
Resistance to Ticks and the Path to Anti-Tick and Transmission Blocking Vaccines



Abstract
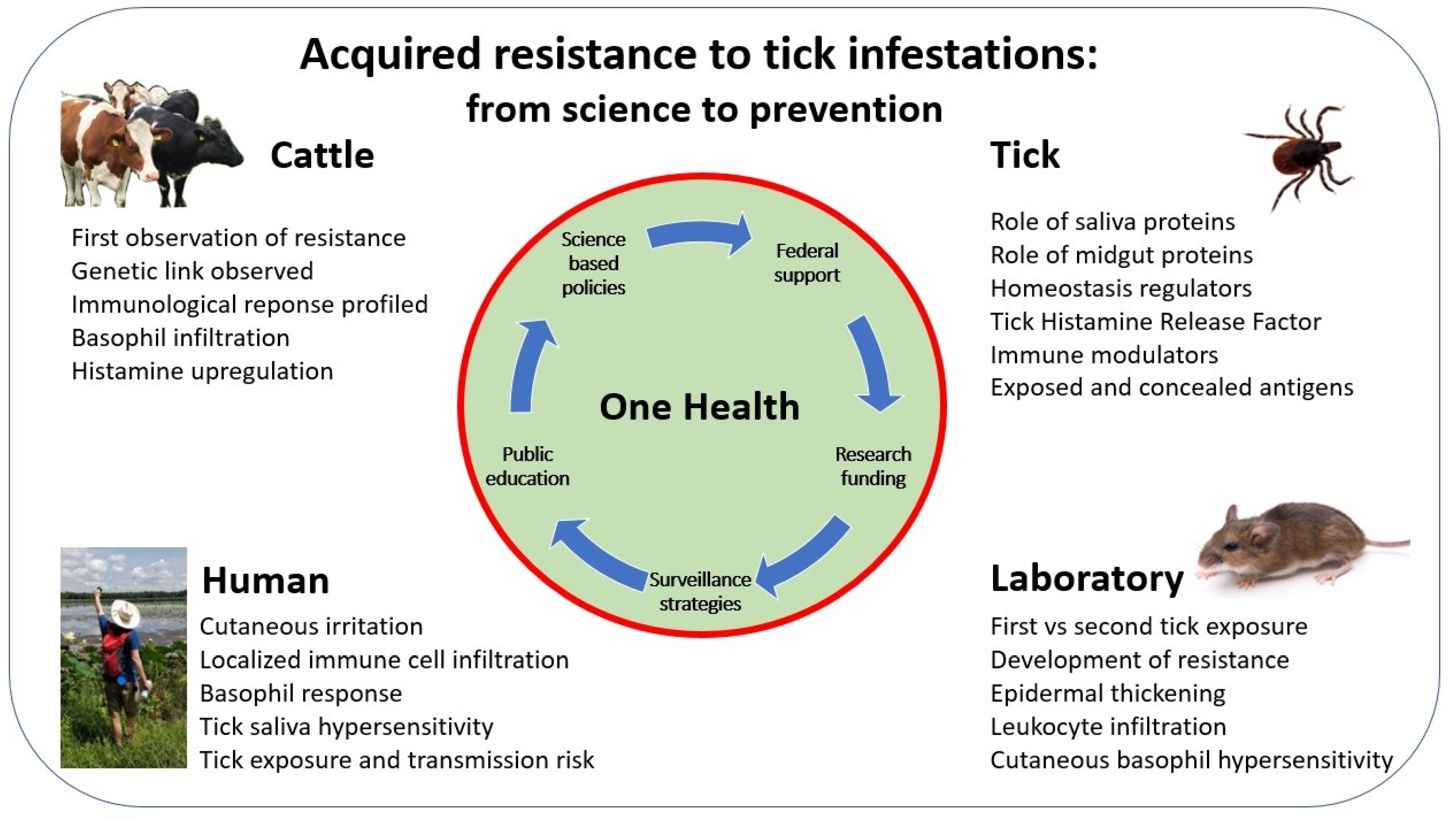
1. Introduction
- Acquired resistance to tick infestations, and the evolution of our understanding throughout decades of research.
- Common elements of host responses leading to acquired resistance.
- Vaccine development and the use of acquired resistance mechanisms, tick physiology, and tick microbiome as targets.
2. Acquired Resistance to Tick Infestations and the Evolution of Our Understanding
2.1. Bovine Resistance to Ticks—First Encounter
2.2. Laboratory Studies—Our Best Defense Is Knowledge
2.3. Human Hypersensitivity to Tick Bite—The Final Frontier
3. Common Elements of Host Responses Leading to Acquired Resistance
3.1. Basophil Response of Acquired Resistance
3.2. Direct Action of Histamine on Ticks
3.3. Tick Saliva: Many Questions Remain
4. Anti-Tick Vaccination Strategies
4.1. The Use of Acquired Resistance Mechanisms as a Guide for the Development of Vaccines
4.2. Host Antibodies Enter Tick Haemocoele: More Potential Vaccine Antigen Targets
4.3. Tick Tissue Antigens
4.4. Salivary Gland Derived Antigens
4.5. Novel Antigen Sources
5. Reservoir Targeted Vaccines
6. Concluding Thoughts
Author Contributions
Funding
Institutional Animal Care and Use Committee Statement
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Jongejan, F.; Uilenberg, G. The global importance of ticks. Parasitology. 2004, 129, S3–S14. [Google Scholar] [CrossRef] [PubMed]
- Dantas-Torres, F. Climate changes, biodiversity, ticks and tick-borne diseases: The butterfly effect. Int. J. Parasitol. Parasites Wildl. 2015, 4, 452–461. [Google Scholar] [CrossRef]
- Gray, J.S.; Dautel, H.; Estrada-Peña, A.; Kahl, O.; Lindgren, E. Effects of climate change on ticks and tick-borne diseases in Europe. Interdiscip. Perspect. Infect. Dis. 2009, 2009, 593232. [Google Scholar] [CrossRef]
- Ogden, N.H.; Lindsay, L.R. Effects of climate and climate change on vectors and vector-borne diseases: Ticks are different. Trends Parasitol. 2016, 32, 646–656. [Google Scholar] [CrossRef]
- Paules, C.I.; Marston, H.D.; Bloom, M.E.; Fauci, A.S. Tickborne diseases—Confronting a growing threat. N. Engl. J. Med. 2018, 379, 701–703. [Google Scholar] [CrossRef]
- Eisen, L. Control of ixodid ticks and prevention of tick-borne diseases in the United States: The prospect of a new Lyme disease vaccine and the continuing problem with tick exposure on residential properties. Ticks Tick Borne Dis. 2021, 12, 101649. [Google Scholar] [CrossRef]
- Abbas, R.Z.; Zaman, M.A.; Colwell, D.D.; Gilleard, J.; Iqbal, Z. Acaricide resistance in cattle ticks and approaches to its management: The state of play. Vet. Parasitol. 2014, 203, 6–20. [Google Scholar] [CrossRef] [PubMed]
- Jeschke, P. Status and outlook for acaricide and insecticide discovery. Pest Manag. Sci. 2021, 77, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Eisen, R.J.; Kugeler, K.J.; Eisen, L.; Beard, C.B.; Paddock, C.D. Tick-borne zoonoses in the United States: Persistent and emerging threats to human health. ILAR J. 2017, 58, 319–335. [Google Scholar] [CrossRef]
- Eisen, L.; Dolan, M.C. Evidence for personal protective measures to reduce human contact with blacklegged ticks and for environmentally based control methods to suppress host-seeking blacklegged ticks and reduce infection with Lyme disease spirochetes in tick vectors and rodent reservoirs. J. Med. Entomol. 2016, 53, 1063–1092. [Google Scholar] [CrossRef]
- Beard, C.B.; Visser, S.N.; Petersen, L.R. The need for a national strategy to address vector-borne disease threats in the United States. J. Med. Entomol. 2019, 56, 1199–1203. [Google Scholar] [CrossRef]
- Rochlin, I.; Ninivaggi, D.V.; Benach, J.L. Malaria and Lyme disease—The largest vector-borne US epidemics in the last 100 years: Success and failure of public health. BMC Public Health 2019, 19, 804. [Google Scholar] [CrossRef] [PubMed]
- Eisen, L. Stemming the rising tide of human-biting ticks and tickborne diseases, United States. Emerg. Infect. Dis. 2020, 26, 641–647. [Google Scholar] [CrossRef]
- Eisen, R.J.; Paddock, C.D. Tick and tickborne pathogen surveillance as a public health tool in the United States. J. Med. Entomol. 2020, 22, tjaa087. [Google Scholar] [CrossRef]
- Eisen, L.; Stafford, K.C. Barriers to effective tick management and tick-bite prevention in the United States (Acari: Ixodidae). J. Med. Entomol. 2020, 6, tjaa079. [Google Scholar] [CrossRef]
- Little, E.A.H.; Williams, S.C.; Stafford, K.C., 3rd; Linske, M.A.; Molaei, G. Evaluating the effectiveness of an integrated tick management approach on multiple pathogen infection in Ixodes scapularis questing nymphs and larvae parasitizing white-footed mice. Exp. Appl. Acarol. 2020, 80, 127–136. [Google Scholar] [CrossRef]
- Dantas-Torres, F.; Chomel, B.B.; Otranto, D. Ticks and tick-borne diseases: A One Health perspective. Trends Parasitol. 2012, 28, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Destoumieux-Garzón, D.; Mavingui, P.; Boetsch, G.; Boissier, J.; Darriet, F.; Duboz, P.; Fritsch, C.; Giraudoux, P.; Le Roux, F.; Morand, S.; et al. The One Health concept: 10 years old and a long road ahead. Front. Vet. Sci. 2018, 5, 14. [Google Scholar] [CrossRef]
- Willadsen, P. Anti-tick vaccines. Parasitology. 2004, 129, S367–S387. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Martín, V.; Manzano-Román, R.; Obolo-Mvoulouga, P.; Oleaga, A.; Pérez-Sánchez, R. Development of vaccines against Ornithodoros soft ticks: An update. Ticks Tick Borne Dis. 2015, 6, 211–220. [Google Scholar] [CrossRef]
- Rego, R.O.M.; Trentelman, J.J.A.; Anguita, J.; Nijhof, A.M.; Sprong, H.; Klempa, B.; Hajdusek, O.; Tomás-Cortázar, J.; Azagi, T.; Strnad, M.; et al. Counterattacking the tick bite: Towards a rational design of anti-tick vaccines targeting pathogen transmission. Parasit. Vectors 2019, 12, 229. [Google Scholar] [CrossRef] [PubMed]
- Ndawula, C., Jr.; Tabor, A.E. Cocktail anti-tick vaccines: The unforeseen constraints and approaches toward enhanced efficacies. Vaccines 2020, 8, 457. [Google Scholar] [CrossRef] [PubMed]
- Bhowmick, B.; Han, Q. Understanding tick biology and its implications in anti-tick and transmission blocking vaccines against tick-borne pathogens. Front. Vet. Sci. 2020, 7, 319. [Google Scholar] [CrossRef] [PubMed]
- Stafford, K.C., 3rd; Williams, S.C.; van Oosterwijk, J.G.; Linske, M.A.; Zatechka, S.; Richer, L.M.; Molaei, G.; Przybyszewski, C.; Wikel, S.K. Field evaluation of a novel oral reservoir-targeted vaccine against Borrelia burgdorferi utilizing an inactivated whole-cell bacterial antigen expression vehicle. Exp. Appl. Acarol. 2020, 80, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.C.; van Oosterwijk, J.G.; Linske, M.A.; Zatechka, S.; Richer, L.M.; Przybyszewski, C.; Wikel, S.K.; Stafford, K.C., III. Administration of an orally delivered substrate targeting a mammalian zoonotic pathogen reservoir population: Novel application and biomarker analysis. Vector Borne Zoonotic Dis. 2020, 20, 603–612. [Google Scholar] [CrossRef]
- Van Oosterwijk, J.G. Anti-tick and pathogen transmission blocking vaccines. Parasite Immunol. 2021, 43, e21831. [Google Scholar] [CrossRef]
- Bogovic, P.; Strle, F. Tick-borne encephalitis: A review of epidemiology, clinical characteristics, and management. World J. Clin. Cases. 2015, 3, 430–441. [Google Scholar] [CrossRef]
- Boulanger, N.; Wikel, S. Induced transient immune tolerance in ticks and vertebrate host: A keystone of tick-borne diseases? Front. Immunol. 2021, 12, 625993. [Google Scholar] [CrossRef]
- Neelakanta, G.; Sultana, H. Transmission-blocking vaccines: Focus on anti-vector vaccines against tick-borne diseases. Arch. Immunol. Ther. Exp. 2015, 63, 169–179. [Google Scholar] [CrossRef]
- De la Fuente, J.; Kopáček, P.; Lew-Tabor, A.; Maritz-Olivier, C. Strategies for new and improved vaccines against ticks and tick-borne diseases. Parasite Immunol. 2016, 38, 754–769. [Google Scholar] [CrossRef]
- De la Fuente, J.; Estrada-Peña, A.; Contreras, M. Modeling tick vaccines: A key tool to improve protection efficacy. Expert Rev. Vaccines 2020, 19, 217–225. [Google Scholar] [CrossRef]
- Almazan, C.; Tipacamu, G.A.; Rodriguez, S.; Mosqueda, J.; Perez de Leon, A. Immunological control of ticks and tick-borne diseases that impact cattle health and production. Front. Biosci. 2018, 23, 1535–1551. [Google Scholar] [CrossRef]
- Rodriguez-Vivas, R.I.; Jonsson, N.N.; Bhushan, C. Strategies for the control of Rhipicephalus microplus ticks in a world of conventional acaricide and macrocyclic lactone resistance. Parasitol. Res. 2018, 117, 3–29. [Google Scholar] [CrossRef] [PubMed]
- Mateos-Hernández, L.; Obregón, D.; Maye, J.; Borneres, J.; Versille, N.; de la Fuente, J.; Estrada-Peña, A.; Hodžić, A.; Šimo, L.; Cabezas-Cruz, A. Anti-tick microbiota vaccine impacts Ixodes ricinus performance during feeding. Vaccines 2020, 8, 702. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.G. Dynamics and impact of tick-borne diseases of cattle. Trop. Anim. Health Prod. 1997, 29, 1S–3S. [Google Scholar] [CrossRef]
- Heyman, P.; Cochez, C.; Hofhuis, A.; van der Giessen, J.; Sprong, H.; Porter, S.R.; Losson, B.; Saegerman, C.; Donoso-Mantke, O.; Niedrig, M.; et al. A clear and present danger: Tick-borne diseases in Europe. Expert Rev. Anti. Infect. Ther. 2010, 8, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.R. Eradication and control of livestock ticks: Biological, economic and social perspectives. Parasitology. 2011, 138, 945–959. [Google Scholar] [CrossRef]
- Tabor, A.E.; Ali, A.; Rehman, G.; Rocha Garcia, G.; Zangirolamo, A.F.; Malardo, T.; Jonsson, N.N. Cattle Tick Rhipicephalus microplus-Host Interface: A Review of Resistant and Susceptible Host Responses. Front. Cell. Infect. Microbiol. 2017, 7, 506. [Google Scholar] [CrossRef]
- Narladkar, B.W. Projected economic losses due to vector and vector-borne parasitic diseases in livestock of India and its significance in implementing the concept of integrated practices for vector management. Vet. World. 2018, 11, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Lew-Tabor, A.E.; Rodriguez Valle, M. A review of reverse vaccinology approaches for the development of vaccines against ticks and tick borne diseases. Ticks Tick Borne Dis. 2016, 7, 573–585. [Google Scholar] [CrossRef]
- Hunt, J.S. Texas fever—Inoculation (Report on certain inoculated animals). Qld. Agric. J. 1899, 4, 216–220. [Google Scholar]
- Bishopp, F.C.; Wood, H.P. The biology of some North American ticks of the genus Dermacentor. Parasitology. 1913, 6, 153–187. [Google Scholar] [CrossRef]
- Johnston, T.H.; Bancroft, M.J. A tick resistant condition in cattle. Proc. R Soc. Qld. 1918, 30, 219–317. [Google Scholar]
- Riek, R.F. Studies on the reactions of animals to infestation with ticks. VI. Resistance of cattle to infestation with the tick Boophilus microplus (Canestrini). Aust. J. Agric. Res. 1962, 13, 532–550. [Google Scholar] [CrossRef]
- Hewetson, R.W. Resistance of cattle to cattle tick, Boophilus microplus. II. The inheritance of resistance by experimental infestations. Aust. J. Agric. Res. 1968, 19, 497–505. [Google Scholar] [CrossRef]
- Hewetson, R.W. Resistance of cattle to cattle tick, Boophilus microplus. III. Development of resistance to experimental infestation by purebred Sahiwal and Shorthorn cattle. Aust. J. Agric. Res. 1971, 22, 331–342. [Google Scholar] [CrossRef]
- Roberts, J.A. Acquisition by the host of resistance to the cattle tick, Boophilus microplus (Canestrini). J. Parasitol. 1968, 54, 657–662. [Google Scholar] [CrossRef]
- Willadsen, P. Immunity to ticks. Adv. Parasitol. 1980, 18, 293–311. [Google Scholar] [CrossRef]
- Willadsen, P.; Jongejan, F. Immunology of the tick-host interaction and the control of ticks and tick-borne diseases. Parasitol. Today 1999, 15, 258–262. [Google Scholar] [CrossRef]
- Piper, E.K.; Jackson, L.A.; Bagnall, N.H.; Kongsuwan, K.K.; Lew, A.E.; Jonsson, N.N. Gene expression in the skin of Bos taurus and Bos indicus cattle infested with the cattle tick, Rhipicephalus (Boophilus) microplus. Vet. Immunol. Immunopathol. 2008, 126, 110–119. [Google Scholar] [CrossRef]
- Piper, E.K.; Jonsson, N.N.; Gondro, C.; Lew-Tabor, A.E.; Moolhuijzenm, P.; Vance, M.E.; Jackson, L.A. Immunological profiles of Bos taurus and Bos indicus cattle infested with the cattle tick, Rhipicephalus (Boophilus) microplus. Clin. Vaccine Immunol. 2009, 16, 1074–1086. [Google Scholar] [CrossRef]
- Domingues, R.; Wohlres-Viana, S.; Reis, D.R.; Teixeira, H.C.; Ferreira, A.P.; Guimarães, S.E.; Prata, M.C.; Furlong, J.; Verneque, R.S.; Machado, M.A. Expression of immune response genes in peripheral blood of cattle infested with Rhipicephalus microplus. Genet. Mol. Res. 2014, 13, 4013–4021. [Google Scholar] [CrossRef]
- Jonsson, N.N.; Piper, E.K.; Constantinoiu, C.C. Host resistance in cattle to infestation with the cattle tick Rhipicephalus microplus. Parasite Immunol. 2014, 36, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Franzin, A.M.; Maruyama, S.R.; Garcia, G.R.; Oliveira, R.P.; Ribeiro, J.M.; Bishop, R.; Maia, A.A.; Moré, D.D.; Ferreira, B.R.; Santos, I.K. Immune and biochemical responses in skin differ between bovine hosts genetically susceptible and resistant to the cattle tick Rhipicephalus microplus. Parasit. Vectors 2017, 10, 51. [Google Scholar] [CrossRef]
- Mota, R.R.; Silva, F.F.; Lopes, P.S.; Tempelman, R.J.; Sollero, B.P.; Aguilar, I.; Cardoso, F.F. Analyses of reaction norms reveal new chromosome regions associated with tick resistance in cattle. Animal 2018, 12, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Robbertse, L.; Richards, S.A.; Clift, S.J.; Barnard, A.C.; Leisewitz, A.; Crafford, J.E.; Maritz-Olivier, C. Comparison of the differential regulation of T and B-lymphocyte subsets in the skin and lymph nodes amongst three cattle breeds as potential mediators of immune-resistance to Rhipicephalus microplus. Ticks Tick Borne Dis. 2018, 9, 976–987. [Google Scholar] [CrossRef]
- Allen, J.R. Tick resistance: Basophils in skin reactions of resistant guinea pigs. Int. J. Parasitol. 1973, 3, 195–200. [Google Scholar] [CrossRef]
- Schleger, A.V.; Lincoln, D.T.; McKenna, R.V.; Kemp, D.H.; Roberts, J.A. Boophilus microplus: Cellular responses to larval attachment and their relationship to host resistance. Aust. J. Biol. Sci. 1976, 29, 499–512. [Google Scholar] [CrossRef]
- Allen, J.R.; Doube, B.M.; Kemp, D.H. Histology of bovine skin reactions to Ixodes holocyclus, Neuman. Can. J. Comp. Med. 1977, 41, 26–35. [Google Scholar]
- McNeil, B.; Dong, X. Peripheral mechanisms of itch. Neurosci. Bull. 2012, 28, 100–110. [Google Scholar] [CrossRef]
- Dong, X.; Dong, X. Peripheral and central mechanisms of itch. Neuron 2018, 98, 482–494. [Google Scholar] [CrossRef]
- Liu, T.; Ji, R.R. New insights into the mechanisms of itch: Are pain and itch controlled by distinct mechanisms? Pflugers Arch. 2013, 465, 1671–1685. [Google Scholar] [CrossRef] [PubMed]
- Wikel, S.K. Vector arthropods and host pain and itch responses. In Arthropod Vector: Controller of Disease Transmission; Eikel, S., Aksoy, S., Dimopoulos, G., Eds.; Elsevier: London, UK, 2017; Volume 2, pp. 13–29. [Google Scholar]
- Kaufman, W.R. Tick-host interaction: A synthesis of current concepts. Parasitol. Today. 1989, 5, 47–56. [Google Scholar] [CrossRef]
- Snowball, G.J. The effect of self-licking by cattle on infestation of cattle tick, Boophilus microplus (Canestrini). Aust. J. Agric. Res. 1956, 7, 227–237. [Google Scholar] [CrossRef]
- De Castro, J.J.; Newson, R.M. Host resistance in cattle tick control. Parasitol. Today 1993, 9, 13–17. [Google Scholar] [CrossRef]
- Bennett, G.F. Boophilus microplus (acarina: Ixodidae): Experimental infestations on cattle restrained from grooming. Exp. Parasitol. 1969, 26, 323–328. [Google Scholar] [CrossRef]
- Koudstaal, D.; Kemp, D.H.; Kerr, J.D. Boophilus microplus: Rejection of larvae from British breed cattle. Parasitology. 1978, 76, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Hart, B.L. Role of grooming in biological control of ticks. Ann. N. Y. Acad. Sci. 2000, 916, 565–569. [Google Scholar] [CrossRef]
- Burke, G.; Wikel, S.K.; Spielman, A.; Telford, S.R.; McKay, K.; Krause, P.J. Tick-borne Infection Study Group. Hypersensitivity to ticks and Lyme disease risk. Emerg. Infect. Dis. 2005, 11, 36–41. [Google Scholar] [CrossRef]
- Hoeppli, R.; Feng, L.C. Histological reactions in the skin due to ectoparasites: Dermacentor sinicus P. Schulze from hedgehog, Haemaphysalis campanulata hoeppliana P. Schulze from dog, Cimex lectularius and Pediculus vestimenti from man. Natl. Med. J. China 1931, 17, 541–556. [Google Scholar]
- Trager, W. Acquired immunity to ticks. J. Parasitol. 1939, 25, 57–81. [Google Scholar] [CrossRef]
- Tatchell, R.J.; Moorhouse, D.E. Neutrophils: Their role in the formation of a tick feeding lesion. Science 1970, 167, 1002–1003. [Google Scholar] [CrossRef] [PubMed]
- Theis, J.H.; Budwiser, P.D. Rhipicephalus sanguineus: Sequential histopathology at the host-arthropod interface. Exp. Parasitol. 1974, 36, 77–105. [Google Scholar] [CrossRef]
- Jellison, W.L.; Kohls, G.M. Tick-host anemia: A secondary anemia induced by Dermacentor andersoni Stiles. J. Parasitol. 1938, 24, 143–154. [Google Scholar] [CrossRef]
- Wikel, S.K.; Allen, J.R. Acquired resistance to ticks. I. Passive transfer of resistance. Immunology 1976, 30, 311–316. [Google Scholar] [PubMed]
- Wikel, S.K.; Allen, J.R. Acquired resistance to ticks. II. Effects of Cyclophosphamide on resistance. Immunology 1976, 30, 479–484. [Google Scholar]
- Brown, S.J.; Askenase, P.W. Cutaneous basophil responses and immune resistance of guinea pigs to ticks: Passive transfer with peritoneal exudate cells or serum. J. Immunol. 1981, 127, 2163–2167. [Google Scholar]
- Wikel, S.K.; Allen, J.R. Acquired resistance to ticks. iii. Cobra venom factor and the resistance response. Immunology 1977, 32, 457–465. [Google Scholar]
- Wikel, S.K. Acquired resistance to ticks: Expression of resistance by C4-deficient guinea pigs. Am. J. Trop. Med. Hyg. 1979, 28, 586–590. [Google Scholar] [CrossRef]
- Allen, J.R.; Khalil, H.M.; Wikel, S.K. Langerhans cells trap tick salivary gland antigens in tick-resistant guinea pigs. J. Immunol. 1979, 122, 563–565. [Google Scholar]
- Bell, J.F.; Stewart, S.J.; Wikel, S.K. Resistance to tick-borne Francisella tularensis by tick-sensitized rabbits: Allergic klendusity. Am. J. Trop. Med. Hyg. 1979, 28, 876–880. [Google Scholar] [CrossRef]
- Nazario, S.; Das, S.; de Silva, A.M.; Deponte, K.; Marcantonio, N.; Anderson, J.F.; Fish, D.; Fikrig, E.; Kantor, F.S. Prevention of Borrelia burgdorferi transmission in guinea pigs by tick immunity. Am. J. Trop. Med. Hyg. 1998, 58, 780–785. [Google Scholar] [CrossRef]
- Tijsse-Klasen, E.; Koopmans, M.P.; Sprong, H. Tick-borne pathogen—Reversed and conventional discovery of disease. Front. Public Health 2014, 2, 73. [Google Scholar] [CrossRef] [PubMed]
- Adrion, E.R.; Aucott, J.; Lemke, K.W.; Weiner, J.P. Health care costs, utilization and patterns of care following Lyme disease. PLoS ONE 2015, 10, e0116767. [Google Scholar] [CrossRef]
- Talagrand-Reboul, E.; Boyer, P.H.; Bergström, S.; Vial, L.; Boulanger, N. Relapsing fevers: Neglected tick-borne diseases. Front. Cell. Infect. Microbiol. 2018, 8, 98. [Google Scholar] [CrossRef] [PubMed]
- Boulanger, N.; Boyer, P.; Talagrand-Reboul, E.; Hansmann, Y. Ticks and tick-borne diseases. Med. Mal. Infect. 2019, 49, 87–97. [Google Scholar] [CrossRef]
- Mac, S.; da Silva, S.R.; Sander, B. The economic burden of Lyme disease and the cost-effectiveness of Lyme disease interventions: A scoping review. PLoS ONE. 2019, 14, e0210280. [Google Scholar] [CrossRef] [PubMed]
- Madison-Antenucci, S.; Kramer, L.D.; Gebhardt, L.L.; Kauffman, E. Emerging Tick-Borne Diseases. Clin. Microbiol. Rev. 2020, 33, e00083-18. [Google Scholar] [CrossRef] [PubMed]
- Colwell, D.D.; Dantas-Torres, F.; Otranto, D. Vector-borne parasitic zoonoses: Emerging scenarios and new perspectives. Vet. Parasitol. 2011, 182, 14–21. [Google Scholar] [CrossRef]
- Karesh, W.B.; Dobson, A.; Lloyd-Smith, J.O.; Lubroth, J.; Dixon, M.A.; Bennett, M.; Aldrich, S.; Harrington, T.; Formenty, P.; Loh, E.H.; et al. Ecology of zoonoses: Natural and unnatural histories. Lancet 2012, 380, 1936–1945. [Google Scholar] [CrossRef]
- Pfäffle, M.; Litwin, N.; Muders, S.V.; Petney, T.N. The ecology of tick-borne diseases. Int. J. Parasitol. 2013, 43, 1059–1077. [Google Scholar] [CrossRef]
- Baneth, G. Tick-borne infections of animals and humans: Common ground. Int. J. Parasitol. 2014, 44, 591–596. [Google Scholar] [CrossRef]
- Kilpatrick, A.M.; Dobson, A.D.M.; Levi, T.; Salkeld, D.J.; Swei, A.; Ginsberg, H.S.; Kjemtrup, A.; Padgett, K.A.; Jensen, P.M.; Fish, D.; et al. Lyme disease ecology in a changing world: Consensus, uncertainty and critical gaps for improving control. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372, 20160117. [Google Scholar] [CrossRef] [PubMed]
- Rizzoli, A.; Silaghi, C.; Obiegala, A.; Rudolf, I.; Hubálek, Z.; Földvári, G.; Plantard, O.; Vayssier-Taussat, M.; Bonnet, S.; Spitalská, E.; et al. Ixodes ricinus and its transmitted pathogens in urban and peri-urban areas in Europe: New hazards and relevance for public health. Front. Public Health 2014, 2, 251. [Google Scholar] [CrossRef] [PubMed]
- Mierzejewska, E.J.; Estrada-Peña, A.; Bajer, A. Spread of Dermacentor reticulatus is associated with the loss of forest area. Exp. Appl. Acarol. 2017, 72, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Eisen, R.J.; Eisen, L. The blacklegged tick, Ixodes scapularis: An increasing public health concern. Trends Parasitol. 2018, 34, 295–309. [Google Scholar] [CrossRef] [PubMed]
- Tokarz, R.; Sameroff, S.; Tagliafierro, T.; Jain, K.; Williams, S.H.; Cucura, D.M.; Rochlin, I.; Monzon, J.; Carpi, G.; Tufts, D.; et al. Identification of Novel Viruses in Amblyomma americanum, Dermacentor variabilis, and Ixodes scapularis Ticks. mSphere 2018, 3, e00614-17. [Google Scholar] [CrossRef]
- Medlock, J.M.; Hansford, K.M.; Bormane, A.; Derdakova, M.; Estrada-Peña, A.; George, J.C.; Golovljova, I.; Jaenson, T.G.; Jensen, J.K.; Jensen, P.M.; et al. Driving forces for changes in geographical distribution of Ixodes ricinus ticks in Europe. Parasit. Vectors 2013, 6, 1. [Google Scholar] [CrossRef]
- Kullberg, B.J.; Vrijmoeth, H.D.; van de Schoor, F.; Hovius, J.W. Lyme borreliosis: Diagnosis and management. BMJ 2020, 369, m1041. [Google Scholar] [CrossRef]
- Kugeler, K.J.; Schwartz, A.M.; Delorey, M.J.; Mead, P.S.; Hinckley, A.F. Estimating the frequency of Lyme Disease diagnoses, United States, 2010-2018. Emerg. Infect. Dis. 2021, 27, 616–619. [Google Scholar] [CrossRef]
- Piesman, J.; Mather, T.N.; Sinsky, R.J.; Spielman, A. Duration of tick attachment and Borrelia burgdorferi transmission. J. Clin. Microbiol. 1987, 25, 557–558. [Google Scholar] [CrossRef]
- Krause, P.J.; Grant-Kels, J.M.; Tahan, S.R.; Dardick, K.R.; Alarcon-Chaidez, F.; Bouchard, K.; Visini, C.; Deriso, C.; Foppa, I.M.; Wikel, S. Dermatologic changes induced by repeated Ixodes scapularis bites and implications for prevention of tick-borne infection. Vector Borne Zoonotic Dis. 2009, 9, 603–610. [Google Scholar] [CrossRef]
- Kimura, R.; Sugita, K.; Ito, A.; Goto, H.; Yamamoto, O. Basophils are recruited and localized at the site of tick bites in humans. J. Cutan. Pathol. 2017, 44, 1091–1093. [Google Scholar] [CrossRef] [PubMed]
- Sokol, C.L.; Medzhitov, R. Emerging functions of basophils in protective and allergic immune responses. Mucosal Immunol. 2010, 3, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Voehringer, D. Protective and pathological roles of mast cells and basophils. Nat. Rev. Immunol. 2013, 13, 362–375. [Google Scholar] [CrossRef] [PubMed]
- Karasuyama, H.; Tabakawa, Y.; Ohta, T.; Wada, T.; Yoshikawa, S. Crucial role for basophils in acquired protective immunity to tick infestation. Front. Physiol. 2018, 9, 1769. [Google Scholar] [CrossRef] [PubMed]
- Glatz, M.; Means, T.; Haas, J.; Steere, A.C.; Müllegger, R.R. Characterization of the early local immune response to Ixodes ricinus tick bites in human skin. Exp. Dermatol. 2017, 26, 263–269. [Google Scholar] [CrossRef]
- Narasimhan, S.; Kurokawa, C.; Diktas, H.; Strank, N.O.; Černý, J.; Murfin, K.; Cao, Y.; Lynn, G.; Trentleman, J.; Wu, M.J.; et al. Ixodes scapularis saliva components that elicit responses associated with acquired tick-resistance. Ticks Tick Borne Dis. 2020, 11, 101369. [Google Scholar] [CrossRef]
- Strnad, M.; Grubhoffer, L.; Rego, R.O.M. Novel targets and strategies to combat borreliosis. Appl. Microbiol. Biotechnol. 2020, 104, 1915–1925. [Google Scholar] [CrossRef]
- Chmelař, J.; Kotál, J.; Kovaříková, A.; Kotsyfakis, M. The use of tick salivary proteins as novel therapeutics. Front. Physiol. 2019, 10, 812. [Google Scholar] [CrossRef]
- Wikel, S.K. Host immunity to ticks. Ann. Rev. Entomol. 1996, 41, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Wikel, S.K. Ticks and tick-borne pathogens at the cutaneous interface: Host defenses, tick countermeasures, and a suitable environment for pathogen establishment. Front. Microbiol. 2013, 4, 337. [Google Scholar] [CrossRef]
- Wikel, S.K. Tick-host-pathogen systems immunobiology: An interactive trio. Front. Biosci. 2018, 22, 2105–2121. [Google Scholar] [CrossRef] [PubMed]
- Chmelař, J.; Kotál, J.; Kopecký, J.; Pedra, J.H.; Kotsyfakis, M. All for one and one for all on the tick-host battlefield. Trends Parasitol. 2016, 32, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Šimo, L.; Kazimirova, M.; Richardson, J.; Bonne, S.I. The essential role of tick salivary glands and saliva in tick feeding and pathogen transmission. Front. Cell Infect. Microbiol. 2017, 7, 281. [Google Scholar] [CrossRef]
- Narasimhan, S.; Kurookawa, C.; DeBlasio, M.; Matias, J.; Sajid, A.; Pal, U.; Lynn, G.; Fikrig, E. Acquired resistance to ticks: The trail is hot. Parasite Immunol. 2021, e12808. [Google Scholar] [CrossRef]
- Allen, J.R. Immunology of interactions between ticks and laboratory animals. Exp. Appl. Acarol. 1989, 7, 5–13. [Google Scholar] [CrossRef]
- Wikel, S.K. Tick modulation of host immunity: An important factor in pathogen transmission. Int. J. Parasitol. 1999, 29, 851–859. [Google Scholar] [CrossRef]
- Karasuyama, H.; Miyake, K.; Yoshikawa, S. Immunobiology of acquired resistance to ticks. Front. Immunol. 2020, 11, 601504. [Google Scholar] [CrossRef]
- Bernard, Q.; Grillon, A.; Lenormand, C.; Ehret-Sabatier, L.; Boulanger, N. Skin interface, a key player for Borrelia multiplication and persistence in Lyme borreliosis. Trends Parasitol. 2020, 36, 304–314. [Google Scholar] [CrossRef] [PubMed]
- denHollander, N.; Allen, J.R. Dermacentor variabilis: Resistance to ticks acquired by mast cell-deficient and other strains of mice. Exp. Parasitol. 1985, 59, 169–179. [Google Scholar] [CrossRef]
- Steeves, E.B.; Allen, J.R. Basophils in skin reactions of mast cell-deficient mice infested with Dermacentor variabilis. Int. J. Parasitol. 1990, 20, 655–667. [Google Scholar] [CrossRef]
- Matsuda, H.; Fukui, K.; Kiso, Y.; Kitamura, Y. Inability of genetically mast cell-deficient W/Wv mice to acquire resistance against larval Haemaphysalis longicornis ticks. J. Parasitol. 1985, 71, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Tabakawa, Y.; Ohta, T.; Yoshikawa, S.; Robinson, E.J.; Yamaji, K.; Ishiwata, K.; Kawano, Y.; Miyake, K.; Yamanishi, Y.; Ohtsu, H.; et al. Histamine released from skin-infiltrating basophils but not mast cells is crucial for acquired tick resistance in mice. Front. Immunol. 2018, 9, 1540. [Google Scholar] [CrossRef]
- Ohta, T.; Yoshikawa, S.; Tabakawa, Y.; Yamaji, K.; Ishiwata, K.; Shitara, H.; Taya, C.; Oh-Hora, M.; Kawano, Y.; Miyake, K.; et al. Skin CD4+ memory T cells play an essential role in acquired anti-tick immunity through interleukin-3-mediated basophil recruitment to tick-feeding sites. Front. Immunol. 2017, 8, 1348. [Google Scholar] [CrossRef] [PubMed]
- Wada, T.; Ishiwata, K.; Koseki, H.; Ishikura, T.; Ugajin, T.; Ohnuma, N.; Obata, K.; Ishikawa, R.; Yoshikawa, S.; Mukai, K.; et al. Selective ablation of basophils in mice reveals their nonredundant role in acquired immunity against ticks. J. Clin. Investig. 2010, 120, 2867–2875. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, S.; Miyake, K.; Kamiya, A.; Karasuyama, H. The role of basophils in acquired protective immunity to tick infestation. Parasite Immunol. 2021, 43, e12804. [Google Scholar] [CrossRef]
- Willadsen, P.; Wood, G.M.; Riding, G.A. The relation between skin histamine concentration, histamine sensitivity, and the resistance of cattle to the tick, Boophilus microplus. Z. Parasitenkd. 1979, 59, 87–93. [Google Scholar] [CrossRef]
- Paine, S.H.; Kemp, D.H.; Allen, J.R. In vitro feeding of Dermacentor andersoni (Stiles): Effects of histamine and other mediators. Parasitology 1983, 86, 419–428. [Google Scholar] [CrossRef]
- Wikel, S.K. Histamine content of tick attachment sites and the effects of H1 and H2 histamine antagonists on the expression of resistance. Ann. Trop. Med. Parasitol. 1982, 76, 179–185. [Google Scholar] [CrossRef]
- Chinery, W.A.; Ayitey-Smith, E. Histamine blocking agent in the salivary gland homogenate of the tick Rhipicephalus sanguineus sanguineus. Nature 1977, 265, 366–367. [Google Scholar] [CrossRef] [PubMed]
- Paesen, G.C.; Adams, P.L.; Harlos, K.; Nuttall, P.A.; Stuart, D.I. Tick histamine-binding proteins: Isolation, cloning, and three-dimensional structure. Mol. Cell. 1999, 3, 661–671. [Google Scholar] [CrossRef]
- Paesen, G.C.; Adams, P.L.; Nuttall, P.A.; Stuart, D.L. Tick histamine-binding proteins: Lipocalins with a second binding cavity. Biochim. Biophys. Acta 2000, 1482, 92–101. [Google Scholar] [CrossRef]
- Sangamnatdej, S.; Paesen, G.C.; Slovak, M.; Nuttall, P.A. A high affinity serotonin- and histamine-binding lipocalin from tick saliva. Insect Mol. Biol. 2002, 11, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Neelakanta, G.; Sultana, H.; Sonenshine, D.E.; Andersen, J.F. Identification and characterization of a histamine-binding lipocalin-like molecule from the relapsing fever tick Ornithodoros turicata. Insect Mol. Biol. 2018, 27, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Narasimhan, S.; Zhang, L.; Liu, L.; Wang, P.; Fikrig, E. Tick histamine release factor is critical for Ixodes scapularis engorgement and transmission of the Lyme disease agent. PLoS Pathog. 2010, 6, e1001205. [Google Scholar] [CrossRef]
- Wikel, S.K. Immune responses to arthropods and their products. Annu. Rev. Entomol. 1982, 27, 21–48. [Google Scholar] [CrossRef]
- Kotál, J.; Langhansová, H.; Lieskovská, J.; Andersen, J.F.; Francischetti, I.M.; Chavakis, T.; Kopecký, J.; Pedra, J.H.; Kotsyfakis, M.; Chmelař, J. Modulation of host immunity by tick saliva. J. Proteomics. 2015, 128, 58–68. [Google Scholar] [CrossRef]
- Nuttall, P.A. Tick saliva and its role in pathogen transmission. Wien. Klin. Wochenschr. 2019. [Google Scholar] [CrossRef]
- Martins, L.; Bensaoud, C.; Kotal, J.; Chmelar, J.; Kotsyfakis, M. Tick salivary gland transcriptomics and proteomics. Parasite Immunol. 2021, 43, e12807. [Google Scholar] [CrossRef]
- Kazimírová, M.; Štibrániová, I. Tick salivary compounds: Their role in modulation of host defences and pathogen transmission. Front. Cell. Infect. Microbiol. 2013, 3, 43. [Google Scholar] [CrossRef]
- Ribeiro, J.; Alarcon-Chaidez, F.; Francischetti, I.M.B.; Mans, B.; Mather, T.N.; Valenzuela, J.G.; Wikel, S.K. An annotated catalog of salivary gland transcripts from Ixodes scapularis ticks. Insect Biochem. Mol. Biol. 2006, 36, 111–129. [Google Scholar] [CrossRef] [PubMed]
- Alarcon-Chaidez, F.J.; Sun, J.; Wikel, S.K. Construction and characterization of a cDNA library from the salivary glands of Dermacentor andersoni Stiles (Acari: Ixodidae). Insect Biochem. Mol. Biol. 2007, 37, 48–71. [Google Scholar] [CrossRef]
- Chmelař, J.; Anderson, J.M.; Mu, J.; Jochim, R.C.; Valenzuela, J.G.; Kopecký, J. Insight into the sialome of the castor bean tick, Ixodes ricinus. BMC Genom. 2008, 9, 233. [Google Scholar] [CrossRef] [PubMed]
- Aljamali, M.N.; Hern, L.; Kupfer, D.; Downard, S.; So, S.; Roe, B.A.; Sauer, J.R.; Essenberg, R.C. Transcriptome analysis of the salivary glands of the female tick Amblyomma americanum (Acari: Ixodidae). Insect Mol. Biol. 2009, 18, 129–154. [Google Scholar] [CrossRef] [PubMed]
- Karim, S.; Ribeiro, J.M. An Insight into the sialome of the Lone Star tick, Amblyomma americanum, with a glimpse on its time dependent gene expression. PLoS ONE 2015, 10, e0131292. [Google Scholar] [CrossRef] [PubMed]
- Esteves, E.; Maruyama, S.R.; Kawahara, R.; Fujita, A.; Martins, L.A.; Righi, A.A.; Costa, F.B.; Palmisano, G.; Labruna, M.B.; Sá-Nunes, A.; et al. Analysis of the salivary gland transcriptome of unfed and partially fed Amblyomma sculptum ticks and descriptive proteome of the saliva. Front. Cell. Infect. Microbiol. 2017, 7, 476. [Google Scholar] [CrossRef]
- Bensaoud, C.; Aounallah, H.; Sciani, J.M.; Faria, F.; Chudzinski-Tavassi, A.M.; Bouattour, A.; M’ghirbi, Y. Proteomic informed by transcriptomic for salivary glands components of the camel tick Hyalomma dromedarii. BMC Genom. 2019, 20, 675. [Google Scholar] [CrossRef]
- Narasimhan, S.; Booth, C.J.; DePonte, K.; Wu, M.J.; Liang, X.; Mohanty, S.; Kantor, F.; Fikrig, E. Host-specific expression of Ixodes scapularis salivary genes. Ticks Tick Borne Dis. 2019, 10, 386–397. [Google Scholar] [CrossRef]
- Oliveira, C.J.; Sá-Nunes, A.; Francischetti, I.M.; Carregaro, V.; Anatriello, E.; Silva, J.S.; Santos, I.K.; Ribeiro, J.M.; Ferreira, B.R. Deconstructing tick saliva: Non-protein molecules with potent immunomodulatory properties. J. Biol. Chem. 2011, 286, 10960–10969. [Google Scholar] [CrossRef]
- Giachetto, P.F.; Cunha, R.C.; Nhani, A., Jr.; Garcia, M.V.; Ferro, J.A.; Andreotti, R. Gene expression in the salivary gland of Rhipicephalus (Boophilus) microplus fed on tick-susceptible and tick-resistant hosts. Front. Cell. Infect. Microbiol. 2020, 9, 477. [Google Scholar] [CrossRef] [PubMed]
- Trager, W. Further observations on acquired immunity to the tick, Dermacentor variabilis Say. J. Parasitol. 1939, 25, 137–139. [Google Scholar] [CrossRef]
- Ackerman, S.; Clare, F.B.; McGill, T.W.; Sonenshine, D.E. Passage of host serum components, including antibody, across the digestive tract of Dermacentor variabilis (Say). J. Parasitol. 1981, 67, 737–740. [Google Scholar] [CrossRef] [PubMed]
- McGowan, M.J.; Barker, R.W.; Homer, J.T.; McNew, R.W.; Holscher, K.H. Success of tick feeding on calves immunized with Amblyomma americanum (Acari: Ixodidae) extract. J. Med. Entomol. 1981, 18, 328–331. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, P.R.; Allen, J.R. A test of the efficacy of immunizing cattle against Rocky Mountain wood ticks. J. Entomol. Soc. Brit. Columbia 1983, 80, 37–41. [Google Scholar]
- Wikel, S.K.; Whelen, A.C. Ixodid-host immune interaction. Identification and characterization of relevant antigens and tick-induced host immunosuppression. Vet. Parasitol. 1986, 20, 149–174. [Google Scholar] [CrossRef]
- Rodríguez-Mallon, A. Developing anti-tick vaccines. Methods Mol. Biol. 2016, 1404, 243–259. [Google Scholar] [CrossRef] [PubMed]
- Quiroz-Castañeda, R.E.; Amaro-Estrada, I.; Rodríguez-Camarillo, S.D. Anaplasma marginale: Diversity, virulence, and vaccine landscape through a genomics approach. Biomed. Res. Int. 2016, 2016, 9032085. [Google Scholar] [CrossRef] [PubMed]
- Beugnet, F.; Moreau, Y. Babesiosis. Rev. Sci. Tech. 2015, 34, 627–639. [Google Scholar] [CrossRef]
- Allsopp, B.A. Heartwater—Ehrlichia ruminantium infection. Rev. Sci. Tech. 2015, 34, 557–568. [Google Scholar] [CrossRef]
- Agina, O.A.; Shaari, M.R.; Isa, N.M.M.; Ajat, M.; Zamri-Saad, M.; Hamzah, H. Clinical pathology, immunopathology and advanced vaccine technology in bovine theileriosis: A review. Pathogens 2020, 9, 697. [Google Scholar] [CrossRef] [PubMed]
- Schlein, Y.; Sira, D.T.; Jacobson, R.L. The passage of serum immunoglobulins through the gut of Sarcophaga falculata, Pand. Ann. Trop. Med. Parasitol. 1976, 70, 227–230. [Google Scholar] [CrossRef] [PubMed]
- Cooley, R.A.; Kohls, G.M. The Argasidae of North America, Central America and Cuba. Am. Midl. Nat. Monograph. 1944, 1, 1–152. [Google Scholar]
- Ackerman, S.; Floyd, M.; Sonenshine, D.E. Artificial immunity to Dermacentor variabilis (Acari: Ixodidae): Vaccination using tick antigens. J. Med. Entomol. 1980, 17, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Nuttall, P.A. Excretion of host immunoglobulin in tick saliva and detection of IgG-binding proteins in tick haemolymph and salivary glands. Parasitology 1994, 109, 525–530. [Google Scholar] [CrossRef]
- Brossard, M.; Rais, O. Passage of hemolysins through the midgut epithelium of female Ixodes ricinus L. fed on rabbits infested or reinfested with ticks. Experientia 1984, 40, 561–563. [Google Scholar] [CrossRef]
- Minoura, H.; Chinzei, Y.; Kitamura, S. Ornithodoros moubata: Host immunoglobulin G in tick hemolymph. Exp. Parasitol. 1985, 60, 355–363. [Google Scholar] [CrossRef]
- Ben-Yakir, D. Quantitative studies of host immunoglobulin G in the hemolymph of ticks (Acari). J. Med. Entomol. 1989, 26, 243–246. [Google Scholar] [CrossRef] [PubMed]
- Vaz, I.S., Jr.; Martinez, R.H.; Oliveira, A.; Heck, A.; Logullo, C.; Gonzales, J.C.; Dewes, H.; Masuda, A. Functional bovine immunoglobulins in Boophilus microplus hemolymph. Vet. Parasitol. 1996, 62, 155–160. [Google Scholar] [CrossRef]
- Jasinskas, A.; Barbour, A.G. The Fc fragment mediates the uptake of immunoglobulin C from the midgut to hemolymph in the ixodid tick Amblyomma americanum (Acari: Ixodidae). J. Med. Entomol. 2005, 42, 359–366. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Allen, J.R.; Humphreys, S.J. Immunisation of guinea pigs and cattle against ticks. Nature 1979, 280, 491–493. [Google Scholar] [CrossRef]
- Willadsen, P.; Kemp, D.H. Vaccination with ‘concealed’ antigens for tick control. Parasitol. Today 1988, 4, 196–198. [Google Scholar] [CrossRef]
- Tellam, R.L.; Smith, D.; Kemp, D.H.; Willadsen, P. Vaccination against ticks. In Animal Parasite Control Utilizing Biotechnology; Yong, W.K., Ed.; CRC Press: Boca Raton, FL, USA, 1992; pp. 303–331. [Google Scholar]
- Canales, M.; Labruna, M.B.; Soares, J.F.; Prudencio, C.R.; de la Fuente, J. Protective efficacy of bacterial membranes containing surface-exposed BM95 antigenic peptides for the control of cattle tick infestations. Vaccine 2009, 27, 7244–7248. [Google Scholar] [CrossRef]
- Kumar, B.; Azhahianambi, P.; Ray, D.D.; Chaudhuri, P.; de La Fuente, J.; Kumar, R.; Ghosh, S. Comparative efficacy of rHaa86 and rBm86 against Hyalomma anatolicum anatolicum and Rhipicephalus (Boophilus) microplus. Parasite Immunol. 2012, 34, 297–301. [Google Scholar] [CrossRef] [PubMed]
- García-García, J.C.; Montero, C.; Redondo, M.; Vargas, M.; Canales, M.; Boue, O.; Rodríguez, M.; Joglar, M.; Machado, H.; González, I.L.; et al. Control of ticks resistant to immunization with Bm86 in cattle vaccinated with the recombinant antigen Bm95 isolated from the cattle tick, Boophilus microplus. Vaccine 2000, 18, 2275–2287. [Google Scholar] [CrossRef]
- Rodríguez Mallón, A.; Javier González, L.; Encinosa Guzmán, P.E.; Bechara, G.H.; Sanches, G.S.; Pousa, S.; Cabrera, G.; Cabrales, A.; Garay, H.; Mejías, R.; et al. Functional and mass spectrometric evaluation of an anti-tick antigen based on the P0 peptide conjugated to Bm86 protein. Pathogens 2020, 9, 513. [Google Scholar] [CrossRef] [PubMed]
- Nijhof, A.M.; Balk, J.A.; Postigo, M.; Rhebergen, A.M.; Taoufik, A.; Jongejan, F. Bm86 homologues and novel ATAQ proteins with multiple epidermal growth factor (EGF)-like domains from hard and soft ticks. Int. J. Parasitol. 2010, 40, 1587–1597. [Google Scholar] [CrossRef][Green Version]
- Aguirre Ade, A.; Lobo, F.P.; Cunha, R.C.; Garcia, M.V.; Andreotti, R. Design of the ATAQ peptide and its evaluation as an immunogen to develop a Rhipicephalus vaccine. Vet. Parasitol. 2016, 221, 30–38. [Google Scholar] [CrossRef]
- Knorr, S.; Anguita, J.; Cortazar, J.T.; Hajdusek, O.; Kopáček, P.; Trentelman, J.J.; Kershaw, O.; Hovius, J.W.; Nijhof, A.M. Preliminary evaluation of tick protein extracts and recombinant ferritin 2 as anti-tick vaccines targeting Ixodes ricinus in cattle. Front. Physiol. 2018, 9, 1696. [Google Scholar] [CrossRef]
- Finn, R.N.; Cedra, J. Evolution and functional diversity of aquaporins. Biol. Bull. 2015, 229, 6–23. [Google Scholar] [CrossRef]
- Campbell, E.M.; Ball, A.; Hoppler, S.; Bowman, A.S. Invertebrate aquaporins: A review. J. Comp. Physiol. B 2008, 178, 935–955. [Google Scholar] [CrossRef]
- Kaufman, W.R.; Phillips, J.E. Ion and water balance in the ixodid tick Dermacentor andersoni. I. Routes of ion and water excretion. J. Exp. Biol. 1973, 58, 523–536. [Google Scholar] [CrossRef]
- Sauer, J.R.; McSwain, J.L.; Bowman, A.S.; Essenberg, R.C. Tick salivary gland physiology. Annu. Rev. Entomol. 1995, 40, 245–267. [Google Scholar] [CrossRef]
- Bowman, A.S.; Sauer, J.R. Tick salivary glands: Function, physiology and future. Parasitology 2004, 129, S67–S81. [Google Scholar] [CrossRef]
- Hussein, H.E.; Scoles, G.A.; Ueti, M.W.; Suarez, C.E.; Adham, F.K.; Guerrero, F.D.; Bastos, R.G. Targeted silencing of the aquaporin 2 gene of Rhipicephalus (Boophilus) microplus reduces tick fitness. Parasit. Vectors 2015, 8, 618. [Google Scholar] [CrossRef]
- Guerrero, F.D.; Andreotti, R.; Bendele, K.G.; Cunha, R.C.; Miller, R.J.; Yeater, K.; Pérez de León, A.A. Rhipicephalus (Boophilus) microplus aquaporin as an effective vaccine antigen to protect against cattle tick infestations. Parasit. Vectors 2014, 7, 475. [Google Scholar] [CrossRef]
- Évora, P.M.; Sanches, G.S.; Guerrero, F.D.; León, A.P.; Bechara, G.H. Immunogenic potential of Rhipicephalus (Boophilus) microplus aquaporin 1 against Rhipicephalus sanguineus in domestic dogs. Rev. Bras. Parasitol. Vet. 2017, 26, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Contreras, M.; de la Fuente, J. Control of infestation by Ixodes ricinus tick larvae in rabbits vaccinated with aquaporin recombinant antigens. Vaccine 2017, 35, 1323–1328. [Google Scholar] [CrossRef] [PubMed]
- Ndekezi, C.; Nkamwesiga, J.; Ochwo, S.; Kimuda, M.P.; Mwiine, F.N.; Tweyongyere, R.; Amanyire, W.; Muhanguzi, D. Identification of Ixodid tick-specific aquaporin-1 potential anti-tick vaccine epitopes: An in-silico analysis. Front. Bioeng. Biotechnol. 2019, 7, 236. [Google Scholar] [CrossRef] [PubMed]
- Manzano-Román, R.; García-Varas, S.; Encinas-Grandes, A.; Pérez-Sánchez, R. Purification and characterization of a 45-kDa concealed antigen from the midgut membranes of Ornithodoros erraticus that induces lethal anti-tick immune responses in pigs. Vet. Parasitol. 2007, 145, 314–325. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pérez-Sánchez, R.; Manzano-Román, R.; Obolo-Mvoulouga, P.; Oleaga, A. In silico selection of functionally important proteins from the mialome of Ornithodoros erraticus ticks and assessment of their protective efficacy as vaccine targets. Parasit. Vectors 2019, 12, 508. [Google Scholar] [CrossRef]
- Guinat, C.; Gogin, A.; Blome, S.; Keil, G.; Pollin, R.; Pfeiffer, D.U.; Dixon, L. Transmission routes of African swine fever virus to domestic pigs: Current knowledge and future research directions. Vet. Rec. 2016, 178, 262–267. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente, J.; Maritz-Olivier, C.; Naranjo, V.; Ayoubi, P.; Nijhof, A.M.; Almazán, C.; Canales, M.; Pérez de la Lastra, J.M.; Galindo, R.C.; Blouin, E.F.; et al. Evidence of the role of tick subolesin in gene expression. BMC Genom. 2008, 9, 372. [Google Scholar] [CrossRef] [PubMed]
- Artigas-Jerónimo, S.; Villar, M.; Cabezas-Cruz, A.; Valdés, J.J.; Estrada-Peña, A.; Alberdi, P.; de la Fuente, J. Functional evolution of subolesin/akirin. Front. Physiol. 2018, 9, 1612. [Google Scholar] [CrossRef] [PubMed]
- Olds, C.L.; Mwaura, S.; Odongo, D.O.; Scoles, G.A.; Bishop, R.; Daubenberger, C. Induction of humoral immune response to multiple recombinant Rhipicephalus appendiculatus antigens and their effect on tick feeding success and pathogen transmission. Parasit. Vectors 2016, 9, 484. [Google Scholar] [CrossRef]
- Trentelman, J.J.A.; Teunissen, H.; Kleuskens, J.A.G.M.; van de Crommert, J.; de la Fuente, J.; Hovius, J.W.R.; Schetters, T.P.M. A combination of antibodies against Bm86 and Subolesin inhibits engorgement of Rhipicephalus australis (formerly Rhipicephalus microplus) larvae in vitro. Parasit. Vectors 2019, 12, 362. [Google Scholar] [CrossRef]
- Hassan, I.A.; Wang, Y.; Zhou, Y.; Cao, J.; Zhang, H.; Zhou, J. Cross protection induced by combined Subolesin-based DNA and protein immunizations against adult Haemaphysalis longicornis. Vaccine 2020, 38, 907–915. [Google Scholar] [CrossRef]
- Kasaija, P.D.; Contreras, M.; Kabi, F.; Mugerwa, S.; de la Fuente, J. Vaccination with recombinant subolesin antigens provides cross-tick species protection in Bos indicus and crossbred cattle in Uganda. Vaccines 2020, 8, 319. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Sauer, J.R.; Bowman, A.S.; Dillwith, J.W. Identification and characterization of anticoagulant activities in the saliva of the lone star tick, Amblyomma americanum (L.). J. Parasitol. 1997, 83, 38–43. [Google Scholar] [CrossRef]
- Francischetti, I.M.; Valenzuela, J.G.; Andersen, J.F.; Mather, T.N.; Ribeiro, J.M. Ixolaris, a novel recombinant tissue factor pathway inhibitor (TFPI) from the salivary gland of the tick, Ixodes scapularis: Identification of factor X and factor Xa as scaffolds for the inhibition of factor VIIa/tissue factor complex. Blood 2002, 99, 3602–3612. [Google Scholar] [CrossRef]
- Maritz-Olivier, C.; Stutzer, C.; Jongejan, F.; Neitz, A.W.; Gaspar, A.R. Tick anti-hemostatics: Targets for future vaccines and therapeutics. Trends Parasitol. 2007, 23, 397–407. [Google Scholar] [CrossRef][Green Version]
- Costa, G.C.A.; Ribeiro, I.C.T.; Melo-Junior, O.; Gontijo, N.F.; Sant’Anna, M.R.V.; Pereira, M.H.; Pessoa, G.C.D.; Koerich, L.B.; Oliveira, F.; Valenzuela, J.G.; et al. Amblyomma sculptum salivary protease inhibitors as potential anti-tick vaccines. Front. Immunol. 2021, 11, 611104. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Parizi, L.F.; Guizzo, M.G.; Tirloni, L.; Seixas, A.; Vaz Ida, S., Jr.; Termignoni, C. Immunoprotective potential of a Rhipicephalus (Boophilus) microplus metalloprotease. Vet. Parasitol. 2015, 207, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Suppan, J.; Engel, B.; Marchetti-Deschmann, M.; Nürnberger, S. Tick attachment cement—Reviewing the mysteries of a biological skin plug system. Biol. Rev. Camb. Philos. Soc. 2018, 93, 1056–1076. [Google Scholar] [CrossRef] [PubMed]
- Anguita, J.; Ramamoorthi, N.; Hovius, J.W.; Das, S.; Thomas, V.; Persinski, R.; Conze, D.; Askenase, P.W.; Rincón, M.; Kantor, F.S.; et al. Salp15, an Ixodes scapularis salivary protein, inhibits CD4(+) T cell activation. Immunity 2002, 16, 849–859. [Google Scholar] [CrossRef]
- Juncadella, I.J.; Anguita, J. The immunosuppresive tick salivary protein, Salp15. Adv. Exp. Med. Biol. 2009, 666, 121–131. [Google Scholar] [CrossRef]
- Juncadella, I.J.; Garg, R.; Ananthnarayanan, S.K.; Yengo, C.M.; Anguita, J. T-cell signaling pathways inhibited by the tick saliva immunosuppressor, Salp15. FEMS Immunol. Med. Microbiol. 2007, 49, 433–438. [Google Scholar] [CrossRef]
- Garg, R.; Juncadella, I.J.; Ramamoorthi, N.; Ananthanarayanan, S.K.; Thomas, V.; Rincón, M.; Krueger, J.K.; Fikrig, E.; Yengo, C.M.; Anguita, J.; et al. Cutting edge: CD4 is the receptor for the tick saliva immunosuppressor, Salp15. J. Immunol. 2006, 177, 6579–6583. [Google Scholar] [CrossRef]
- Ramamoorthi, N.; Narasimhan, S.; Pal, U.; Bao, F.; Yang, X.F.; Fish, D.; Anguita, J.; Norgard, M.V.; Kantor, F.S.; Anderson, J.F.; et al. The Lyme disease agent exploits a tick protein to infect the mammalian host. Nature 2005, 436, 573–577. [Google Scholar] [CrossRef]
- Kolb, P.; Wallich, R.; Nassal, M. Whole-chain tick saliva proteins presented on hepatitis B virus capsid-like particles induce high-titered antibodies with neutralizing potential. PLoS ONE 2015, 10, e0136180. [Google Scholar] [CrossRef]
- Valenzuela, J.G.; Charlab, R.; Mather, T.N.; Ribeiro, J.M. Purification, cloning, and expression of a novel salivary anticomplement protein from the tick, Ixodes scapularis. J. Biol. Chem. 2000, 275, 18717–18723. [Google Scholar] [CrossRef] [PubMed]
- Ullmann, A.J.; Dolan, M.C.; Sackal, C.A.; Fikrig, E.; Piesman, J.; Zeidner, N.S. Immunization with adenoviral-vectored tick salivary gland proteins (SALPs) in a murine model of Lyme borreliosis. Ticks Tick Borne Dis. 2013, 4, 160–163. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Wang, P.; Adusumilli, S.; Booth, C.J.; Narasimhan, S.; Anguita, J.; Fikrig, E. Antibodies against a tick protein, Salp15, protect mice from the Lyme disease agent. Cell Host Microbe. 2009, 6, 482–492. [Google Scholar] [CrossRef]
- Parizi, L.F.; Rangel, C.K.; Sabadin, G.A.; Saggin, B.F.; Kiio, I.; Xavier, M.A.; da Silva Matos, R.; Camargo-Mathias, M.I.; Seixas, A.; Konnai, S.; et al. Rhipicephalus microplus cystatin as a potential cross-protective tick vaccine against Rhipicephalus appendiculatus. Ticks Tick Borne Dis. 2020, 11, 101378. [Google Scholar] [CrossRef] [PubMed]
- Huercha, S.R.; Li, M.; Fan, X.; Hu, Z.; Wu, L.; Li, Y.; Zhang, W.; Zhang, Y.; Ma, Y.; Bayin, C. Characterization of glutathione S-transferase of Dermacantor marginatus and effect of the recombinant antigen as a potential anti-tick vaccine. Vet. Parasitol. 2020, 279, 109043. [Google Scholar] [CrossRef] [PubMed]
- Almazán, C.; Fourniol, L.; Rakotobe, S.; Šimo, L.; Bornères, J.; Cote, M.; Peltier, S.; Maye, J.; Versillé, N.; Richardson, J.; et al. Failed disruption of tick feeding, viability, and molting after immunization of mice and sheep with recombinant Ixodes ricinus salivary proteins IrSPI and IrLip1. Vaccines 2020, 8, 475. [Google Scholar] [CrossRef]
- Narasimhan, S.; Fikrig, E. Tick microbiome: The force within. Trends Parasitol. 2015, 31, 315–323. [Google Scholar] [CrossRef]
- Bonnet, S.I.; Binetruy, F.; Hernández-Jarguín, A.M.; Duron, O. The tick microbiome: Why non-pathogenic microorganisms matter in tick biology and pathogen transmission. Front. Cell. Infect. Microbiol. 2017, 7, 236. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; Cabezas-Cruz, A.; Pollet, T.; Vayssier-Taussat, M.; Cosson, J.F. High throughput sequencing and network analysis disentangle the microbial communities of ticks and hosts within and between ecosystems. Front. Cell. Infect. Microbiol. 2018, 8, 236. [Google Scholar] [CrossRef]
- Greay, T.L.; Gofton, A.W.; Paparini, A.; Ryan, U.M.; Oskam, C.L.; Irwin, P.J. Recent insights into the tick microbiome gained through next-generation sequencing. Parasit. Vectors 2018, 11, 12. [Google Scholar] [CrossRef]
- Bonnet, S.I.; Pollet, T. Update on the intricate tango between tick microbiomes and tick-borne pathogens. Parasite Immunol. 2020, 13, e12813. [Google Scholar] [CrossRef]
- Andreotti, R.; Pérez de León, A.A.; Dowd, S.E.; Guerrero, F.D.; Bendele, K.G.; Scoles, G.A. Assessment of bacterial diversity in the cattle tick Rhipicephalus (Boophilus) microplus through tag-encoded pyrosequencing. BMC Microbiol. 2011, 11, 6. [Google Scholar] [CrossRef]
- Carpi, G.; Cagnacci, F.; Wittekindt, N.E.; Zhao, F.; Qi, J.; Tomsho, L.P.; Drautz, D.I.; Rizzoli, A.; Schuster, S.C. Metagenomic profile of the bacterial communities associated with Ixodes ricinus ticks. PLoS ONE 2011, 6, e25604. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente, J.; Antunes, S.; Bonnet, S.; Cabezas-Cruz, A.; Domingos, A.G.; Estrada-Peña, A.; Johnson, N.; Kocan, K.M.; Mansfield, K.L.; Nijhof, A.M.; et al. Tick-pathogen interactions and vector competence: Identification of molecular drivers for tick-borne diseases. Front. Cell. Infect. Microbiol. 2017, 7, 114. [Google Scholar] [CrossRef] [PubMed]
- Narasimhan, S.; Schuijt, T.J.; Abraham, N.M.; Rajeevan, N.; Coumou, J.; Graham, M.; Robson, A.; Wu, M.J.; Daffre, S.; Hovius, J.W.; et al. Modulation of the tick gut milieu by a secreted tick protein favors Borrelia burgdorferi colonization. Nat. Commun. 2017, 8, 184. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.A.; Schmidtmann, E.T. Isolation of aerobic microbes from Ixodes scapularis (Acari: Ixodidae), the vector of Lyme disease in the eastern United States. J. Econ. Entomol. 1998, 91, 864–868. [Google Scholar] [CrossRef]
- De la Fuente, J.; Contreras, M. Tick vaccines: Current status and future directions. Expert Rev. Vaccines 2015, 14, 1367–1376. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Peña, A.; Cabezas-Cruz, A.; Obregón, D. Resistance of tick gut microbiome to anti-tick vaccines, pathogen infection and antimicrobial peptides. Pathogens 2020, 9, 309. [Google Scholar] [CrossRef]
- Ben-Yakir, D.; Barker, R.W. The development of Amblyomma americanum and Dermacentor variabilis (Acari: Ixodidae) fed on rabbits immunized with tick hemolymph. Parasitol. Res. 1987, 73, 284–288. [Google Scholar] [CrossRef]
- Grubhoffer, L.; Rudenko, N.; Vancova, M.; Golovchenko, M.; Sterba, J. Circulatory system and hemolymph. In Biology of Ticks, 2nd ed.; Sonenshine, D.E., Roe, R.M., Eds.; Oxford University Press: New York, NY, USA, 2014; Volume 1, pp. 258–286. [Google Scholar]
- Araman, S.F. Protein digestion and synthesis in Ixodid females. In Recent Advances in Acarology; Rodriguez, J., Ed.; Academic Press: New York, NY, USA, 1979; Volume 1, pp. 385–395. [Google Scholar]
- Wang, H.; Nuttall, P.A. Immunoglobulin-binding proteins in ticks: New target for vaccine development against a blood-feeding parasite. Cell. Mol. Life Sci. 1999, 56, 286–295. [Google Scholar] [CrossRef]
- Imamura, S.; da Silva Vaz Junior, I.; Sugino, M.; Ohashi, K.; Onuma, M. A serine protease inhibitor (serpin) from Haemaphysalis longicornis as an anti-tick vaccine. Vaccine 2005, 23, 1301–1311. [Google Scholar] [CrossRef]
- Mitchell, R.D., 3rd; Sonenshine, D.E.; Pérez de León, A.A. Vitellogenin receptor as a target for tick control: A mini-review. Front. Physiol. 2019, 10, 618. [Google Scholar] [CrossRef] [PubMed]
- Hajdusek, O.; Sima, R.; Perner, J.; Loosova, G.; Harcubova, A.; Kopacek, P. Tick iron and heme metabolism—New target for an anti-tick intervention. Ticks Tick Borne Dis. 2016, 7, 565–572. [Google Scholar] [CrossRef]
- Galay, R.L.; Miyata, T.; Umemiya-Shirafuji, R.; Maeda, H.; Kusakisako, K.; Tsuji, N.; Mochizuki, M.; Fujisaki, K.; Tanaka, T. Evaluation and comparison of the potential of two ferritins as anti-tick vaccines against Haemaphysalis longicornis. Parasit. Vectors 2014, 7, 482. [Google Scholar] [CrossRef] [PubMed]
- Fogaça, A.C.; Sousa, G.; Pavanelo, D.B.; Esteves, E.; Martins, L.A.; Urbanová, V.; Kopáček, P.; Daffre, S. Tick immune system: What is known, the interconnections, the gaps, and the challenges. Front. Immunol. 2021, 12, 628054. [Google Scholar] [CrossRef]
- Luo, J.; Shen, H.; Ren, Q.; Guan, G.; Zhao, B.; Yin, H.; Chen, R.; Zhao, H.; Luo, J.; Li, X.; et al. Characterization of an MLP homologue from Haemaphysalis longicornis (Acari: Ixodidae) ticks. Pathogens. 2020, 9, 284. [Google Scholar] [CrossRef]
- Parizi, L.F.; Githaka, N.W.; Logullo, C.; Konnai, S.; Masuda, A.; Ohashi, K.; da Silva Vaz, I., Jr. The quest for a universal vaccine against ticks: Cross-immunity insights. Vet. J. 2012, 194, 158–165. [Google Scholar] [CrossRef]
- Trimnell, A.R.; Hails, R.S.; Nuttall, P.A. Dual action ectoparasite vaccine targeting ’exposed’ and ‘concealed’ antigens. Vaccine 2002, 20, 3560–3568. [Google Scholar] [CrossRef]
- Schultze, T.L.; Parkin, W.E.; Bosler, E.M. Vector tick populations and lyme disease. Ann. N. Y. Acad. Sci. 1988, 539, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Novikov, V.N.; Vshivkova, O.A.; Kruglik, O.V.; Morgulis, I.I.; Khlebopros, R.G. Elimination of tick populations from ecosystems. Doklady Biol. Sci. 2008, 420, 209–210. [Google Scholar] [CrossRef]
- Rosà, R.; Pugliese, A. Effects of tick population dynamics and host densities on the persistence of tick-borne infections. Math. Biosci. 2007, 208, 216–240. [Google Scholar] [CrossRef] [PubMed]
- Ogden, N.H.; Bigras-Poulin, M.; O’Callaghan, C.J.; Barker, I.K.; Lindsay, L.R.; Maarouf, A.; Smoyer-Tomic, K.E.; Waltner-Toews, D.; Charron, D. A dynamic population model to investigate effects of climate on geographic range and seasonality of the tick Ixodes scapularis. Int. J. Parasit. 2005, 35, 345–389. [Google Scholar] [CrossRef] [PubMed]
- Fan, G.; Thieme, H.R.; Zhu, H. Delay differential systems for tick population dynamics. J. Math. Biol. 2015, 71, 1017–1048. [Google Scholar] [CrossRef]
- Jennings, R.; Kuang, Y.; Thieme, H.R.; Wu, J.; Wu, X. How ticks keep ticking in the adversity of host immune reactions. J. Math. Biol. 2009, 78, 1331–1364. [Google Scholar] [CrossRef]
- Keesing, F.; Holt, R.D.; Ostfeld, R.S. Effects of species diversity on disease risk. Ecol. Lett. 2006, 9, 485–498. [Google Scholar] [CrossRef]
- Ostfeld, R.S. Biodiversity loss and the rise of zoonotic pathogens. Clin. Microbiol. Infect. 2009, 15, 40–433. [Google Scholar] [CrossRef]
- Barnett, K.; Civitello, D. Ecological and evolutionary challenges for wildlife vaccination. Trends Parasitol. 2020, 36, 970–978. [Google Scholar] [CrossRef]
- Kurtenbach, K.; Dizij, A.; Voet, P.; Hauser, P.; Simon, M.M. Vaccination of natural reservoir hosts with recombinant lapidated OspA induces a transmission-blocking immunity against Lyme disease spirochaetes associated with high levels of LA-2 equivalent antibodies. Vaccine 1997, 15, 1670–1674. [Google Scholar] [CrossRef]
- Tsao, J.I.; Wootton, J.T.; Bunikis, J.; Luna, M.G.; Fish, D.; Barbour, A.G. An ecological approach to preventing human infection: Vaccinating wild mouse reservoirs intervenes in the Lyme disease cycle. Proc. Natl. Acad. Sci. USA 2004, 101, 18159–18164. [Google Scholar] [CrossRef]
- Scheckelhoff, M.R.; Telford, S.R.; Hu, L.T. Protective efficacy of an oral vaccine to reduce carriage of Borrelia burgdorferi (strain N40) in mouse and tick reservoirs. Vaccine 2006, 24, 1949–1957. [Google Scholar] [CrossRef]
- Telford, S.R., 3rd; Cunningham, J.A.; Waltari, E.; Hu, L. Nest box-deployed bait for delivering oral vaccines to white-footed mice. Ticks Tick Borne Dis. 2011, 2, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, D.; Bensaci, M.; Luker, K.E.; Luker, G.; Wisdom, S.; Telford, S.R.; Hu, L.T. Development of a baited oral vaccine for use in reservoir-targeted strategies against Lyme disease. Vaccine 2011, 29, 7818–7825. [Google Scholar] [CrossRef] [PubMed]
- Meirelles Richer, L.; Aroso, M.; Contente-Cuomo, T.; Ivanova, L.; Gomes-Solecki, M. Reservoir targeted vaccine for lyme borreliosis induces a yearlong, neutralizing antibody response to OspA in white-footed mice. Clin. Vaccine Immunol. 2011, 18, 1809–1816. [Google Scholar] [CrossRef] [PubMed]
- Richer, L.M.; Brisson, D.; Melo, R.; Ostfeld, R.S.; Zeidner, N.; Gomes-Solecki, M. Reservoir targeted vaccine against Borrelia burgdorferi: A new strategy to prevent Lyme disease transmission. J. Infect. Dis. 2014, 209, 1972–1980. [Google Scholar] [CrossRef]


{kind=link}
| Observations | Publication | Reference Number |
|---|---|---|
| Cattle develop immunity to tick infestation in field | Hunt, 1899 | [41] |
| Cattle resistance developed to Dermacentor andersoni infestation associated with exudate that traps ticks | Bishopp and Wood, 1913 | [42] |
| Expression of bovine acquired resistance to tick infestation has a genetic component, especially for Bos indicus background | Johnston and Bancroft, 1918; Riek, 1962 | [43] [44] |
| Cutaneous Hypersensitivity Induced Grooming Response Observations | Publication | Reference Number |
|---|---|---|
| Bovine acquired resistance cutaneous hypersensitivity associated with exudate that traps ticks, bite site vesicles, poorly fed and dead ticks | Bishopp and Wood, 1913; Johnston and Bancroft, 1918; Riek, 1962 | [42] [43] [44] |
| Increased histamine levels; Mast cell degranulation at bite sites on resistant cattle; Basophil influx with ticks trapped in serous exudate | Riek, 1962 Schleger et al., 1976 Allen et al., 1977 | [44] [58] [59] |
| Grooming in response to tick induced pruritus is an important factor in expression of acquired resistance | Snowball, 1956 Bennett, 1969 de Castro and Newson, 1993 Hart, 2000 | [65] [67] [66] [69] |
| Human hypersensitivity to tick bite correlates with reduced incidence of tick-borne infections | Burke et al., 2005 | [70] |
| Observations | Publication | Reference Number |
|---|---|---|
| Basophils in acquired resistance to ticks | Karasuyama et al., 2018, 2020; Yoshikawa et al., 2021 | [107,120] [128] |
| Basophil and mast cell similarities and differences in their biology, roles in host defense and disease pathogenesis, and availability of specific molecular tools to distinguish their effector functions | Voehringer, 2013; Karasuyama et al., 2018, 2020 Tabakawa et al., 2018 Yoshikawa et al., 2021 | [106] [107,120] [125] [128] |
| Basophil function as antigen presenting cells for Th2 responses | Ohta et al., 2017 Karasuyama et al., 2018, 2020 | [126] [107,120] |
| Antigen Category | Tick Species | Antigen Types | References |
|---|---|---|---|
| Midgut, Reproductive Tissue, Malpighian Tubule | |||
| Dermacentor andersoni | Extracts of midgut and reproductive tissues | [172] | |
| Rhipicephalus (Boophilus) microplus | Bm 86, BM 86 combined with Bm 95 | [173,174,177] | |
| Rhipicephalus (Boophilus) annulatus, Hyalomma anatolicum anatolicum | Bm 86 homologues | [175,176] | |
| Rhipicephalus sanguineus, Rhipicephalus (B.) microplus | Ribosomal peptide, PO, combined with Bm 86 | [178] | |
| Rhipicephalus (B.) microplus. Rhipicephalus appendiculatus | Midgut and Malpighian tubule protein with epidermal growth factor domains | [179] | |
| Water Balance | |||
| Ixodes ricinus, Rhipicephalus (B.) microplus | Aquaporins with essential roles in blood feeding | [183,187] | |
| Rhipicephalus (B.) microplus, Rhipicephalus sanguineus, Ixodes ricinus, Ornithodoros erraticus | Aquaporins | [188,189,190,193] | |
| Tick Cell Signal Transduction | |||
| Multiple tick species | Subolesin widely conserved in ticks | [195] | |
| Rhipicephalus (Boophilus) australis | Bm 86 and subolesin | [198] | |
| Haemophysalis longicornis | Subolesin followed by subolesin-PO chimeric polypeptide | [199] | |
| Salivary Gland Proteins | |||
| Amblyomma sculptum | Protease inhibitors of factor Xa, trypsin, thrombin | [204] | |
| Rhipicephalus (B.) microplus | Metalloprotease, rBrRm-MP4 | [205] | |
| Rhipicephalus appendiculatus | Attachment cement protein, 64 P | [21,202] | |
| Ixodes scapularis | Salp 15 | [207,208] | |
| Dermacentor marginatus | Glutathione S-transferase | [217] | |
| Ixodes ricinus | Serine protease inhibitor | [218] | |
| Tick Microbiome | |||
| Multiple species | Multiple potential targets for disruption of tick physiological processes with microbiome differences among species and life cycle stages | [219,220,221,222,223,224,225] | |
| Ixodes ricinus | Microbiome Enterobacteriaceae | [34] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Oosterwijk, J.G.; Wikel, S.K. Resistance to Ticks and the Path to Anti-Tick and Transmission Blocking Vaccines. Vaccines 2021, 9, 725. https://doi.org/10.3390/vaccines9070725
van Oosterwijk JG, Wikel SK. Resistance to Ticks and the Path to Anti-Tick and Transmission Blocking Vaccines. Vaccines. 2021; 9(7):725. https://doi.org/10.3390/vaccines9070725
Chicago/Turabian Stylevan Oosterwijk, Jolieke G., and Stephen K. Wikel. 2021. "Resistance to Ticks and the Path to Anti-Tick and Transmission Blocking Vaccines" Vaccines 9, no. 7: 725. https://doi.org/10.3390/vaccines9070725
APA Stylevan Oosterwijk, J. G., & Wikel, S. K. (2021). Resistance to Ticks and the Path to Anti-Tick and Transmission Blocking Vaccines. Vaccines, 9(7), 725. https://doi.org/10.3390/vaccines9070725