Effect of Temperature-Dependent Bacterial Growth during Milk Protein Fractionation by Means of 0.1 µM Microfiltration on the Length of Possible Production Cycle Times


Abstract
1. Introduction
2. Materials and Methods
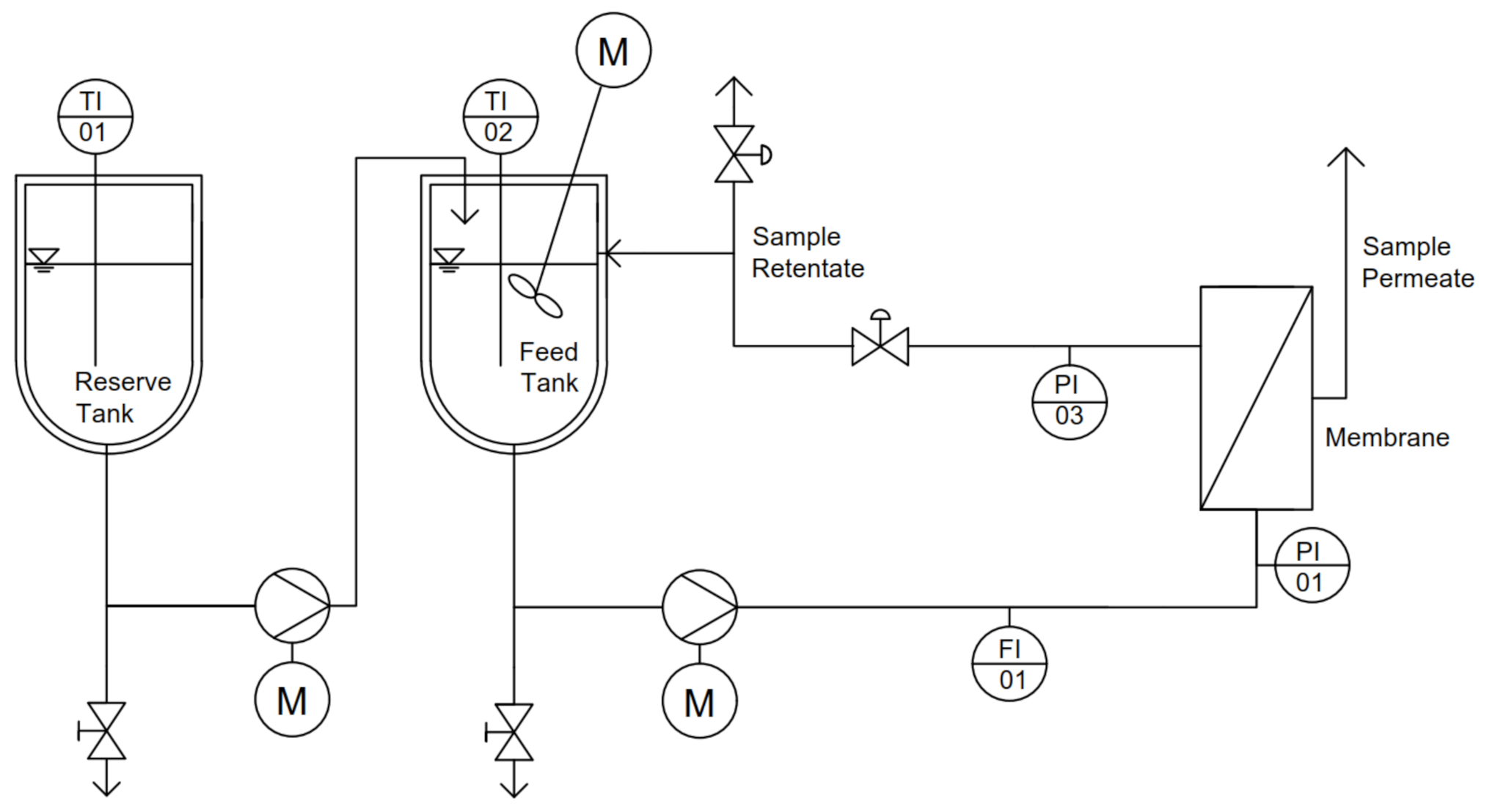
2.1. Skim Milk and Microfiltration Plant
2.2. Microfiltration of Skim Milk
2.3. Cleaning
2.4. Total Bacteria Count
2.5. Data Evaluation and Statistical Analysis
3. Results
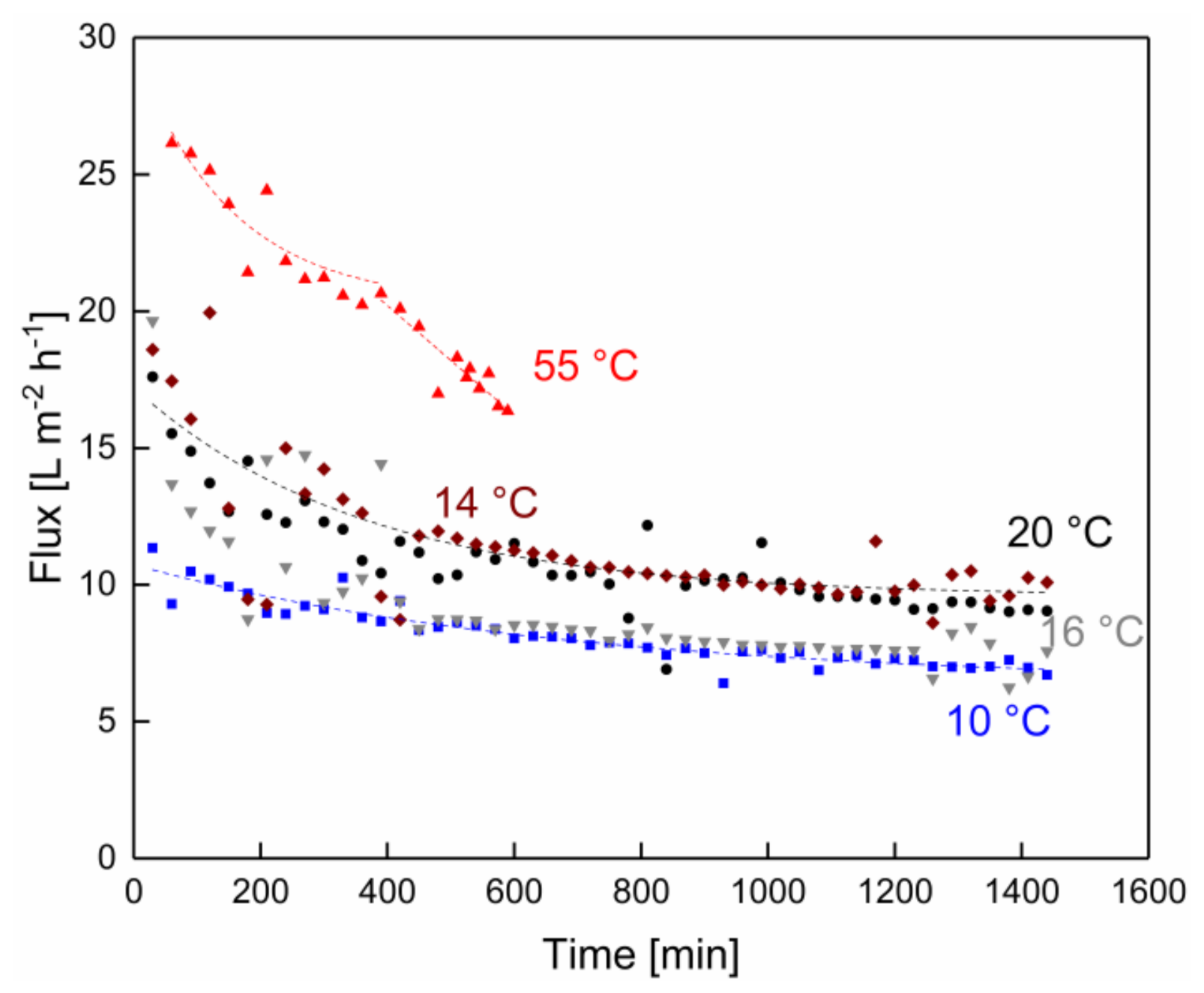
3.1. Impact of Temperature on the Filtration Performance over Time
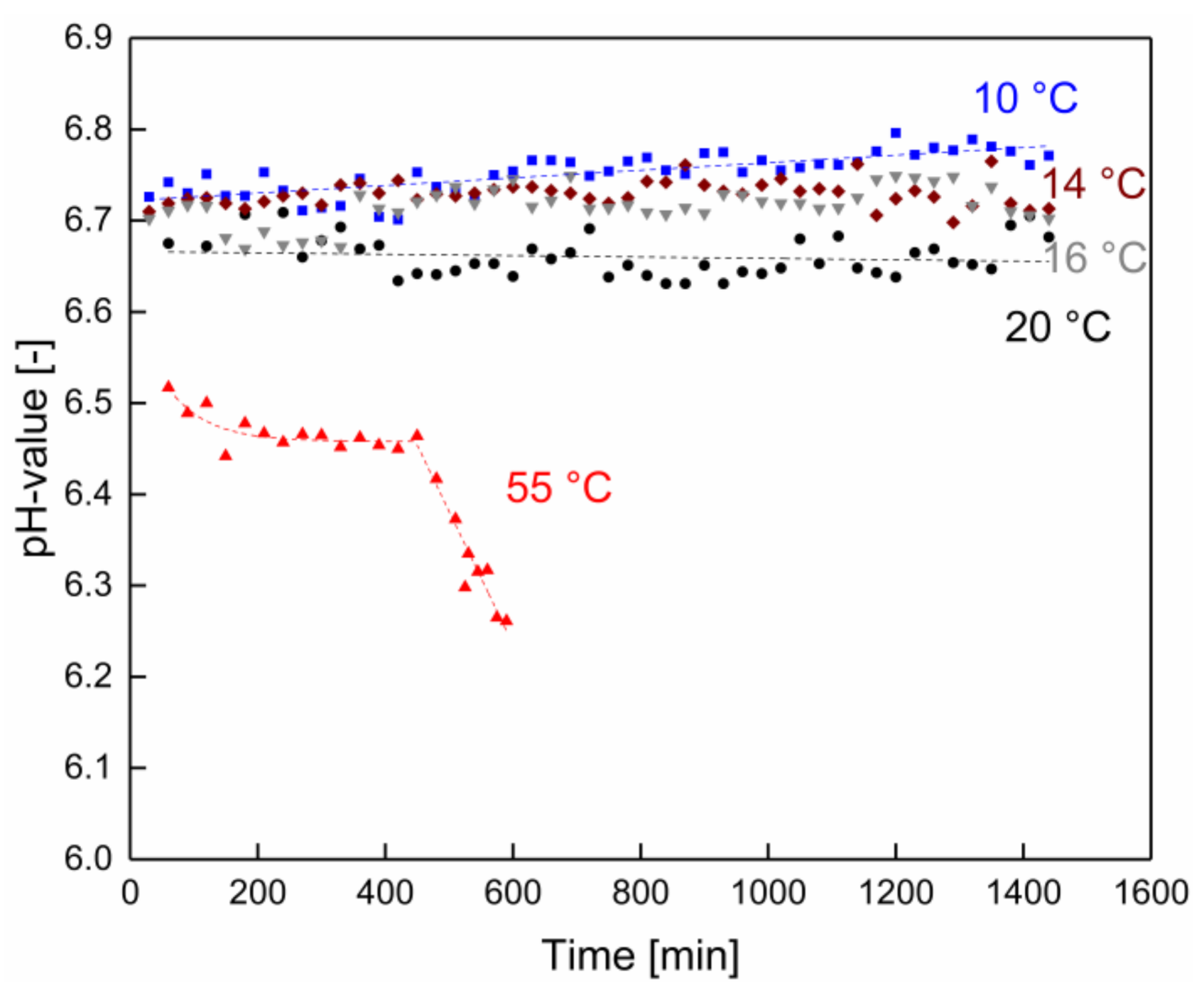
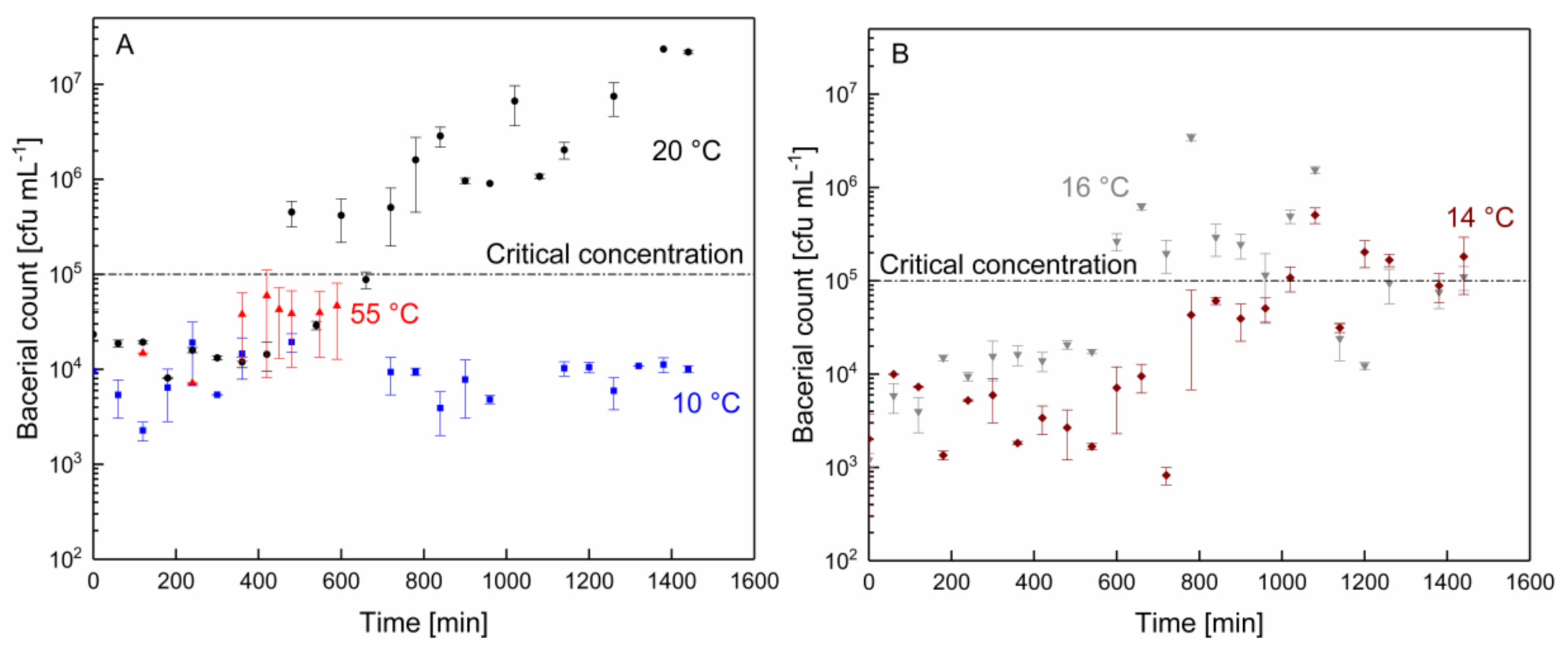
3.2. Changes in pH and Microbial Count as a Function of the Filtration Time
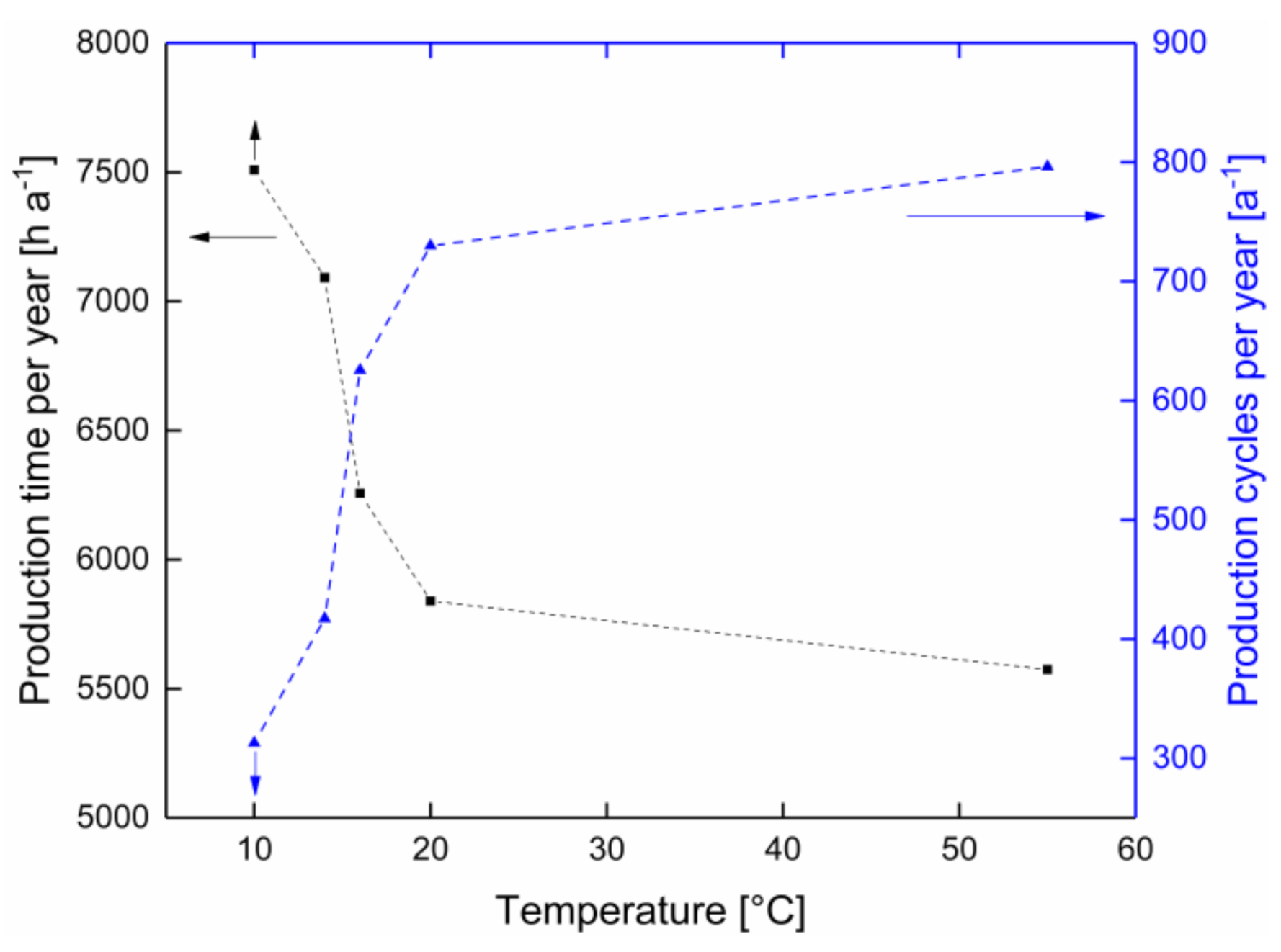
3.3. Calculation of the Possible Number of Production Cycles and Overall Annual Process Time
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hartinger, M.; Heidebrecht, H.-J.; Schiffer, S.; Dumpler, J.; Kulozik, U. Milk Protein Fractionation by Means of Spiral-Wound Microfiltration Membranes: Effect of the Pressure Adjustment Mode and Temperature on Flux and Protein Permeation. Foods 2019, 8, 180. [Google Scholar] [CrossRef]
- Zulewska, J.; Newbold, M.; Barbano, D.M. Efficiency of serum protein removal from skim milk with ceramic and polymeric membranes at 50 degrees C. J. Dairy Sci. 2009, 92, 1361–1377. [Google Scholar] [CrossRef] [PubMed]
- Rose, D. Relation Between Micellar and Serum Casein in Bovine Milk. J. Dairy Sci. 1968, 51, 1897–1902. [Google Scholar] [CrossRef]
- Chamberland, J.; Messier, T.; Dugat-Bony, E.; Lessard, M.-H.; Labrie, S.; Doyen, A.; Pouliot, Y. Influence of feed temperature to biofouling of ultrafiltration membrane during skim milk processing. Int. Dairy J. 2019, 93, 99–105. [Google Scholar] [CrossRef]
- Elwell, M.W.; Barbano, D.M. Use of Microfiltration to Improve Fluid Milk Quality. J. Dairy Sci. 2006, 89, E20–E30. [Google Scholar] [CrossRef]
- Gésan-Guiziou, G. Removal of bacteria, spores and somatic cells from milk by centrifugation and microfiltration techniques. Improving the Safety and Quality of Milk; Elsevier: Amsterdam, The Netherlands, 2010; pp. 349–372. ISBN 9781845694388. [Google Scholar]
- Schmidt, V.S.J.; Kaufmann, V.; Kulozik, U.; Scherer, S.; Wenning, M. Microbial biodiversity, quality and shelf life of microfiltered and pasteurized extended shelf life (ESL) milk from Germany, Austria and Switzerland. Int. J. Food Microbiol. 2012, 154, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, W.; Kiesner, C.; Clawin-Rädecker, I.; Martin, D.; Einhoff, K.; Lorenzen, P.C.; Meisel, H.; Hammer, P.; Suhren, G.; Teufel, P. Processing of extended shelf life milk using microfiltration. Int. J. Dairy Technol 2006, 59, 229–235. [Google Scholar] [CrossRef]
- Kaufmann, V.; Scherer, S.; Kulozik, U. Verfahren zur Verlängerung der Haltbarkeit von Konsummilch und ihre stofflichen Veränderungen: ESL-Milch. J. Verbr. Lebensm. 2010, 5, 59–64. [Google Scholar] [CrossRef]
- Lipnizki, F.; Boelsmand, J.; Madsen, R.F. Concepts of industrial-scale diafiltration systems. Desalination 2002, 144, 179–184. [Google Scholar] [CrossRef]
- Skrzypek, M.; Burger, M. Isoflux® ceramic membranes — Practical experiences in dairy industry. Desalination 2010, 250, 1095–1100. [Google Scholar] [CrossRef]
- Porcellato, D.; Aspholm, M.; Skeie, S.B.; Monshaugen, M.; Brendehaug, J.; Mellegård, H. Microbial diversity of consumption milk during processing and storage. Int. J. Food Microbiol. 2018, 266, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Burgess, S.A.; Lindsay, D.; Flint, S.H. Thermophilic bacilli and their importance in dairy processing. Int. J. Food Microbiol. 2010, 144, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.M.; Barach, J.T.; Speck, M.L. Heat Resistant Proteases Produced in Milk by Psychrotorophic Bacteria of Dairy Origin. J. Dairy Sci. 1975, 58, 828–834. [Google Scholar] [CrossRef]
- Deeth, H.C.; Khusniati, T.; Datta, N.; Wallace, R.B. Spoilage patterns of skim and whole milks. J. Dairy Res. 2002, 69, 227–241. [Google Scholar] [CrossRef]
- Kaufmann, V. Einflussfaktoren auf die Produktcharakteristika von Extended Shelf Life (ESL)-Milch. Ph.D. Thesis, Technische Universität München, München, Germany, 2012. [Google Scholar]
- Parkar, S.G.; Flint, S.H.; Palmer, J.S.; Brooks, J.D. Factors influencing attachment of thermophilic bacilli to stainless steel. J. Appl. Microbiol. 2001, 90, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Guidelines—EHEDG. Available online: https://www.ehedg.org/guidelines/ (accessed on 8 October 2020).
- Jarto, I.; Lucey, J.A.; Smith, K.E. Impact of processing temperature on production of milk protein permeate during microfiltration of skim or whole milk. Int. J. Dairy Technol. 2018, 71, 844–848. [Google Scholar] [CrossRef]
- Crowley, S.V.; Caldeo, V.; McCarthy, N.A.; Fenelon, M.A.; Kelly, A.L.; O’Mahony, J.A. Processing and protein-fractionation characteristics of different polymeric membranes during filtration of skim milk at refrigeration temperatures. Int. Dairy J. 2015, 48, 23–30. [Google Scholar] [CrossRef]
- Samarzija, D.; Zamberlin, S.; Pogacic, T. Psychrotrophic bacteria and their negative effects on milk and dairy products quality. Mljekarstvo 2012, 62, 77–95. [Google Scholar]
- Ternström, A.; Lindberg, A.M.; Molin, G. Classification of the spoilage flora of raw and pasteurized bovine milk, with special reference to Pseudomonas and Bacillus. J. Appl. Bacteriol. 1993, 75, 25–34. [Google Scholar] [CrossRef]
- Seale, B.; Bremer, P.; Flint, S.; Brooks, J.; Palmer, J. Overview of the Problems Resulting from Biofilm Contamination in the Dairy Industry. In Biofilms in the dairy industry; Knight, G., Brooks, J., Teh, K.H., Flint, S., Eds.; Wiley Blackwell: Chichester, UK; Hoboken, NJ, USA, 2015; ISBN 9781118876282. [Google Scholar]
- Bast, E. Mikrobiologische Methoden. Eine Einführung in grundlegende Arbeitstechniken, 3rd ed.; Springer Spektrum: Berlin, Germany, 2014; ISBN 978-3-8274-1813-5. [Google Scholar]
- Marx, M.; Sixt, A.; Hofsommer, J.; Wörthmann, M.; Kulozik, U. Manufacturing of demineralized whey concentrates with extended shelf life: Impact of the degree of demineralization on functional and microbial quality criteria. Food Bioprod. Process. 2019, 114, 1–11. [Google Scholar] [CrossRef]
- Rezaei, H.; Ashtiani, F.Z.; Fouladitajar, A. Effects of operating parameters on fouling mechanism and membrane flux in cross-flow microfiltration of whey. Desalination 2011, 274, 262–271. [Google Scholar] [CrossRef]
- Zulewska, J.; Barbano, D.M. Influence of casein on flux and passage of serum proteins during microfiltration using polymeric spiral-wound membranes at 50 °C. J. Dairy Sci. 2013, 96, 2048–2060. [Google Scholar] [CrossRef]
- Iametti, S.; de Gregori, B.; Vecchio, G.; Bonomi, F. Modifications occur at different structural levels during the heat denaturation of beta-lactoglobulin. Eur. J. Biochem. 1996, 237, 106–112. [Google Scholar] [CrossRef]
- Ng, K.S.Y.; Dunstan, D.E.; Martin, G.J.O. Influence of processing temperature on flux decline during skim milk ultrafiltration. Sep. Purif. Technol. 2018, 195, 322–331. [Google Scholar] [CrossRef]
- Steinhauer, T.; Kulozik, U.; Gebhardt, R. Structure of milk protein deposits formed by casein micelles and β-lactoglobulin during frontal microfiltration. J. Membr. Sci. 2014, 468, 126–132. [Google Scholar] [CrossRef]
- Chaplin, L.G.; Lyster, R.L.J. Effect of temperature on the pH of skim milk. J. Dairy Res. 1988, 55, 277–280. [Google Scholar] [CrossRef]
- Kühnl, W.; Piry, A.; Kaufmann, V.; Grein, T.; Ripperger, S.; Kulozik, U. Impact of colloidal interactions on the flux in cross-flow microfiltration of milk at different pH values: A surface energy approach. J. Membr. Sci. 2010, 352, 107–115. [Google Scholar] [CrossRef]
- Sadiq, F.A.; Flint, S.; Yuan, L.; Li, Y.; Liu, T.; He, G. Propensity for biofilm formation by aerobic mesophilic and thermophilic spore forming bacteria isolated from Chinese milk powders. Int. J. Food Microbiol. 2017, 262, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Bijl, E.; Svensson, B.; Hettinga, K. The Extracellular Protease AprX from Pseudomonas and its Spoilage Potential for UHT Milk: A Review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 834–852. [Google Scholar] [CrossRef]
- Kroll, S.; Klostermeyer, H. Heat inactivation of exogenous proteinases from Pseudomonas fluorescens. I. Possibility of inactivation in milk. Zeitschrift fur Lebensmittel-Untersuchung und -Forschung 1984, 179, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Flint, S.; Bennett, R.; Brooks, J.; Md Zain, S.N. Biofilm Contamination of Ultrafiltration and Reverse Osmosis Plants. In Biofilms in the Dairy Industry; Knight, G., Brooks, J., Teh, K.H., Flint, S., Eds.; Wiley Blackwell: Chichester, UK; Hoboken, NJ, USA, 2015; pp. 138–153. ISBN 9781118876282. [Google Scholar]
- Chamberland, J.; Lessard, M.-H.; Doyen, A.; Labrie, S.; Pouliot, Y. A sequencing approach targeting the 16S rRNA gene unravels the biofilm composition of spiral-wound membranes used in the dairy industry. Dairy Sci. Technol. 2017, 96, 827–843. [Google Scholar] [CrossRef]
- Vithanage, N.R.; Dissanayake, M.; Bolge, G.; Palombo, E.A.; Yeager, T.R.; Datta, N. Biodiversity of culturable psychrotrophic microbiota in raw milk attributable to refrigeration conditions, seasonality and their spoilage potential. Int. Dairy J. 2016, 57, 80–90. [Google Scholar] [CrossRef]
- Kieferle, I.; Meintrup, D.; Kulozik, U. Interrelations between consecutive process steps: Using the example of the displacement of dispersions subsequently to the filtration. J. Food Eng. 2019, 263, 155–164. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Production temperature (°C) | 10 | 14 | 16 | 20 | 55 |
| Possible production time (h) | >24 | 17 | 10 | 8 | 7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schiffer, S.; Kulozik, U. Effect of Temperature-Dependent Bacterial Growth during Milk Protein Fractionation by Means of 0.1 µM Microfiltration on the Length of Possible Production Cycle Times. Membranes 2020, 10, 326. https://doi.org/10.3390/membranes10110326
Schiffer S, Kulozik U. Effect of Temperature-Dependent Bacterial Growth during Milk Protein Fractionation by Means of 0.1 µM Microfiltration on the Length of Possible Production Cycle Times. Membranes. 2020; 10(11):326. https://doi.org/10.3390/membranes10110326
Chicago/Turabian StyleSchiffer, Simon, and Ulrich Kulozik. 2020. "Effect of Temperature-Dependent Bacterial Growth during Milk Protein Fractionation by Means of 0.1 µM Microfiltration on the Length of Possible Production Cycle Times" Membranes 10, no. 11: 326. https://doi.org/10.3390/membranes10110326
APA StyleSchiffer, S., & Kulozik, U. (2020). Effect of Temperature-Dependent Bacterial Growth during Milk Protein Fractionation by Means of 0.1 µM Microfiltration on the Length of Possible Production Cycle Times. Membranes, 10(11), 326. https://doi.org/10.3390/membranes10110326