
Rush Hour of LATs towards Their Transport Cycle

 ,
,  ,
, 

Abstract
1. Introduction
2. Structural Information about the APC Family
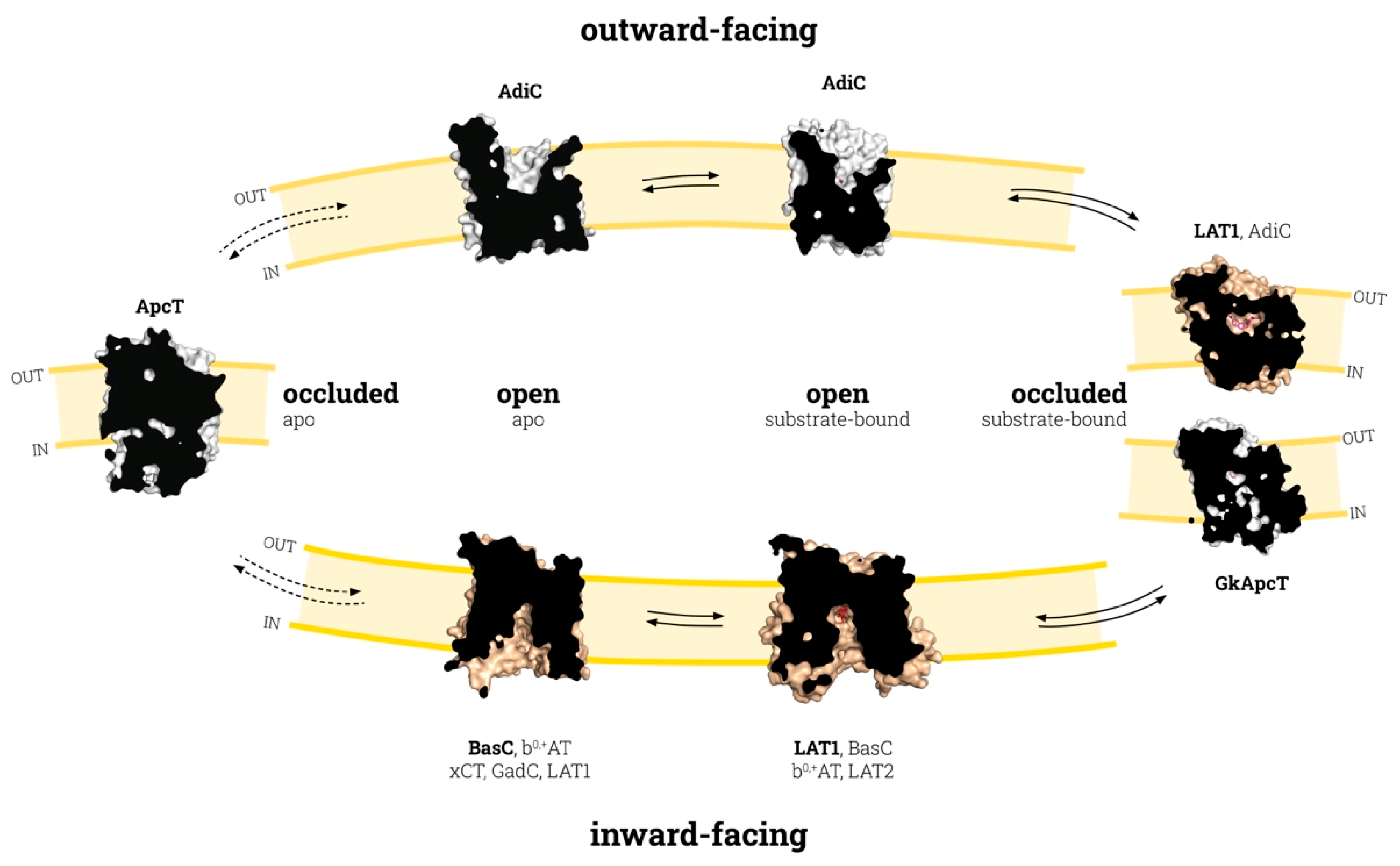
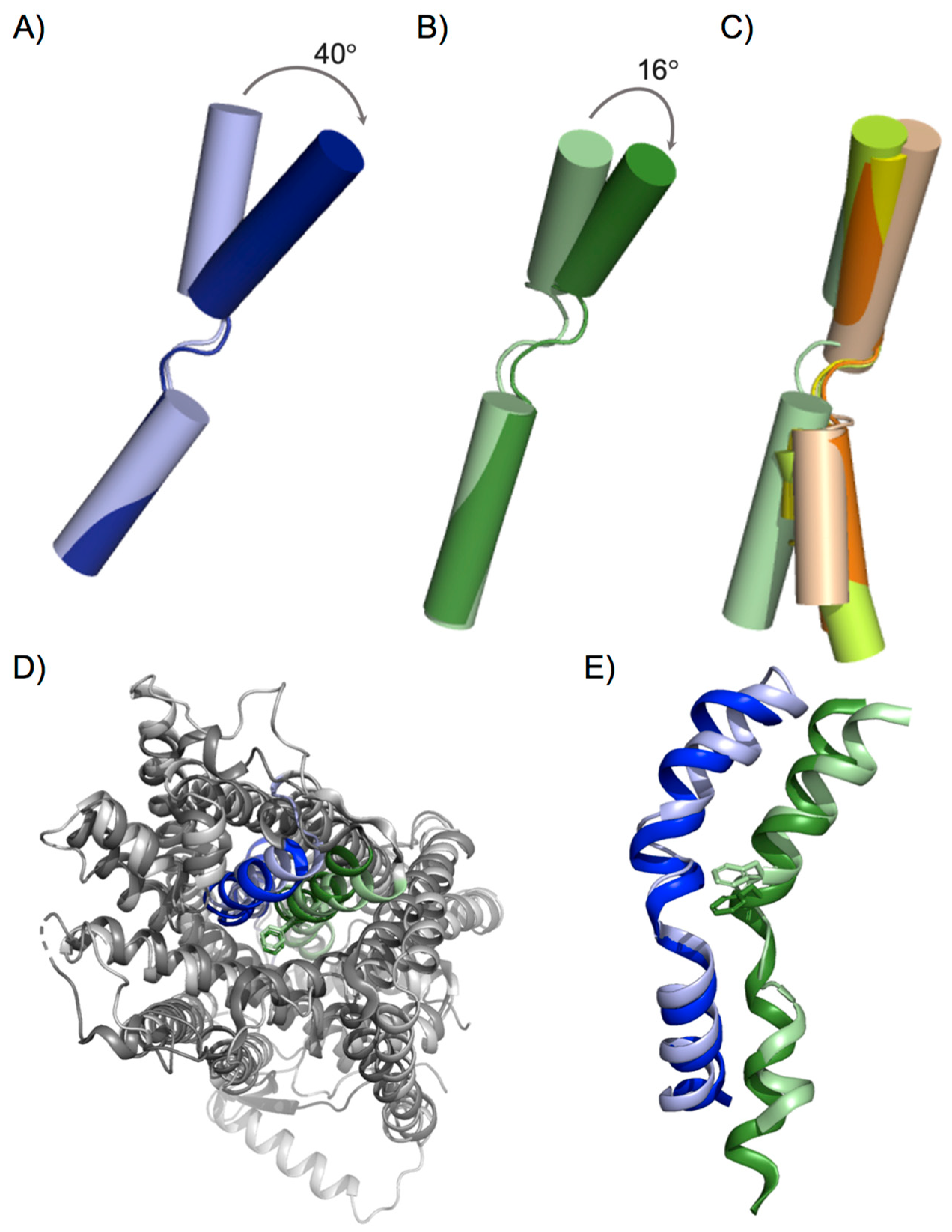
3. Conformation States in the Substrate Translocation Cycle of APC Transporters
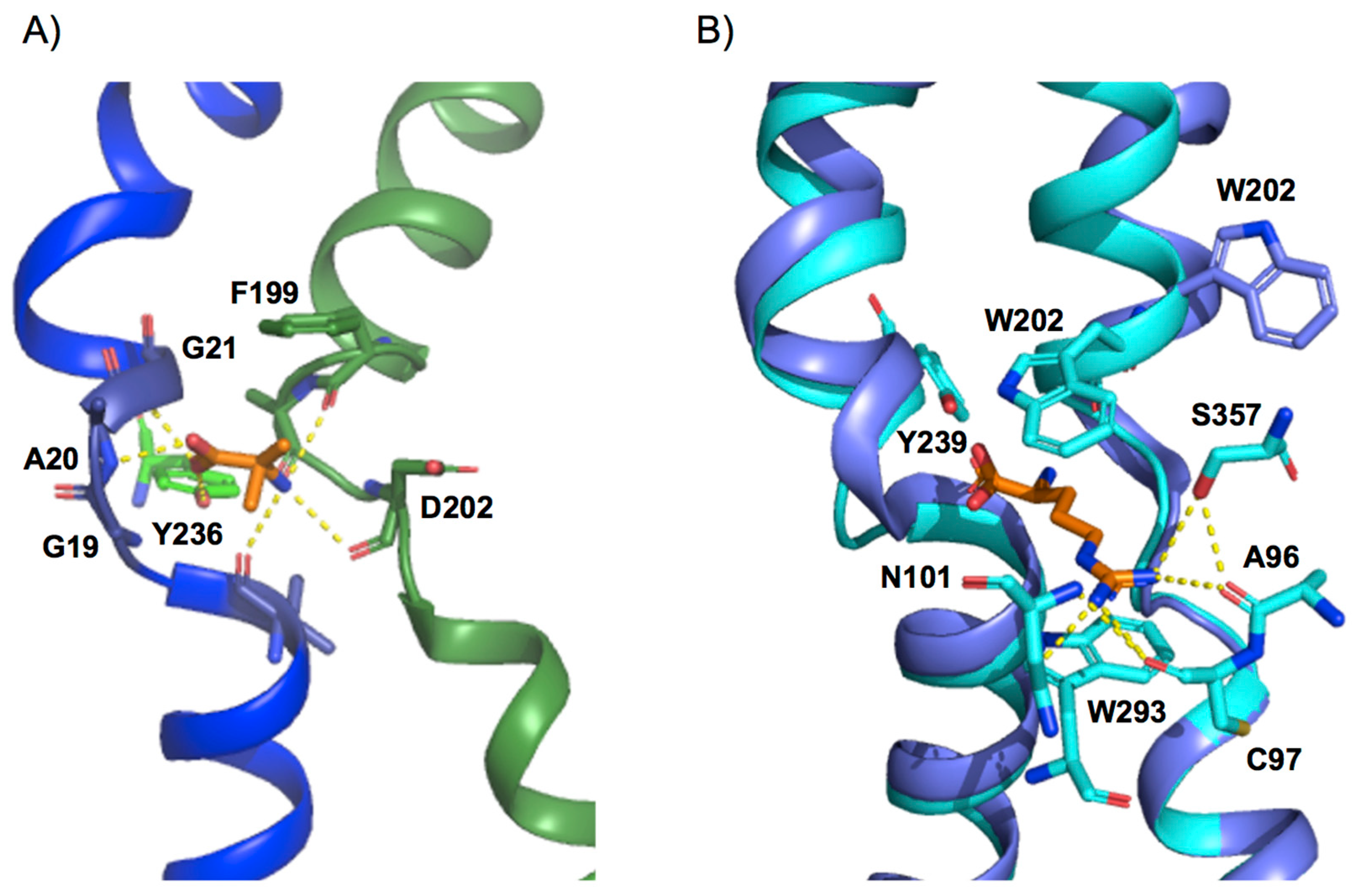
4. Design of the Substrate-Binding Site
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Almén, M.S.; Nordström, K.J.V.; Fredriksson, R.; Schiöth, H.B. Mapping the human membrane proteome: A majority of the human membrane proteins can be classified according to function and evolutionary origin. BMC Biol. 2009, 7, 50. [Google Scholar] [CrossRef]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 6220. [Google Scholar] [CrossRef]
- Overington, J.P.; Al-Lazikani, B.; Hopkins, A.L. How many drug targets are there? Nat. Rev. Drug Discov. 2006, 5, 993–996. [Google Scholar] [CrossRef] [PubMed]
- Biggin, P.C.; Aldeghi, M.; Bodkin, M.J.; Heifetz, A. Beyond membrane protein structure: Drug discovery, dynamics and difficulties. Adv. Exp. Med. Biol. 2016, 922, 161–181. [Google Scholar] [CrossRef]
- Yin, H.; Flynn, A.D. Drugging Membrane Protein Interactions. Annu. Rev. Biomed. Eng. 2016, 18, 51–76. [Google Scholar] [CrossRef] [PubMed]
- Bröer, S.; Bröer, A. Amino acid homeostasis and signalling in mammalian cells and organisms. Biochem. J. 2017, 474, 1935–1963. [Google Scholar] [CrossRef]
- Fotiadis, D.; Kanai, Y.; Palacín, M. The SLC3 and SLC7 families of amino acid transporters. Mol. Asp. Med. 2013, 34, 139–158. [Google Scholar] [CrossRef]
- Vastermark, A.; Wollwage, S.; Houle, M.E.; Rio, R.; Saier, M.H. Expansion of the APC superfamily of secondary carriers. Proteins Struct. Funct. Bioinform. 2014, 82, 2797–2811. [Google Scholar] [CrossRef]
- Kandasamy, P.; Gyimesi, G.; Kanai, Y.; Hediger, M.A. Amino acid transporters revisited: New views in health and disease. Trends Biochem Sci. 2018, 43, 752–789. [Google Scholar] [CrossRef]
- Feliubadaló, L.; Font, M.; Purroy, J.; Rousaud, F.; Estivill, X.; Nunes, V.; Golomb, E.; Centola, M.; Aksentijevich, I.; Kreiss, Y.; et al. Non-type I cystinuria caused by mutations in SLC7A9, encoding a subunit (b0,+AT) of rBAT. Nat. Genet. 1999, 23, 52–57. [Google Scholar] [CrossRef]
- Calonge, M.J.; Gasparini, P.; Chillarón, J.; Chillón, M.; Gallucci, M.; Rousaud, F.; Zelante, L.; Testar, X.; Dallapiccola, B.; Di Silverio, F.; et al. Cystinuria caused by mutations in rBAT, a gene involved in the transport of cystine. Nat. Genet. 1994, 6, 420–425. [Google Scholar] [CrossRef]
- Torrents, D.; Mykkänen, J.; Pineda, M.; Feliubadaló, L.; Estévez, R.; De Rafael, C.; Sanjurjo, P.; Zorzano, A.; Nunes, V.; Huoponen, K.; et al. Identification of SLC7A7, encoding y+LAT-1, as the lysinuric protein intolerance gene. Nat. Genet. 1999, 21, 293–296. [Google Scholar] [CrossRef]
- Espino-Guarch, M.; Font-Llitjós, M.; Murillo-Cuesta, S.; Errasti-Murugarren, E.; Celaya, A.M.; Girotto, G.; Vuckovic, D.; Mezzavilla, M.; Vilches, C.; Bodoy, S.; et al. Mutations in L-type amino acid transporter-2 support SLC7A8 as a novel gene involved in age-related hearing loss. eLife 2018, 7, e31511. [Google Scholar] [CrossRef]
- Knöpfel, E.B.; Vilches, C.; Camargo, S.M.R.; Errasti-Murugarren, E.; Stäubli, A.; Mayayo, C.; Munier, F.L.; Miroshnikova, N.; Poncet, N.; Junza, A.; et al. Dysfunctional LAT2 amino acid transporter is associated with cataract in mouse and humans. Front. Physiol. 2019, 10, 688. [Google Scholar] [CrossRef] [PubMed]
- Tărlungeanu, D.C.; Deliu, E.; Dotter, C.P.; Kara, M.; Janiesch, P.C.; Scalise, M.; Galluccio, M.; Tesulov, M.; Morelli, E.; Sonmez, F.M.; et al. Impaired amino acid transport at the blood brain barrier is a cause of autism spectrum disorder. Cell 2016, 167, 1481–1494.e18. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Noverr, M.C.; Parsons, C.; Kaleeba, J.A.R.; Qin, Z. xCT, not just an amino-acid transporter: A multi-functional regulator of microbial infection and associated diseases. Front. Microbiol. 2015, 6, 120. [Google Scholar] [CrossRef]
- Sason, H.; Billard, J.M.; Smith, G.P.; Safory, H.; Neame, S.; Kaplan, E.; Rosenberg, D.; Zubedat, S.; Foltyn, V.N.; Christoffersen, C.T.; et al. Asc-1 Transporter regulation of synaptic activity via the tonic release of d-serine in the forebrain. Cereb. Cortex 2017, 27, 1573–1587. [Google Scholar] [CrossRef] [PubMed]
- Lo, M.; Ling, V.; Wang, Y.Z.; Gout, P.W. The xc− cystine/glutamate antiporter: A mediator of pancreatic cancer growth with a role in drug resistance. Br. J. Cancer 2008, 99, 464–472. [Google Scholar] [CrossRef]
- Fan, X.; Ross, D.D.; Arakawa, H.; Ganapathy, V.; Tamai, I.; Nakanishi, T. Impact of system L amino acid transporter 1 (LAT1) on proliferation of human ovarian cancer cells: A possible target for combination therapy with anti-proliferative aminopeptidase inhibitors. Biochem. Pharmacol. 2010, 80, 811–818. [Google Scholar] [CrossRef]
- Yamashita, A.; Singh, S.K.; Kawate, T.; Jin, Y.; Gouaux, E. Crystal structure of a bacterial homologue of Na+/Cl−-dependent neurotransmitter transporters. Nature 2005, 437, 215–223. [Google Scholar] [CrossRef]
- Forrest, L.R.; Rudnick, G. The rocking bundle: A mechanism for ion-coupled solute flux by symmetrical transporters. Physiology 2009, 24, 377–386. [Google Scholar] [CrossRef]
- Kazmier, K.; Sharma, S.; Islam, S.M.; Roux, B.; Mchaourab, H.S.; Wright, E.M.; Geffen, D. Conformational cycle and ion-coupling mechanism of the Na+/hydantoin transporter Mhp1. Proc. Natl. Acad. Sci. USA 2014, 111, 14752–14757. [Google Scholar] [CrossRef]
- Kowalczyk, L.; Ratera, M.; Paladino, A.; Bartoccioni, P.; Errasti-Murugarren, E.; Valencia, E.; Portella, G.; Bial, S.; Zorzano, A.; Fita, I.; et al. Molecular basis of substrate-induced permeation by an amino acid antiporter. Proc. Natl. Acad. Sci. USA 2011, 108, 3935–3940. [Google Scholar] [CrossRef] [PubMed]
- Errasti-Murugarren, E.; Fort, J.; Bartoccioni, P.; Díaz, L.; Pardon, E.; Carpena, X.; Espino-Guarch, M.; Zorzano, A.; Ziegler, C.; Steyaert, J.; et al. L amino acid transporter structure and molecular bases for the asymmetry of substrate interaction. Nat. Commun. 2019, 10, 1807. [Google Scholar] [CrossRef] [PubMed]
- Jungnickel, K.E.J.; Parker, J.L.; Newstead, S. Structural basis for amino acid transport by the CAT family of SLC7 transporters. Nat. Commun. 2018, 9, 550. [Google Scholar] [CrossRef]
- Lee, Y.; Wiriyasermkul, P.; Jin, C.; Quan, L.; Ohgaki, R.; Okuda, S.; Kusakizako, T.; Nishizawa, T.; Oda, K.; Ishitani, R.; et al. Cryo-EM structure of the human L-type amino acid transporter 1 in complex with glycoprotein CD98hc. Nat. Struct. Mol. Biol. 2019, 26, 510–517. [Google Scholar] [CrossRef]
- Yan, R.; Zhao, X.; Lei, J.; Zhou, Q. Structure of the human LAT1–4F2hc heteromeric amino acid transporter complex. Nature 2019, 568, 127–130. [Google Scholar] [CrossRef]
- Perez, C.; Koshy, C.; Ressl, S.; Nicklisch, S.; Krämer, R.; Ziegler, C. Substrate specificity and ion coupling in the Na+/betaine symporter BetP. EMBO J. 2011, 30, 1221–1229. [Google Scholar] [CrossRef] [PubMed]
- Shaffer, P.L.; Goehring, A.; Shankaranarayanan, A.; Gouaux, E. Structure and mechanism of a Na+-independent amino acid transporter. Science 2009, 325, 1010–1014. [Google Scholar] [CrossRef]
- Gao, X.; Zhou, L.; Jiao, X.; Lu, F.; Yan, C.; Zeng, X.; Wang, J.; Shi, Y. Mechanism of substrate recognition and transport by an amino acid antiporter. Nature 2010, 463, 828–832. [Google Scholar] [CrossRef]
- Fang, Y.; Jayaram, H.; Shane, T.; Kolmakova-Partensky, L.; Wu, F.; Williams, C.; Xiong, Y.; Miller, C. Structure of a prokaryotic virtual proton pump at 3.2 Å resolution. Nature 2009, 460, 1040–1043. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Lu, F.; Zhou, L.; Dang, S.; Sun, L.; Li, X.; Wang, J.; Shi, Y. Structure and mechanism of an amino acid antiporter. Science 2009, 324, 1565–1568. [Google Scholar] [CrossRef]
- Ma, D.; Lu, P.; Yan, C.; Fan, C.; Yin, P.; Wang, J.; Shi, Y. Structure and mechanism of a glutamate-GABA antiporter. Nature 2012, 483, 632–636. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Ecker, G.F. Insights into the structure, function, and ligand discovery of the large neutral amino acid transporter 1, LAT1. Int. J. Mol. Sci. 2018, 19, 1278. [Google Scholar] [CrossRef] [PubMed]
- Geier, E.G.; Schlessinger, A.; Fan, H.; Gable, J.E.; Irwin, J.J.; Sali, A.; Giacomini, K.M. Structure-based ligand discovery for the Large-neutral Amino Acid Transporter 1, LAT-1. Proc. Natl. Acad. Sci. USA 2013, 110, 5480–5485. [Google Scholar] [CrossRef]
- Rosell, A.; Meury, M.; Alvarez-Marimon, E.; Costa, M.; Perez-Cano, L.; Zorzano, A.; Fernandez-Recio, J.; Palacin, M.; Fotiadis, D. Structural bases for the interaction and stabilization of the human amino acid transporter LAT2 with its ancillary protein 4F2hc. Proc. Natl. Acad. Sci. USA 2014, 111, 2966–2971. [Google Scholar] [CrossRef]
- Torrecillas, I.R.; Conde-Ceide, S.; De Lucas, A.I.; Garcĺa Molina, A.; Trabanco, A.A.; Lavreysen, H.; Pardo, L.; Tresadern, G. Inhibition of the alanine-serine-cysteine-1 transporter by BMS-466442. ACS Chem. Neurosci. 2019, 10, 2510–2517. [Google Scholar] [CrossRef]
- Singh, N.; Scalise, M.; Galluccio, M.; Wieder, M.; Seidel, T.; Langer, T.; Indiveri, C.; Ecker, G.F. Discovery of potent inhibitors for the large neutral amino acid transporter 1 (LAT1) by structure-based methods. Int. J. Mol. Sci. 2019, 20, 27. [Google Scholar] [CrossRef]
- Yan, R.; Li, Y.; Müller, J.; Zhang, Y.; Singer, S.; Xia, L.; Zhong, X.; Gertsch, J.; Altmann, K.H.; Zhou, Q. Mechanism of substrate transport and inhibition of the human LAT1-4F2hc amino acid transporter. Cell Discov. 2021, 7, 16. [Google Scholar] [CrossRef] [PubMed]
- Yan, R.; Li, Y.; Shi, Y.; Zhou, J.; Lei, J.; Huang, J.; Zhou, Q. Cryo-EM structure of the human heteromeric amino acid transporter b0,+AT-rBAT. Sci. Adv. 2020, 6, eaay6379. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Grund, T.N.; Welsch, S.; Mills, D.J.; Michel, M.; Safarian, S.; Michel, H. Structural basis for amino acid exchange by a human heteromeric amino acid transporter. Proc. Natl. Acad. Sci. USA 2020, 117, 21281–21287. [Google Scholar] [CrossRef]
- Yan, R.; Zhou, J.; Li, Y.; Lei, J.; Zhou, Q. Structural insight into the substrate recognition and transport mechanism of the human LAT2–4F2hc complex. Cell Discov. 2020, 6, 82. [Google Scholar] [CrossRef]
- Ilgü, H.; Jeckelmann, J.-M.; Gapsys, V.; Ucurum, Z.; de Groot, B.L.; Fotiadis, D. Insights into the molecular basis for substrate binding and specificity of the wild-type L-arginine/agmatine antiporter AdiC. Proc. Natl. Acad. Sci. USA 2016, 113, 10358–10363. [Google Scholar] [CrossRef]
- Sharma, M.; Rohithaswa, A.C. Atomic-level characterization of conformational transition and substrate binding of xCT transporter. bioRxiv 2018, 389643. [Google Scholar] [CrossRef]
- Lee, Y.; Wiriyasermkul, P.; Moriyama, S.; Mills, D.J.; Kühlbrandt, W.; Nagamori, S. Ca 2+-mediated higher-order assembly of b0,+ AT-rBAT is a key step for system b0,+ biogenesis and cystinuria. bioRxiv 2021. [Google Scholar] [CrossRef]
- Oda, K.; Lee, Y.; Wiriyasermkul, P.; Tanaka, Y.; Takemoto, M.; Yamashita, K.; Nagamori, S.; Nishizawa, T.; Nureki, O. Consensus mutagenesis approach improves the thermal stability of system xc− transporter, xCT, and enables cryo-EM analyses. Protein Sci. 2020, 29, 2398–2407. [Google Scholar] [CrossRef]
- Jardetzky, O. Simple allosteric model for membrane pumps. Nature 1966, 211, 969–970. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, M. Transport viewed as a catalytic process. Biochimie 2007, 89, 1042–1048. [Google Scholar] [CrossRef] [PubMed]
- Drew, D.; Boudker, O. Shared Molecular Mechanisms of Membrane Transporters. Annu. Rev. Biochem. 2016, 85, 543–572. [Google Scholar] [CrossRef] [PubMed]
- Stolzenberg, S.; Li, Z.; Quick, M.; Malinauskaite, L.; Nissen, P.; Weinstein, H.; Javitch, J.A.; Shi, L. The role of transmembrane segment 5 (TM5) in Na2 release and the conformational transition of neurotransmitter:sodium symporters toward the inward-open state. J. Biol. Chem. 2017, 292, 7372–7384. [Google Scholar] [CrossRef]
- Tsai, M.-F.; Fang, Y.; Miller, C. Sided functions of an arginine-agmatine antiporter oriented in liposomes. Biochemistry 2012, 51, 1577–1585. [Google Scholar] [CrossRef]
- Tavoulari, S.; Margheritis, E.; Nagarajan, A.; DeWitt, D.C.; Zhang, Y.-W.; Rosado, E.; Ravera, S.; Rhoades, E.; Forrest, L.R.; Rudnick, G. Two Na+ sites control conformational change in a neurotransmitter transporter homolog. J. Biol. Chem. 2016, 291, 1456–1471. [Google Scholar] [CrossRef] [PubMed]
- Loo, D.D.F.; Jiang, X.; Gorraitz, E.; Hirayama, B.A.; Wright, E.M. Functional identification and characterization of sodium binding sites in Na symporters. Proc. Natl. Acad. Sci. USA 2013, 110, E4557–E4566. [Google Scholar] [CrossRef]
- Hinz, K.M.; Meyer, K.; Kinne, A.; Schülein, R.; Köhrle, J.; Krause, G. Structural insights into thyroid hormone transport mechanisms of the L-type amino acid transporter 2. Mol. Endocrinol. 2015, 29, 933–942. [Google Scholar] [CrossRef]
- Nagamori, S.; Wiriyasermkul, P.; Okuda, S.; Kojima, N.; Hari, Y.; Kiyonaka, S.; Mori, Y.; Tominaga, H.; Ohgaki, R.; Kanai, Y. Structure-activity relations of leucine derivatives reveal critical moieties for cellular uptake and activation of mTORC1-mediated signaling. Amino Acids 2016, 48, 1045–1058. [Google Scholar] [CrossRef] [PubMed]
- Zur, A.A.; Chien, H.-C.; Augustyn, E.; Flint, A.; Heeren, N.; Finke, K.; Hernandez, C.; Hansen, L.; Miller, S.; Lin, L.; et al. LAT1 activity of carboxylic acid bioisosteres: Evaluation of hydroxamic acids as substrates. Bioorg. Med. Chem. Lett. 2016, 26, 5000–5006. [Google Scholar] [CrossRef] [PubMed]
- Bartoccioni, P.; Fort, J.; Zorzano, A.; Errasti-Murugarren, E.; Palacín, M. Functional characterization of the alanine-serine-cysteine exchanger of Carnobacterium sp AT7. J. Gen. Physiol. 2019, 151, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Meier, C. Activation of system L heterodimeric amino acid exchangers by intracellular substrates. EMBO J. 2002, 21, 580–589. [Google Scholar] [CrossRef]
- Malinauskaite, L.; Quick, M.; Reinhard, L.; Lyons, J.A.; Yano, H.; Javitch, J.A.; Nissen, P. A mechanism for intracellular release of Na+ by neurotransmitter/sodium symporters. Nat. Struct. Mol. Biol. 2014, 21, 1006–1012. [Google Scholar] [CrossRef]
- Khafizov, K.; Perez, C.; Koshy, C.; Quick, M.; Fendler, K.; Ziegler, C.; Forrest, L.R. Investigation of the sodium-binding sites in the sodium-coupled betaine transporter BetP. Proc. Natl. Acad. Sci. USA 2012, 109, E3035–E3044. [Google Scholar] [CrossRef]
- Zhang, Z.; Albers, T.; Fiumera, H.L.; Gameiro, A.; Grewer, C. A conserved Na+ binding site of the sodium-coupled neutral amino acid transporter 2 (SNAT2). J. Biol. Chem. 2009, 284, 25314–25323. [Google Scholar] [CrossRef]
- Bartoccioni, P.; Del Rio, C.; Ratera, M.; Kowalczyk, L.; Baldwin, J.M.; Zorzano, A.; Quick, M.; Baldwin, S.A.; Vázquez-Ibar, J.L.; Palacín, M. Role of transmembrane domain 8 in substrate selectivity and translocation of SteT, a member of the L-Amino acid Transporter (LAT) family. J. Biol. Chem. 2010, 285, 28764–28776. [Google Scholar] [CrossRef]
- Font, M.; Feliubadaló, L.; Estivill, X.; Nunes, V.; Golomb, E.; Kreiss, Y.; Pras, E.; Bisceglia, L.; d’Adamo, A.P.; Zelante, L.; et al. Functional analysis of mutations in SLC7A9, and genotype–phenotype correlation in non-Type I cystinuria. Hum. Mol. Genet. 2001, 10, 305–316. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rotoli, B.M.; Barilli, A.; Ingoglia, F.; Visigalli, R.; Bianchi, M.G.; Ferrari, F.; Martinelli, D.; Dionisi-Vici, C.; Dall’Asta, V. Analysis of LPI-causing mutations on y+LAT1 function and localization. Orphanet J. Rare Dis. 2019, 14, 63. [Google Scholar] [CrossRef] [PubMed]
- Sperandeo, M.P.; Andria, G.; Sebastio, G. Lysinuric protein intolerance: Update and extended mutation analysis of the SLC7A7 gene. Hum. Mutat. 2008, 29, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Font-Llitjós, M.; Jiménez-Vidal, M.; Bisceglia, L.; Di Perna, M.; De Sanctis, L.; Rousaud, F.; Zelante, L.; Palacín, M.; Nunes, V. New insights into cystinuria: 40 New mutations, genotype-phenotype correlation, and digenic inheritance causing partial phenotype. J. Med. Genet. 2005, 42, 58–68. [Google Scholar] [CrossRef]
- Closs, E.I.; Boissel, J.P.; Habermeier, A.; Rotmann, A. Structure and function of cationic amino acid transporters (CATs). J. Membr. Biol. 2006, 213, 67–77. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Transporter | Conformational State | Structural Information | Ref. | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Abbr. & Class | Name and Spp. | Facing | Opening | Substrate | Amph. | Å | PDB | Notes | |
| AdiC TC# 2.A.3.2.5 | Arginine:agmatine antiporter (E. coli) | Outward | Open | - | NG | 3.61 | 3LRB | [32] | |
| - | NG | 4.00 | 3LRC | [32] | |||||
| - | DM | 3.20 | 3NCY | (a) | [31] | ||||
| - | NG | 2.21 | 5J4I | N101A | [43] | ||||
| L-Arg | Cymal-6 | 3.00 | 3OB6 | [23] | |||||
| Agmatine | NG | 2.59 | 5J4N | [43] | |||||
| Occluded | L-Arg | NG | 3.00 | 3L1L | N22A | [30] | |||
| ApcT TC# 2.A.3.6.3 | Proton coupled amino acid transporter (M. jannaschii) | Inward | Occluded | OG | 2.32 | 3GIA | [29] | ||
| OG | 2.59 | 3GI8 | K158A (a) | [29] | |||||
| OG | 2.48 | 3GI9 | (a) | [29] | |||||
| GadC TC# 2.A.3.7.3 | Glutamate:GABA antiporter (E. coli) | Inward | Open | NG + LDAO | 3.10 | 4DJK | [33] | ||
| NG + LDAO | 3.19 | 4DJI | [33] | ||||||
| GkApcT TC# 2.A.3.3.n | Proton-coupled amino acid transporter (G. kaustophilus) | Inward | Occluded | L-Arg | DDM | 3.13 | 6F34 | (b) | [25] |
| L-Ala | DDM | 2.86 | 5OQT | (b) | [25] | ||||
| BasC TC# 2.A.3.8.n | Ala-Ser-Cys antiporter (Carnobacterium sp. AT7) | Inward | Open | DM | 2.92 | 6F2G | (c) | [24] | |
| 2-AIB * | DM | 3.40 | 6F2W | (c) | [24] | ||||
| LAT1 SLC7A5 TC# 2.A.3.8.25 | L-type amino acid transporter 1 (H. sapiens) | Inward | Open | BCH * | Digitonin | 3.50 | 6IRT | A36E | [27] |
| GDN | 3.30 | 6IRS | A36E | [27] | |||||
| Digitonin | 3.31 | 6JMQ | (a) | [26] | |||||
| Outward | Occluded | JX-075 * | GDN | 2.90 | 7DSK | [39] | |||
| JX-078 * | GDN | 2.90 | 7DSL | [39] | |||||
| JX-119 * | GDN | 3.10 | 7DSN | [39] | |||||
| Diiodo-Tyr * | GDN | 3.40 | 7DSQ | [39] | |||||
| b0,+AT SLC7A9 TC# 2.A.3.8.19 | b0,+-type amino acid transporter 1 (H. sapiens and Ovis sp.) | Inward | Open | L-Arg | GDN | 2.30 | 6LI9 | [40] | |
| GDN | 2.70 | 6LID | [40] | ||||||
| Digitonin | 2.90 | 6YUP | [41] | ||||||
| Digitonin | 3.40 | 6YV1 | [41] | ||||||
| Nanodisc | unreleased | [45] | |||||||
| Nanodisc | unreleased | [45] | |||||||
| LAT2 SLC7A8 TC# 2.A.3.8.20 | L-type amino acid transporter 2 (H. sapiens) | Inward | Open | L-Trp | GDN | 2.90 | 7CMH | [42] | |
| L-Leu | GDN | 3.40 | 7CMI | [42] | |||||
| xCT SLC7A9 TC# 2.A.3.8.19 | Cystine:glutamate antiporter (H. sapiens) | Inward | Open | Digitonin | 6.20 | 7CCS | [46] | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nicolàs-Aragó, A.; Fort, J.; Palacín, M.; Errasti-Murugarren, E. Rush Hour of LATs towards Their Transport Cycle. Membranes 2021, 11, 602. https://doi.org/10.3390/membranes11080602
Nicolàs-Aragó A, Fort J, Palacín M, Errasti-Murugarren E. Rush Hour of LATs towards Their Transport Cycle. Membranes. 2021; 11(8):602. https://doi.org/10.3390/membranes11080602
Chicago/Turabian StyleNicolàs-Aragó, Adrià, Joana Fort, Manuel Palacín, and Ekaitz Errasti-Murugarren. 2021. "Rush Hour of LATs towards Their Transport Cycle" Membranes 11, no. 8: 602. https://doi.org/10.3390/membranes11080602
APA StyleNicolàs-Aragó, A., Fort, J., Palacín, M., & Errasti-Murugarren, E. (2021). Rush Hour of LATs towards Their Transport Cycle. Membranes, 11(8), 602. https://doi.org/10.3390/membranes11080602