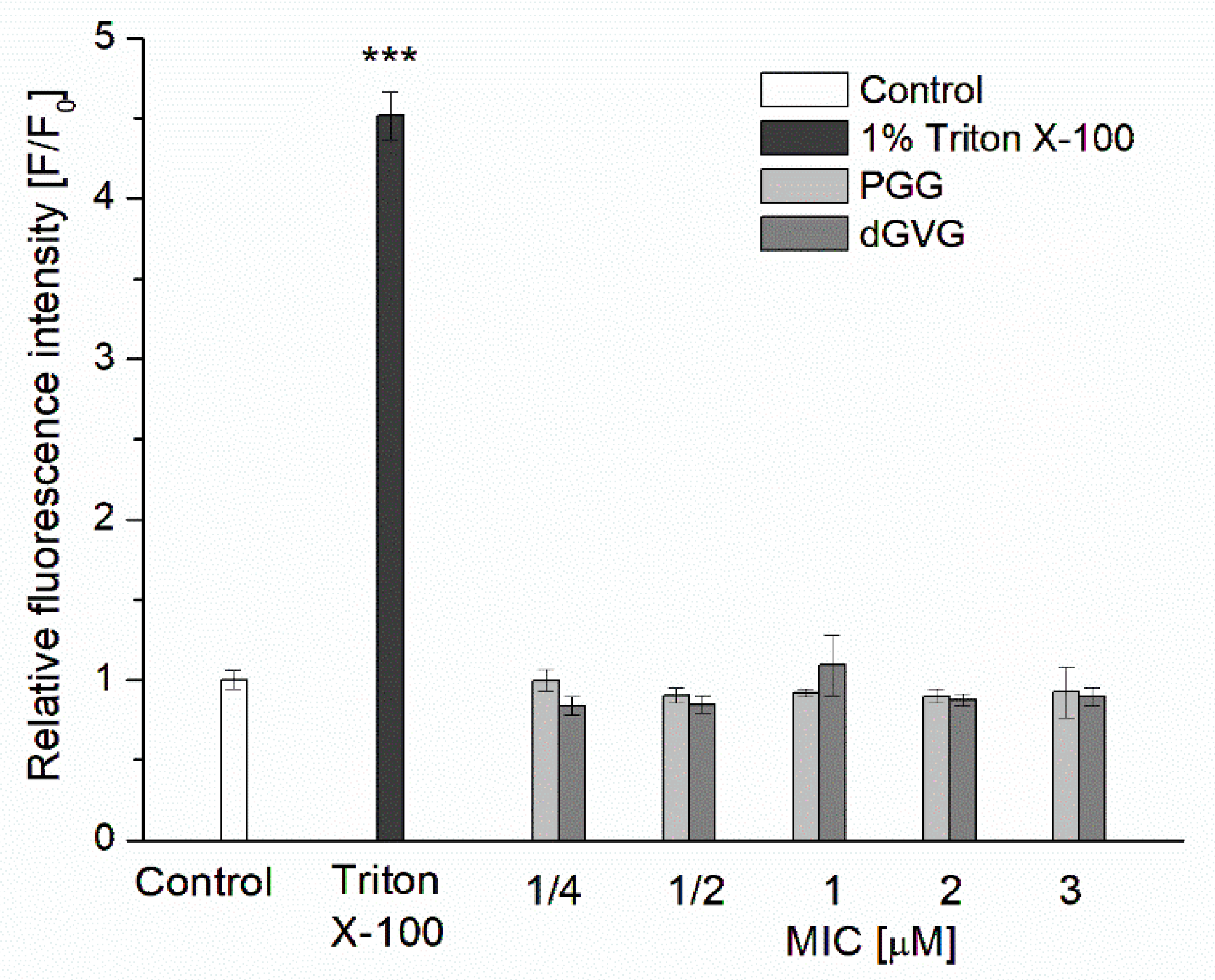
2.4. Bacterial Cell Membrane Permeability Assay
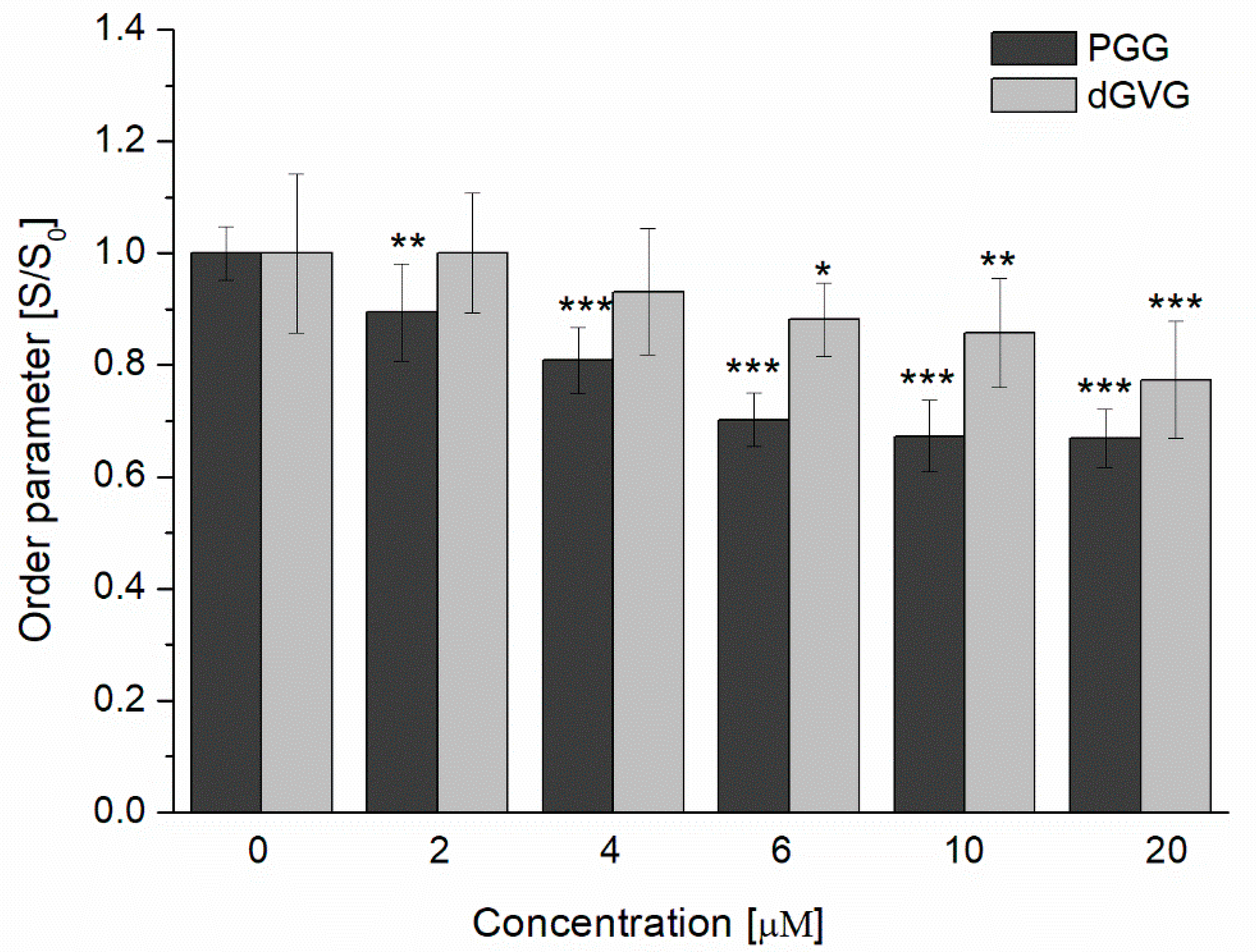
2.5. Measurements of S. aureus Membrane Fluidity
2.6. Detection of Tannin-Bacterial Membrane Interaction by Measuring DPH Quenching of Fluorescence
2.7. Fluorescence Analysis of Membrane Proteins of S. aureus—Tannin Interaction
2.8. Isolation of Naturally Secreted MVs
2.9. Statistical Analysis
3. Results and Discussion
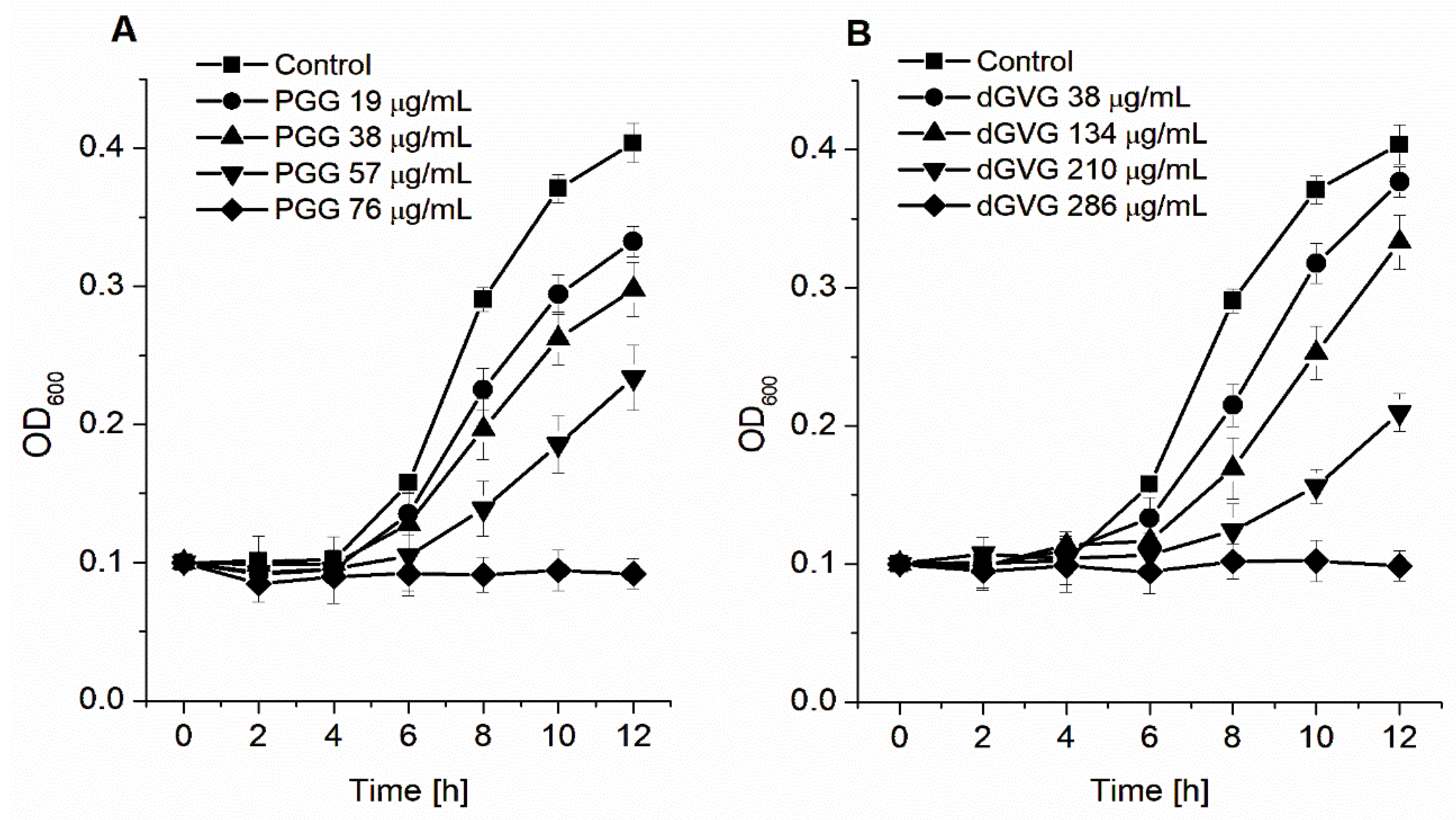
3.1. Antibacterial Activities of the Studied Tannins, Applied Alone and in Combination with Antibiotics
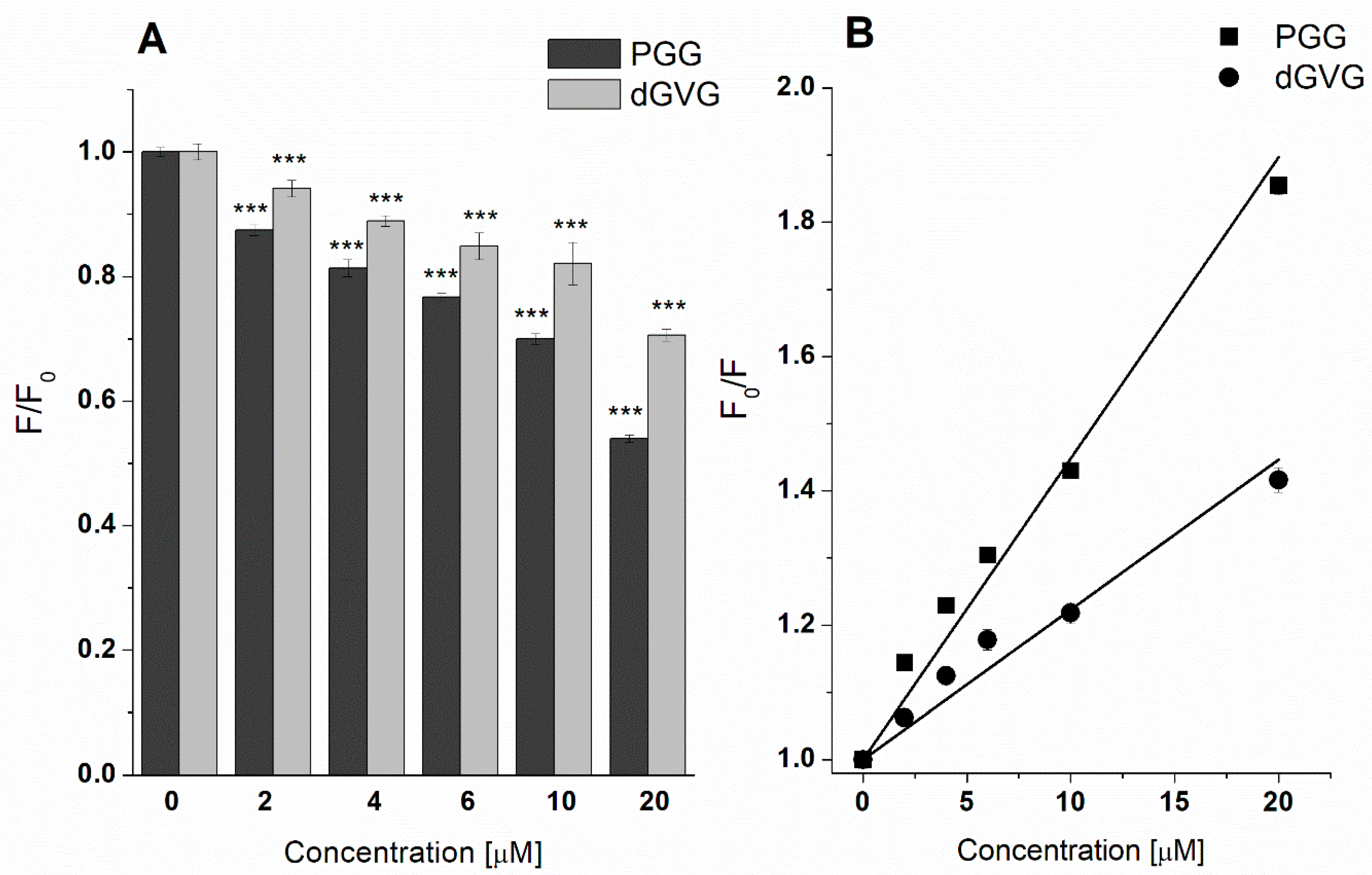
3.2. Influence of PGG and dGVG on the S. aureus Cell Membrane—Fluorescence Studies
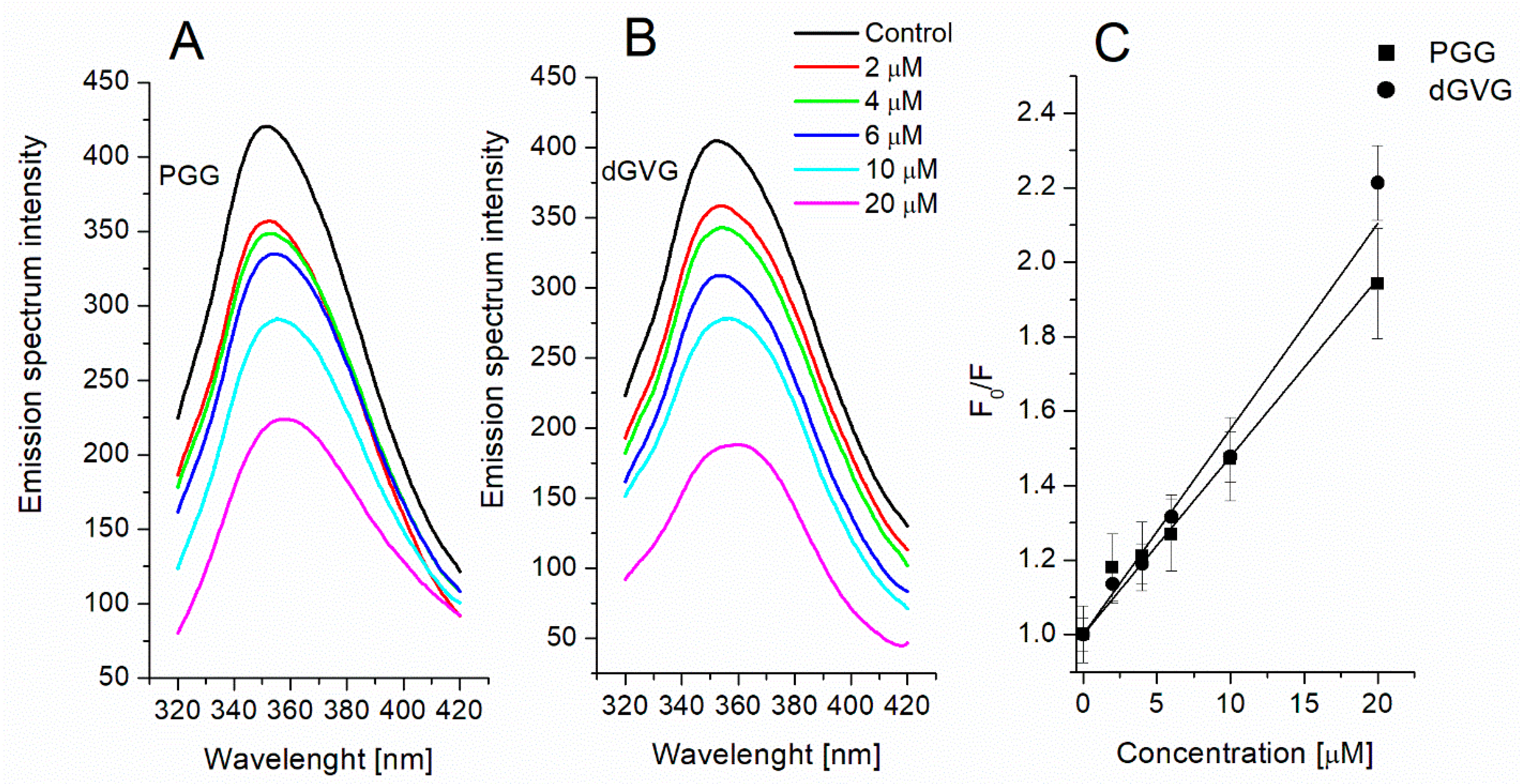
3.3. Evaluation of Interaction of Tannins with Membrane Proteins by Tryptophan Fluorescence
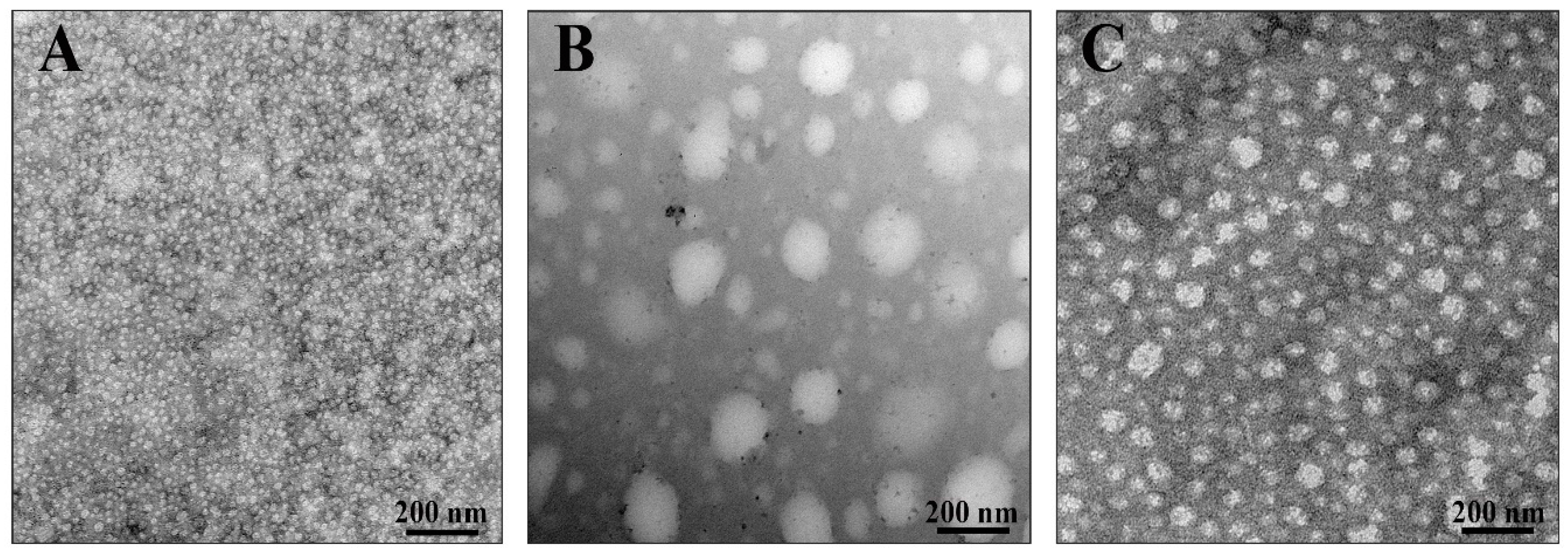
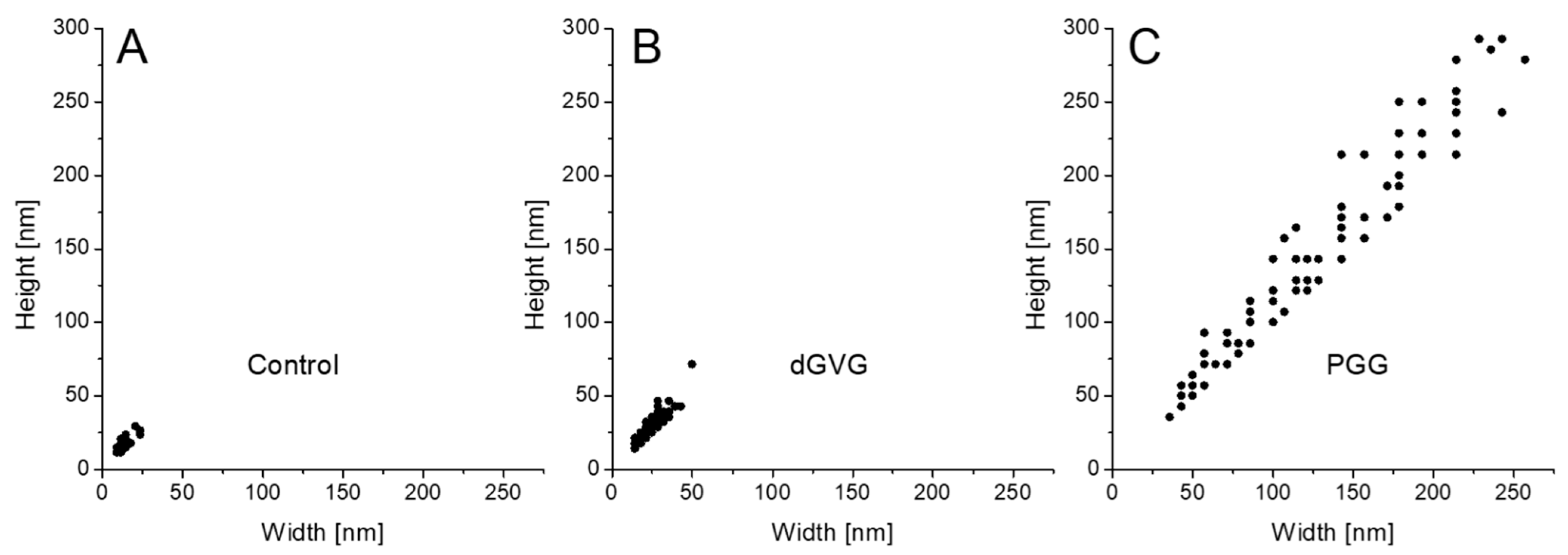
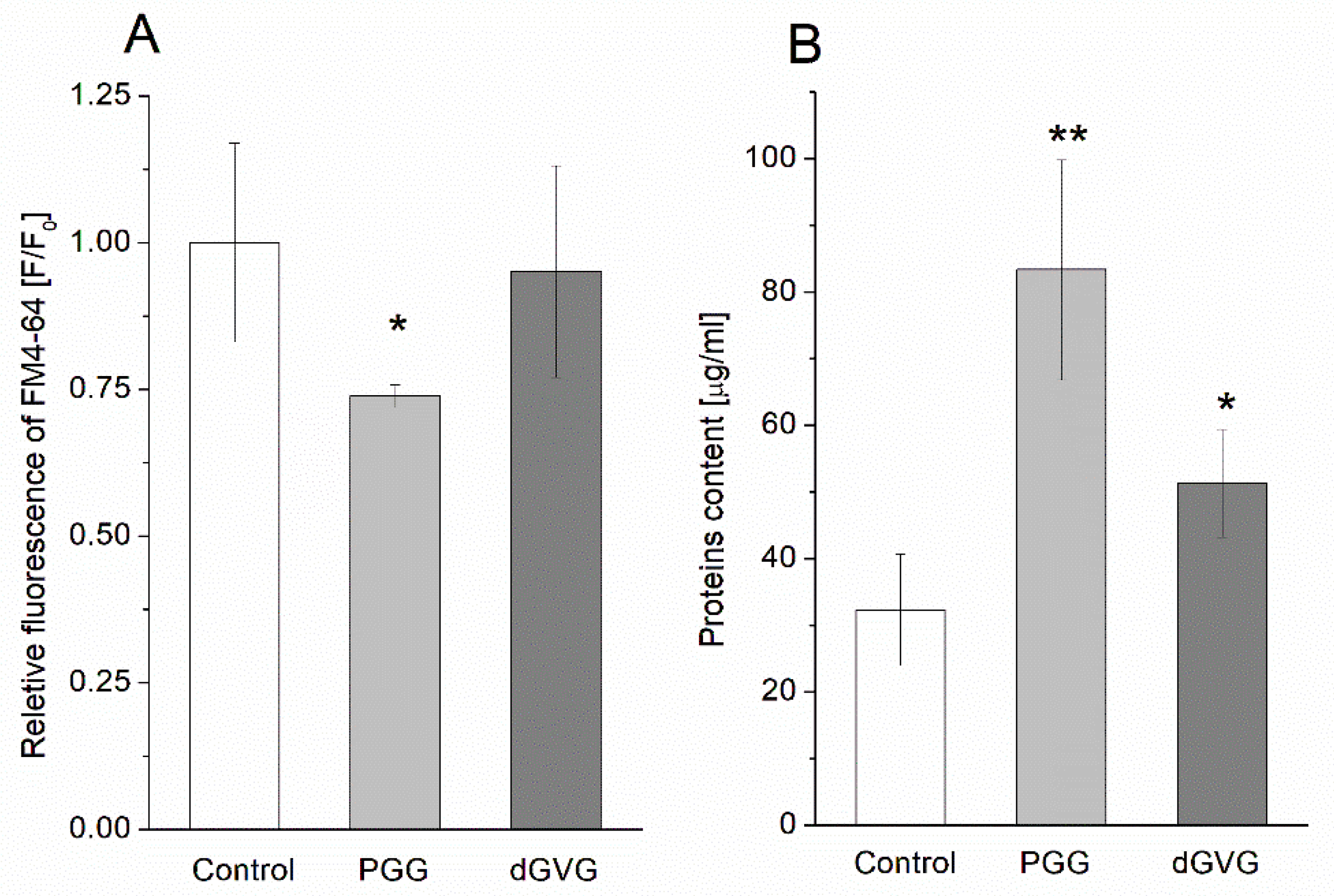
3.4. The Influence Tannins on Characteristics of Membrane Vesicles (MVs) Isolated from S. aureus
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Papuc, K.; Goran, G.V.; Predescu, C.N.; Nicorescu, V.; Stefan, G. Plant Polyphenols as antioxidant and antibacterial agents for shelf-life extension of meat and meat products: Classification, structures, sources, and action mechanisms. Compr. Rev. Food Sci. Food Saf. 2017, 16, 1243–1268. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.; Bhardwaj, K.; Cruz-Martins, N.; Nepovimova, E.; Oleksak, P.; Dhanjal, D.S.; Bhardwaj, S.; Singh, R.; Chopra, C.; Verma, R.; et al. Applications of fruit polyphenols and their functionalized nanoparticles against foodborne bacteria: A mini review. Molecules 2021, 26, 3447. [Google Scholar] [CrossRef] [PubMed]
- Makarewicz, M.; Drozdz, I.; Tarko, T.; Duda-Chodak, A. The interactions between polyphenols and microorganisms, especially gut microbiota. Antioxidants 2021, 28, 188. [Google Scholar] [CrossRef] [PubMed]
- Serrano, J.; Puupponen–Pimia, R.; Dauer, A.; Aura, A.M.; Saura–Calixto, F. Tannins: Current knowledge of food sources, intake, bioavailability and biological effects. Mol. Nutr. Food Res. 2009, 53, 310–329. [Google Scholar] [CrossRef]
- Girard, M.; Bee, G. Invited review: Tannins as a potential alternative to antibiotics to prevent coliform diarrhea in weaned pigs. Animal 2020, 14, 95–107. [Google Scholar] [CrossRef]
- Maatsola, S.; Kurkinen, S.; Engstrom, M.T.; Nyholm, T.K.M.; Pentikainen, O.; Salminen, J.P.; Haataja, S. Inhibition of pneumolysin cytotoxicity by hydrolysable tannins. Antibiotics 2020, 21, 930. [Google Scholar] [CrossRef]
- Sieniawska, E. Activities of tannins—From in vitro studies to clinical trials. Nat. Prod. Commun. 2015, 10, 1877–1884. [Google Scholar] [CrossRef]
- Redondo, L.M.; Chacana, P.A.; Dominguez, E.J.; Miyakawa, M.E.F. Perspectives in the use of tannins as alternative to antimicrobial growth promoter factors in poultry. Front. Microbiol. 2014, 75, 118. [Google Scholar] [CrossRef]
- Yilmaz, H.; Subasi, B.G.; Celebioglu, H.U.; Ozdal, T.; Capanoglu, E. Chemistry of protein-phenolic interactions toward the microbiota and microbial infections. Front. Nutr. 2022, 9, 914118. [Google Scholar] [CrossRef]
- Gupta, P.D.; Birdi, T.J. Development of botanicals to combat antibiotic resistance. J. Ayurveda Integr. Med. 2017, 8, 266–275. [Google Scholar] [CrossRef]
- Oliveira, D.; Borges, A.; Simoes, M. Staphylococcus aureus toxins and their molecular activity in infectious diseases. Toxins 2018, 10, 252. [Google Scholar] [CrossRef] [PubMed]
- Olchowik-Grabarek, E.; Sekowski, S.; Bitiucki, M.; Dobrzynska, I.; Shlyonsky, V.; Ionov, M.; Burzynski, P.; Roszkowska, A.; Swiecicka, I.; Abdulladjanova, N.; et al. Inhibition of interaction between Staphylococcus aureus α-hemolysin and erythrocytes membrane by hydrolysable tannins: Structure-related activity study. Sci. Rep. 2020, 10, 11168. [Google Scholar] [CrossRef] [PubMed]
- Barrajon-Catalan, E.; Fernandez-Arroyo, S.; Saura, D.; Guillen, E.; Fernendez-Gutierrez, A.; Segura-Carretero, A.; Micol, V. Cistaceae aqueous extracts containing ellagitannins show antioxidant and antimicrobial capacity, and cytotoxic activity against human cancer cells. Food Chem. Toxicol. 2010, 48, 2273–2282. [Google Scholar] [CrossRef] [PubMed]
- Quave, C.L.; Estevez-Carmona, M.; Compadre, C.M.; Hobby, G.; Hendrickson, H.; Beenken, K.E.; Smeltzer, M.S. Ellagic acid derivatives from Rubus ulmifolius inhibit Staphylococcus aureus biofilm formation and improve response to antibiotics. PLoS ONE 2012, 7, e28737. [Google Scholar] [CrossRef] [PubMed]
- Payne, D.E.; Martin, N.R.; Parzych, K.R.; Rickard, A.H.; Underwood, A.; Boles, B.R. Tannic acid inhibits Staphylococcus aureus surface colonization in an IsaA-dependent manner. Infect. Immun. 2013, 81, 496–504. [Google Scholar] [CrossRef]
- Eom, S.H.; Kim, D.H.; Lee, S.H.; Yoon, N.Y.; Kim, J.H.; Kim, T.H.; Chung, Y.H.; Kim, S.B.; Kim, Y.M.; Kim, H.W.; et al. In vitro antibacterial activity and synergistic antibiotic effects of phlorotannins isolated from Eisenia bicyclis against methicillin-resistant Staphylococcus aureus. Phytother. Res. 2013, 27, 1260–1264. [Google Scholar] [CrossRef] [PubMed]
- Diniz-Silva, H.T.; da Silva Cirino, I.C.; Falcao-Silva, V.D.S.; Magnani, M.; de Souza, E.L.; Siqueira-Junior, J.P. Tannic acid as a potential modulator of norfloxacin resistance in Staphylococcus aureus over expressing NorA. Chemotherapy 2016, 61, 319–322. [Google Scholar] [CrossRef]
- Al-Harbi, R.; Shaaban, M.; Al-Wegaisi, R.; Moharram, F.; El-Rahman, O.A.; El-Messery, S. Antimicrobial activity and molecular docking of tannins from Pimenta dioica. Lett. Drug Des. Discov. 2018, 15, 508–515. [Google Scholar] [CrossRef]
- Puljula, E.; Walton, G.; Woodward, M.J.; Karonen, M. Antimicrobial activities of ellagitannins against Clostridiales perfringens, Escherichia coli, Lactobacillus plantarum and Staphylococcus aureus. Molecules 2020, 25, 3714. [Google Scholar] [CrossRef]
- Norden, C.W.; Wentzel, H.; Keleti, E. Comparison of techniques for measurement of in vitro antibiotic synergism. J. Infect. Dis. 1979, 140, 629–633. [Google Scholar] [CrossRef]
- Kirmusaoglu, S. Sensitizing of β-lactam resistance by tannic acid in methicillin-resistant S. aureus. World J. Microbiol. Biotechnol. 2019, 35, 57. [Google Scholar] [CrossRef] [PubMed]
- Roth, B.L.; Poot, M.; Yue, S.T.; Millard, P.J. Bacterial viability and antibiotic susceptibility testing with SYTOX Green nucleic acid stain. Appl. Environ. Microbiol. 1997, 63, 2421–2431. [Google Scholar] [CrossRef]
- Olchowik, E.; Lotkowski, K.; Mavlyanov, S.; Abdullajanova, N.; Ionov, M.; Bryszewska, M.; Zamaraeva, M. Stabilization of erythrocytes against oxidative and hypotonic stress by tannins isolated from sumac leaves (Rhus typhina L.) and grape seeds (Vitis vinifera L.). Cell Mol. Biol. Lett. 2012, 17, 333–348. [Google Scholar] [CrossRef] [PubMed]
- Sekowski, S.; Veiko, A.; Olchowik-Grabarek, E.; Dubis, A.; Wilczewska, A.; Markiewicz, K.; Zavodnik, I.; Lapshina, E.; Dobrzynska, I.; Abdulladjanova, N.; et al. Hydrolysable tannins change physicochemical parameters of lipid nano-vesicles and reduce DPPH radical—Experimental studies and quantum chemical analysis. Biochim. Biophys. Acta Biomembr. 2022, 1864, 183778. [Google Scholar] [CrossRef] [PubMed]
- Lakowicz, J.R. Principles of fluorescence Spectroscopy, 2nd ed.; Kluwer Academic/Planum Publishers: New York, NY, USA, 1999. [Google Scholar]
- Prados-Rosales, R.; Brown, L.; Casadevall, A.; Montelvo-Qiuros, S.; Luque-Garcia, J. Isolation i identification of membrane vesicle-associated proteins in Gram-positive bacteria and mycobacteria. MethodsX 2014, 1, 124–129. [Google Scholar] [CrossRef]
- Cho, J.J.; Kim, H.S.; Kim, C.H.; Cho, S.J. Interaction with polyphenols and antibiotics. J. Life Sci. 2017, 27, 476–481. [Google Scholar] [CrossRef][Green Version]
- Sanhueza, L.; Melo, R.; Montero, R.; Maisey, K.; Mendoza, L.; Wilkens, M. Synergistic interactions between phenolic compounds identified in grape pomace extract with antibiotics of different classes against Staphylococcus aureus and Escherichia coli. PLoS ONE 2017, 12, e0172273. [Google Scholar] [CrossRef]
- Amin, M.U.; Khurram, M.; Khattak, B.; Khan, J. Antibiotic additive and synergistic action of rutin, morin and quercetin against methicillin resistant Staphylococcus aureus. BMC Complement. Altern. Med. 2015, 15, 59. [Google Scholar] [CrossRef]
- Din, W.M.; Jin, K.T.; Ramli, R.; Khaithir, T.M.N.; Wiart, C. Antibacterial effects of ellagitannins from Acalypha wilkesiana var. macafeana hort.: Surface morphology analysis with environmental scanning electron microscopy and synergy with antibiotics. Phytother. Res. 2013, 27, 1313–1320. [Google Scholar]
- Basri, D.F.; Zin, N.M.; Bakar, N.S.; Rahmat, F.; Mohtar, M. Synergistic effects of phytochemicals and oxacillin on laboratory passagederived vancomycin-intermediate Staphylococcus aureus strain. J. Med. Sci. 2008, 8, 131–136. [Google Scholar]
- Hartmann, M.; Berditsch, M.; Hawecker, J.; Ardakani, M.F.; Gerthsen, D.; Ulrich, A.S. Damage of the bacterial cell envelope by antimicrobial peptides gramicidin S and PGLa as revealed by transmission and scanning electron microscopy. Antimicrob. Agents Chemother. 2010, 54, 3132–3142. [Google Scholar] [CrossRef] [PubMed]
- Mattio, L.M.; Dallavalle, S.; Musso, L.; Filardi, R.; Franzetti, L.; Pellegrino, L.; D’Incecco, P.; Mora, D.; Pinto, A.; Arioli, S. Antimicrobial activity of resveratrol-derived monomers and dimers against foodborne pathogens. Sci. Rep. 2019, 20, 19525. [Google Scholar] [CrossRef] [PubMed]
- Yi, S.M.; Zhu, L.J.; Fu, L.L.; Li, J.R. Tea polyphenols inhibit Pseudomonas aeruginosa through damage to the cell membrane. Int. J. Food Microbiol. 2010, 144, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Trentin, D.S.; Silva, D.B.; Amaral, M.W.; Zimmer, K.R.; Silva, M.V.; Lopes, N.P.; Giordani, R.B.; Macedo, A.J. Tannins possessing bacteriostatic effect impair Pseudomonas aeruginosa adhesion and biofilm formation. PLoS ONE 2013, 8, e66257. [Google Scholar] [CrossRef] [PubMed]
- Yasir, M.; Dutta, D.; Willcox, M.D.P. Mode of action of the antimicrobial peptide Mel4 is independent of Staphylococcus aureus cell membrane permeability. PLoS ONE 2019, 14, e0215703. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Kaihatsu, K.; Nishino, K.; Ogawa, M.; Kato, N.; Yamaguchi, A. Antibacterial and antifungal activities of new acylated derivatives of epigallocatechin gallate. Front. Microbiol. 2012, 3, 53. [Google Scholar] [CrossRef]
- Donadio, G.; Mensitieri, F.; Santoro, V.; Parisi, V.; Bellone, M.L.; De Tommasi, N.; Izzo, V.; Dal Piaz, F. Interactions with microbial proteins driving the antibacterial activity of flavonoids. Pharmaceutics 2021, 13, 660. [Google Scholar] [CrossRef]
- Scalbert, A. Antimicrobial properties of tannins. Phytochemistry 1991, 30, 3875–3883. [Google Scholar] [CrossRef]
- Nassarawa, S.S.; Nayik, G.A.; Gupta, S.D.; Areche, F.O.; Jagdale, Y.D.; Ansari, M.J.; Hemeg, H.A.; Al-Farga, A.; Alotaibi, S.S. Chemical aspects of polyphenol-protein interactions and their antibacterial activity. Crit. Rev. Food Sci. Nutr. 2022, 27, 1–24. [Google Scholar] [CrossRef]
- Wu, Y.; Bai, J.; Zhong, K.; Huang, Y.; Qi, H.; Jiang, Y.; Gao, H. Antibacterial activity and membrane-disruptive mechanism of 3-p-trans-Coumaroyl-2-hydroxyquinic acid, a novel phenolic compound from pine needles of Cedrus deodara, against Staphylococcus aureus. Molecules 2016, 21, 1084. [Google Scholar] [CrossRef]
- Sekowski, S.; Bitiucki, M.; Ionov, M.; Zdeb, M.; Abdulladjanova, N.; Rakhimov, R.; Mavlyanov, S.; Bryszewska, M.; Zamaraeva, M. Influence of valoneoyl groups on the interactions between Euphorbia tannins and human serum albumin. J. Lumin. 2018, 194, 170–178. [Google Scholar] [CrossRef]
- Nasri, R.; Bidel, L.P.R.; Rugani, N.; Perrier, V.; Ceriere, F.; Dubreucq, E.; Jay-Allemand, C. Inhibition of CpLIP2 lipase Hydrolytic activity by four flavonols (galangin, kaempferol, quercetin, myricetin) compared to orlistat and their binding mechanisms studied by quenching of fluorescence. Molecules 2019, 24, 2888. [Google Scholar] [CrossRef]
- Tintino, S.R.; Morais-Tintino, C.D.; Campina, F.F.; Costa, M.D.S.; Menezes, I.R.A.; de Matos, Y.M.L.S.; Calixto-Junior, J.T.; Pereira, P.S.; Siqueira-Junior, J.P.; Leal-Balbino, T.C.; et al. Tannic acid affects the phenotype of Staphylococcus aureus resistant to tetracycline and erythromycin by inhibition of efflux pumps. Bioorg. Chem. 2017, 74, 197–200. [Google Scholar] [PubMed]
- Sinsinwar, S.; Vadivel, V. Catechin isolated from cashew nut shell exhibits antibacterial activity against clinical isolates of MRSA through ROS-mediated oxidative stress. Appl. Microbiol. Biotechnol. 2020, 104, 8279–8297. [Google Scholar] [CrossRef] [PubMed]
- Bose, S.; Aggarwal, S.; Singh, D.V.; Acharya, N. Extracellular vesicles: An emerging platform in gram-positive bacteria. Microb. Cell 2020, 7, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Chudzik, A.; Paściak, M. Bacterial extracellular vesicles as cell-cell communication mediators. Postep. Hig. Med. Dosw. 2020, 74, 572–588. [Google Scholar] [CrossRef]
- Briaud, P.; Carroll, R.K. Extracellular vesicle biogenesis and functions in Gram-positive bacteria. Infect. Immun. 2020, 16, e00433-20. [Google Scholar] [CrossRef]
- Avila-Calderon, E.D.; Araiza-Villanuweva, M.G.; Cancido-Diaz, J.C.; Lopez-Villegas, E.O.; Sriranganathan, N.; Boyle, S.M.; Contreras-Rodriguez, A. Roles of bacterial membrane vesicles. Arch. Microbiol. 2015, 197, 1–10. [Google Scholar] [CrossRef]
- Cao, Y.; Lin, H. Characterization and function of membrane vesicles in Gram-positive bacteria. Appl. Microbiol. Biotechnol. 2021, 105, 1795–1801. [Google Scholar] [CrossRef]
- Nagakubo, T.; Nomuro, N.; Toyofuku, M. Cracking open bacterial membrane vesicles. Front. Microbiol. 2019, 10, 3026. [Google Scholar] [CrossRef]
- Mozaheb, N.; Mingeot-Leclercq, M.P. Membrane vesicle production as a bacterial defense against stress. Front. Microbiol. 2020, 11, 600221. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, H.; Wang, S.; Du, Y.; Wei, B.; Wu, Q.; Wang, H. Inhibitors of bacterial extracellular vesicles. Front. Microbiol. 2022, 13, 835058. [Google Scholar] [CrossRef] [PubMed]
- Palacios, L.; Rosado, H.; Micol, V.; Rosato, A.E.; Bernal, P.; Arroyo, R.; Grounds, H.; Anderson, J.C.; Stabler, R.A.; Taylor, P.W. Staphylococcal phenotypes induced by naturally occurring and synthetic membrane-interactive polyphenolic β-lactam resistance modifiers. PLoS ONE 2014, 9, e93830. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | MIC [μg/mL] | MBC [μg/mL] |
|---|---|---|
| PGG | 76 (80 μM) | 304 (320 μM) |
| dGVG | 286 (300 μM) | >953 (1000 μM) |
| Methicillin | 1.25 | 5 |
| Oxacillin | 0.75 | 3 |
| Ampicillin | 0.5 | 2 |
| Combination | FIC Value | Effect | Range of Combination MICs [μg/mL] | Reduction Fold of MICs |
|---|---|---|---|---|
| PGG-Methicillin | 1 | Additive | 38–0.625 | 2–2 |
| PGG-Oxacillin | 0.75 | Partial synergism | 19–0.375 | 4–2 |
| PGG-Ampicillin | 0.375 | Synergism | 9.5–0.125 | 8–4 |
| Combination | Range of Combination MBCs [μg/mL] | Reduction Fold of MBCs |
|---|---|---|
| PGG–Methicillin | 152–2.5 | 2–2 |
| PGG–Oxacillin | 152–1.5 | 2–2 |
| PGG–Ampicillin | 76–0.5 | 4–4 |
| PGG | dGVG | |
|---|---|---|
| Ksv [M−1] | (4.74 ± 0.94) × 104 | (5.75 ± 0.61) × 104 |
| kq [M−1·s−1] | (9.48 ± 1.88) × 1012 | (11.50 ± 1.21) × 1012 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olchowik-Grabarek, E.; Sękowski, S.; Kwiatek, A.; Płaczkiewicz, J.; Abdulladjanova, N.; Shlyonsky, V.; Swiecicka, I.; Zamaraeva, M. The Structural Changes in the Membranes of Staphylococcus aureus Caused by Hydrolysable Tannins Witness Their Antibacterial Activity. Membranes 2022, 12, 1124. https://doi.org/10.3390/membranes12111124
Olchowik-Grabarek E, Sękowski S, Kwiatek A, Płaczkiewicz J, Abdulladjanova N, Shlyonsky V, Swiecicka I, Zamaraeva M. The Structural Changes in the Membranes of Staphylococcus aureus Caused by Hydrolysable Tannins Witness Their Antibacterial Activity. Membranes. 2022; 12(11):1124. https://doi.org/10.3390/membranes12111124
Chicago/Turabian StyleOlchowik-Grabarek, Ewa, Szymon Sękowski, Agnieszka Kwiatek, Jagoda Płaczkiewicz, Nodira Abdulladjanova, Vadim Shlyonsky, Izabela Swiecicka, and Maria Zamaraeva. 2022. "The Structural Changes in the Membranes of Staphylococcus aureus Caused by Hydrolysable Tannins Witness Their Antibacterial Activity" Membranes 12, no. 11: 1124. https://doi.org/10.3390/membranes12111124
APA StyleOlchowik-Grabarek, E., Sękowski, S., Kwiatek, A., Płaczkiewicz, J., Abdulladjanova, N., Shlyonsky, V., Swiecicka, I., & Zamaraeva, M. (2022). The Structural Changes in the Membranes of Staphylococcus aureus Caused by Hydrolysable Tannins Witness Their Antibacterial Activity. Membranes, 12(11), 1124. https://doi.org/10.3390/membranes12111124