
Vertebrate Responses against Arthropod Salivary Proteins and Their Therapeutic Potential

 ,
,  ,
, 
Abstract
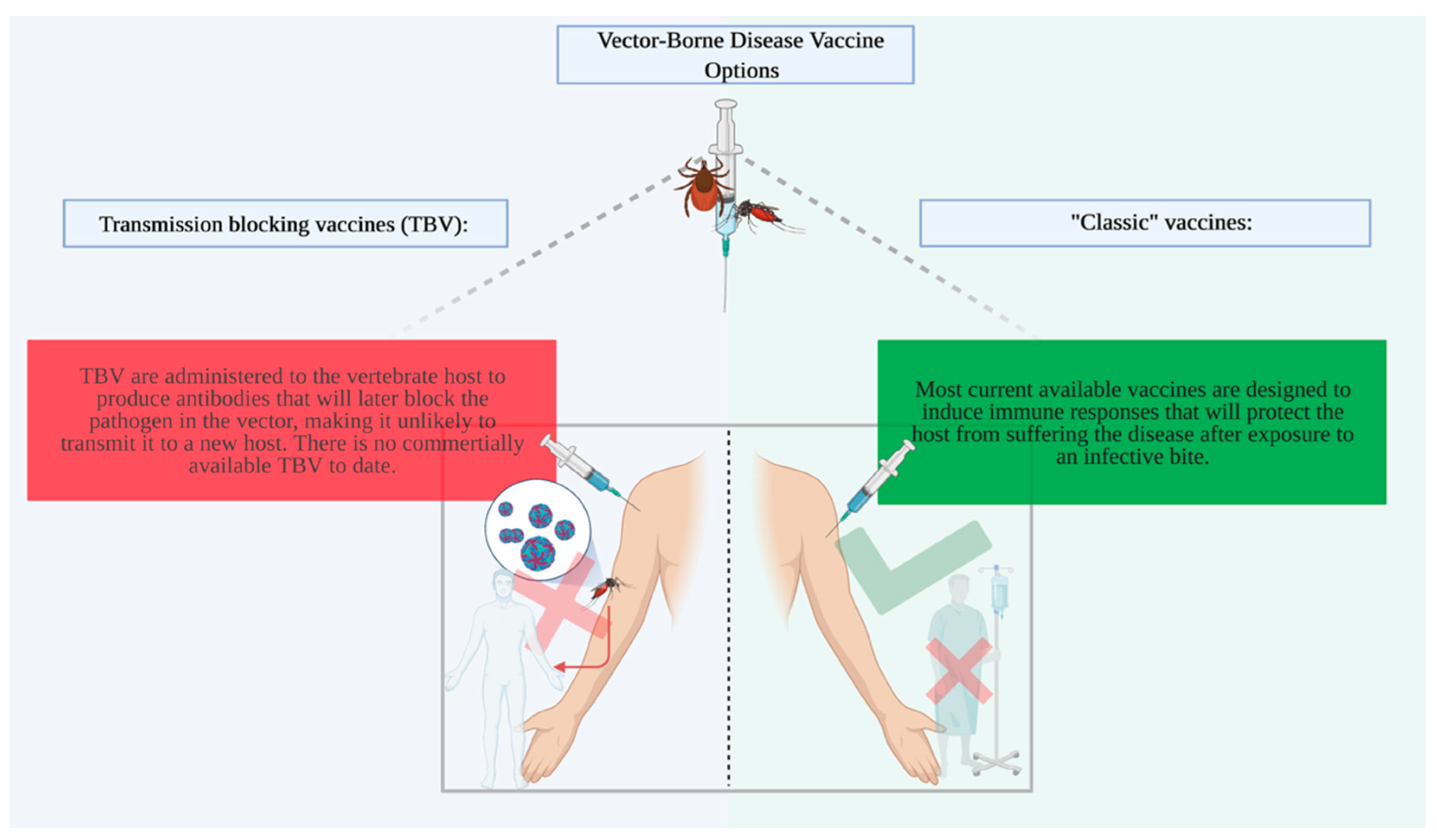
1. Introduction

{kind=link}
{kind=link}
| Pathogen | Vaccine Name | Year Licensed | Efficacy | Component | References |
|---|---|---|---|---|---|
| Dengue virus | Dengvaxia (CYT-TDV) | 2015 | 25–59% | Live attenuated tetravalent chimeric vaccine | [17] |
| Yellow fever virus | YF-VAX | 2001 | >95% | Live attenuated yellow fever virus strain 17D-204 | [11,18] |
| Yellow fever virus | STAMARIL | 1986 | Comparable to YF-VAX | Live attenuated yellow fever virus strain 17D-204 | [19] |
| Japanese Encephalitis virus | IXIARO/JESPECT | 2009 | 99.3% | Live attenuated SA-14-14-2 | [13,20] |
2. Arthropod Salivary Protein Candidates for Vaccines
3. Tick Salivary Proteins and Pathogen Transmission
| Pathogen | Protein | Species | Function | Phase | References |
|---|---|---|---|---|---|
| Tick Borne Encephalitis virus | 64TRP | Rhipicephalus appendiculatus | Disrupts the skin feeding site and then specific anti-64TRP antibodies cross-react with midgut antigenic epitopes. | Pre-clinic | [75,76] |
| Lyme disease | SALP15 | Ixodes scapularis | Inhibition of CD4+ T-cell activation by binding to the CD4 coreceptor of host T-cells, inhibiting receptor ligand-induced early cell signaling. | Pre-clinic | [58,77] |
| Lyme disease | SALP25D | Ixodes scapularis | Detoxified reactive oxygen species at the tick–bacteria–host interface that provides a survival advantage to B. burgdorferi. | Pre-clinic | [62,64] |
| Lyme disease | TSLP | Ixodes scapularis | Protects B. burgdorferi from the complement system. | Pre-clinic | [78] |
| Lyme disease | tHRF | Ixodes scapularis | Facilitates the transmission of Borrelia spp. to the mammalian host. | Pre-clinic | [64] |
| Babesiosis and Theileriosis | HqCRT | Haemaphysalis qinghaiensis | Induces host good humoral response against ticks feeding process. | Pre-clinic | [71,79] |
| Babesiosis and Theileriosis | HlCRT | Haemaphysalis longicornis | Induces host good humoral response against ticks salivary extracts. | Pre-clinic | [72] |
| Babesiosis and Theileriosis | rBmCRT | Rhipicephalus microplus | Induces host good humoral response against ticks salivary extracts. | Pre-clinic | [72] |
| Anaplasmosis | Sialostatin | Ixodes scapularis | Affects the formation of inflammasomes promoting host infection | .Pre-clinic | [80,81] |
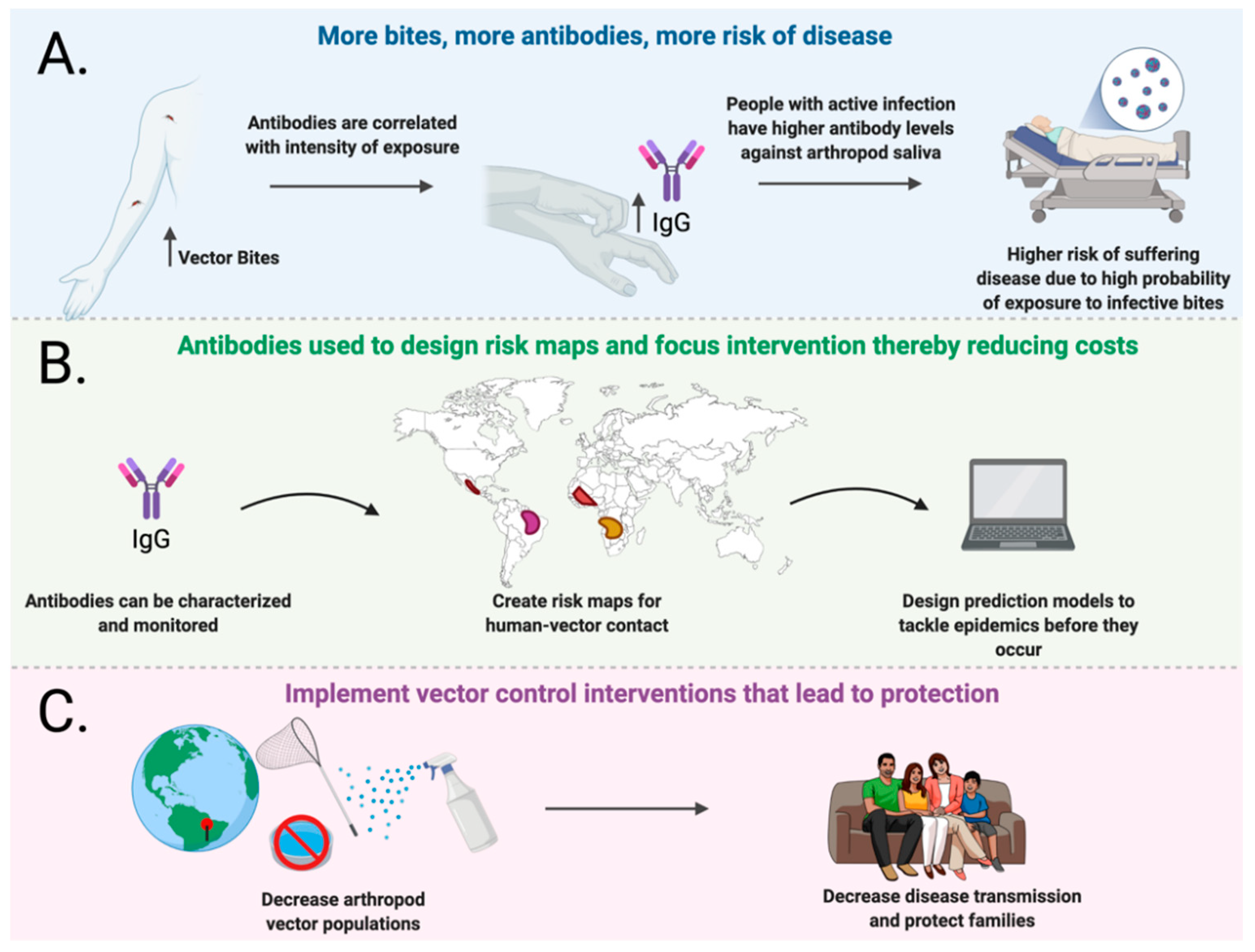
4. Natural Antibody Responses against Arthropod Salivary Proteins and Disease
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Global Vector Control Response 2017–2030; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- WHO. Dengue and Severe Dengue Fact Sheet; WHO: Geneva, Switzerlan, 2020. [Google Scholar]
- Bardach, A.E.; García-Perdomo, H.A.; Alcaraz, A.; López, E.T.; Gándara, R.A.R.; Ruvinsky, S.; Ciapponi, A. Interventions for the control of Aedes aegypti in Latin America and the Caribbean: Systematic review and meta-analysis. Trop. Med. Int. Health 2019, 24, 530–552. [Google Scholar] [CrossRef]
- Dhiman, S. Are malaria elimination efforts on right track? An analysis of gains achieved and challenges ahead. Infect. Dis. Poverty 2019, 8, 14. [Google Scholar] [CrossRef]
- Wilson, A.L.; Courtenay, O.; Kelly-Hope, L.A.; Scott, T.W.; Takken, W.; Torr, S.J.; Lindsay, S.W. The importance of vector control for the control and elimination of vector-borne diseases. PLoS Negl. Trop. Dis. 2020, 14, e0007831. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G.; Beier, J.C. Current vector control challenges in the fight against malaria. Acta Trop. 2017, 174, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Gari, T.; Lindtjørn, B. Reshaping the vector control strategy for malaria elimination in Ethiopia in the context of current evidence and new tools: Opportunities and challenges. Malar. J. 2018, 17, 454. [Google Scholar] [CrossRef]
- Bowman, L.R.; Donegan, S.; McCall, P.J. Is Dengue Vector Control Deficient in Effectiveness or Evidence: Systematic Review and Meta-analysis. PLoS Negl. Trop. Dis. 2016, 10, e0004551. [Google Scholar] [CrossRef] [PubMed]
- Bakhshi, H.; Failloux, A.-B.; Zakeri, S.; Raz, A.; Dinparast, N.D. Mosquito-borne viral diseases and potential transmission blocking vaccine candidates. Infect. Genet. Evol. 2018, 63, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Londono-Renteria, B.; Troupin, A.; Colpitts, T.M. Arbovirosis and potential transmission blocking vaccines. Parasites Vectors 2016, 9, 516. [Google Scholar] [CrossRef] [PubMed]
- Frierson, J.G. The Yellow Fever Vaccine: A History. Yale J. Biol. Med. 2010, 83, 77–85. [Google Scholar]
- Izmirly, A.M.; Alturki, S.O.; Alturki, S.O.; Connors, J.; Haddad, E.K. Challenges in Dengue Vaccines Development: Pre-existing Infections and Cross-Reactivity. Front. Immunol. 2020, 11, 1055. [Google Scholar] [CrossRef]
- Firbas, C.; Jilma, B. Product review on the JE vaccines IXIARO. Hum. Vaccines Immunother. 2015, 11, 411–420. [Google Scholar] [CrossRef]
- Hennessy, S.; Liu, Z.; Tsai, T.F.; Strom, B.L.; Wan, C.M.; Liu, L.H.; Wu, T.X.; Yu, H.J.; Liu, Q.M.; Karabatsos, N.; et al. Effectiveness of live-attenuated Japanese encephalitis vaccine (SA14-14-2): A case-control study. Lancet 1996, 347, 1583–1586. [Google Scholar] [CrossRef]
- Olson, D.; Rick, A.-M.; Krager, S.; Lamb, M.; Asturias, E.J. Vaccine Demand and Willingness-to-pay for Arbovirus Vaccines: A Cross-sectional Survey in Rural Guatemala. Pediatr. Infect. Dis. J. 2018, 37, 1184–1189. [Google Scholar] [CrossRef]
- Espinal, M.A.; Andrus, J.K.; Jauregui, B.; Waterman, S.H.; Morens, D.M.; Santos, J.I.; Horstick, O.; Francis, L.A.; Olson, D. Emerging and ReemergingAedes-Transmitted Arbovirus Infections in the Region of the Americas: Implications for Health Policy. Am. J. Public Health 2019, 109, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.J.; Yoon, I.-K. A review of Dengvaxia(R): Development to deployment. Hum. Vaccines Immunother. 2019, 15, 2295–2314. [Google Scholar] [CrossRef] [PubMed]
- Collins, N.D.; Barrett, A.D.T. Live Attenuated Yellow Fever 17D Vaccine: A Legacy Vaccine Still Controlling Outbreaks in Modern Day. Curr. Infect. Dis. Rep. 2017, 19, 14. [Google Scholar] [CrossRef] [PubMed]
- Cottin, P.; Niedrig, M.; Domingo, C. Safety profile of the yellow fever vaccine Stamaril(R): A 17-year review. Expert Rev. Vaccines 2013, 12, 1351–1368. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y. Phenotypic and genotypic characteristics of Japanese encephalitis attenuated live vaccine virus SA14-14-2 and their stabilities. Vaccine 2010, 28, 3635–3641. [Google Scholar] [CrossRef] [PubMed]
- Coutinho-Abreu, I.V.; Guimarães-Costa, A.B.; Valenzuela, J.G. Impact of insect salivary proteins in blood feeding, host immunity, disease, and in the development of biomarkers for vector exposure. Curr. Opin. Insect Sci. 2015, 10, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Vogt, M.B.; Lahon, A.; Arya, R.P.; Kneubehl, A.R.; Spencer Clinton, J.L.; Paust, S.; Rico-Hesse, R. Mosquito saliva alone has profound effects on the human immune system. PLoS Negl. Trop. Dis. 2018, 12, e0006439. [Google Scholar] [CrossRef]
- Calvo, E.; Mans, B.J.; Andersen, J.F.; Ribeiro, J.M. Function and Evolution of a Mosquito Salivary Protein Family. J. Biol. Chem. 2006, 281, 1935–1942. [Google Scholar] [CrossRef]
- Arcà, B.; Ribeiro, J.M. Saliva of hematophagous insects: A multifaceted toolkit. Curr. Opin. Insect Sci. 2018, 29, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Fong, S.-W.; Kini, R.M.; Ng, L.F.P. Mosquito Saliva Reshapes Alphavirus Infection and Immunopathogenesis. J. Virol. 2018, 92, e01004-17. [Google Scholar] [CrossRef] [PubMed]
- Styer, L.M.; Lim, P.Y.; Louie, K.L.; Albright, R.G.; Kramer, L.D.; Bernard, K.A. Mosquito Saliva Causes Enhancement of West Nile Virus Infection in Mice. J. Virol. 2011, 85, 1517–1527. [Google Scholar] [CrossRef] [PubMed]
- Pingen, M.; Bryden, S.R.; Pondeville, E.; Schnettler, E.; Kohl, A.; Merits, A.; Fazakerley, J.K.; Graham, G.J.; McKimmie, C.S. Host Inflammatory Response to Mosquito Bites Enhances the Severity of Arbovirus Infection. Immunity 2016, 44, 1455–1469. [Google Scholar] [CrossRef]
- Schneider, B.S.; Soong, L.; Coffey, L.L.; Stevenson, H.L.; McGee, C.E.; Higgs, S. Aedes aegypti Saliva Alters Leukocyte Recruitment and Cytokine Signaling by Antigen-Presenting Cells during West Nile Virus Infection. PLoS ONE 2010, 5, e11704. [Google Scholar] [CrossRef] [PubMed]
- Briant, L.; Desprès, P.; Choumet, V.; Missé, D. Role of skin immune cells on the host susceptibility to mosquito-borne viruses. Virology 2014, 464–465, 26–32. [Google Scholar] [CrossRef]
- Hopp, C.S.; Sinnis, P. The innate and adaptive response to mosquito saliva andPlasmodiumsporozoites in the skin. Ann. N. Y. Acad. Sci. 2015, 1342, 37–43. [Google Scholar] [CrossRef]
- Donovan, M.J.; Messmore, A.S.; Scrafford, D.A.; Sacks, D.L.; Kamhawi, S.; McDowell, M.A. Uninfected Mosquito Bites Confer Protection against Infection with Malaria Parasites. Infect. Immun. 2007, 75, 2523–2530. [Google Scholar] [CrossRef] [PubMed]
- Hastings, A.K.; Uraki, R.; Gaitsch, H.; Dhaliwal, K.; Stanley, S.; Sproch, H.; Williamson, E.; MacNeil, T.; Marin-Lopez, A.; Hwang, J.; et al. Aedes aegyptiNeSt1 Protein Enhances Zika Virus Pathogenesis by Activating Neutrophils. J. Virol. 2019, 93. [Google Scholar] [CrossRef]
- Uraki, R.; Hastings, A.K.; Brackney, D.E.; Armstrong, P.M.; Fikrig, E. AgBR1 antibodies delay lethal Aedes aegypti-borne West Nile virus infection in mice. NPJ Vaccines 2019, 4, 23. [Google Scholar] [CrossRef] [PubMed]
- Uraki, R.; Hastings, A.K.; Marin-Lopez, A.; Sumida, T.; Takahashi, T.; Grover, J.R.; Iwasaki, A.; Hafler, D.A.; Montgomery, R.R.; Fikrig, E. Aedes aegypti AgBR1 antibodies modulate early Zika virus infection of mice. Nat. Microbiol. 2019, 4, 948–955. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Guo, X.; Shen, C.; Hao, X.; Sun, P.; Li, P.; Xu, T.; Hu, C.; Rose, O.; Zhou, H.; et al. Salivary factor LTRIN from Aedes aegypti facilitates the transmission of Zika virus by interfering with the lymphotoxin-betaβ receptor. Nat. Immunol. 2018, 19, 342–353. [Google Scholar] [CrossRef] [PubMed]
- Conway, M.J.; Watson, A.M.; Colpitts, T.M.; Dragovic, S.M.; Li, Z.; Wang, P.; Feitosa, F.; Shepherd, D.T.; Ryman, K.D.; Klimstra, W.B.; et al. Mosquito Saliva Serine Protease Enhances Dissemination of Dengue Virus into the Mammalian Host. J. Virol. 2014, 88, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Conway, M.J.; Londono-Renteria, B.; Troupin, A.; Watson, A.M.; Klimstra, W.B.; Fikrig, E.; Colpitts, T.M. Aedes aegypti D7 Saliva Protein Inhibits Dengue Virus Infection. PLoS Negl. Trop. Dis. 2016, 10, e0004941. [Google Scholar] [CrossRef] [PubMed]
- Chagas, A.C.; Ramirez, J.L.; Jasinskiene, N.; James, A.A.; Ribeiro, J.M.; Marinotti, O.; Calvo, E. Collagen-binding protein, Aegyptin, regulates probing time and blood feeding success in the dengue vector mosquito, Aedes aegypti. Proc. Natl. Acad. Sci. USA 2014, 111, 6946–6951. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Nie, K.; Zhu, Y.; Liu, Y.; Wu, P.; Liu, Z.; Du, S.; Fan, H.; Chen, C.-H.; Zhang, R.; et al. A mosquito salivary protein promotes flavivirus transmission by activation of autophagy. Nat. Commun. 2020, 11, 260. [Google Scholar] [CrossRef]
- Manning, J.E.; Oliveira, F.; Coutinho-Abreu, I.V.; Herbert, S.; Meneses, C.; Kamhawi, S.; Baus, H.A.; Han, A.; Czajkowski, L.; Rosas, L.A.; et al. Safety and immunogenicity of a mosquito saliva peptide-based vaccine: A randomised, placebo-controlled, double-blind, phase 1 trial. Lancet 2020, 395, 1998–2007. [Google Scholar] [CrossRef]
- Manning, J.E.; Morens, D.M.; Kamhawi, S.; Valenzuela, J.G.; Memoli, M. Mosquito Saliva: The Hope for a Universal Arbovirus Vaccine? J. Infect. Dis. 2018, 218, 7–15. [Google Scholar] [CrossRef]
- Wang, Y.; Marin-Lopez, A.; Jiang, J.; Ledizet, M.; Fikrig, E. Vaccination with Aedes aegypti AgBR1 Delays Lethal Mosquito-Borne Zika Virus Infection in Mice. Vaccines 2020, 8, 145. [Google Scholar] [CrossRef]
- Surasombatpattana, P.; Ekchariyawat, P.; Hamel, R.; Patramool, S.; Thongrungkiat, S.; Denizot, M.; Delaunay, P.; Thomas, F.; Luplertlop, N.; Yssel, H.; et al. Aedes aegypti Saliva Contains a Prominent 34-kDa Protein that Strongly Enhances Dengue Virus Replication in Human Keratinocytes. J. Investig. Dermatol. 2014, 134, 281–284. [Google Scholar] [CrossRef] [PubMed]
- Elanga Ndille, E.; Doucoure, S.; Poinsignon, A.; Mouchet, F.; Cornelie, S.; D’Ortenzio, E.; Dehecq, J.S.; Remoue, F. Human IgG Antibody Response to Aedes Nterm-34kDa Salivary Peptide, an Epidemiological Tool to Assess Vector Control in Chikungunya and Dengue Transmission Area. PLoS Negl. Trop. Dis. 2016, 10, e0005109. [Google Scholar] [CrossRef]
- Elanga Ndille, E.; Doucoure, S.; Damien, G.; Mouchet, F.; Drame, P.M.; Cornelie, S.; Noukpo, H.; Yamadjako, S.; Djenontin, A.; Moiroux, N.; et al. First Attempt To Validate Human IgG Antibody Response to Nterm-34kDa Salivary Peptide as Biomarker for Evaluating Exposure to Aedes aegypti Bites. PLoS Negl. Trop. Dis. 2012, 6, e1905. [Google Scholar] [CrossRef] [PubMed]
- Nuttall, P. Vaccinating against mosquitoes: Anticipating the unexpected. Lancet 2020, 395, 1953–1954. [Google Scholar] [CrossRef]
- Jablonka, W.; Kim, I.H.; Alvarenga, P.H.; Valenzuela, J.G.; Ribeiro, J.M.C.; Andersen, J.F. Functional and structural similarities of D7 proteins in the independently-evolved salivary secretions of sand flies and mosquitoes. Sci. Rep. 2019, 9, 5340. [Google Scholar] [CrossRef]
- Valenzuela, J.G.; Charlab, R.; Gonzalez, E.C.; De Miranda-Santos, I.K.; Marinotti, O.; Francischetti, I.M.; Ribeiro, J.M. The D7 family of salivary proteins in blood sucking diptera. Insect Mol. Biol. 2002, 11, 149–155. [Google Scholar] [CrossRef]
- Martin-Martin, I.; Paige, A.; Valenzuela Leon, P.C.; Gittis, A.G.; Kern, O.; Bonilla, B.; Chagas, A.C.; Ganesan, S.; Smith, L.B.; Garboczi, D.N.; et al. ADP binding by the Culex quinquefasciatus mosquito D7 salivary protein enhances blood feeding on mammals. Nat. Commun. 2020, 11, 2911. [Google Scholar] [CrossRef]
- Londono-Renteria, B.L.; Shakeri, H.; Rozo-Lopez, P.; Conway, M.J.; Duggan, N.; Jaberi-Douraki, M.; Colpitts, T.M. Serosurvey of Human Antibodies Recognizing Aedes aegypti D7 Salivary Proteins in Colombia. Front. Public Health 2018, 6, 111. [Google Scholar] [CrossRef] [PubMed]
- Reagan, K.L.; Machain-Williams, C.; Wang, T.; Blair, C.D. Immunization of Mice with Recombinant Mosquito Salivary Protein D7 Enhances Mortality from Subsequent West Nile Virus Infection via Mosquito Bite. PLoS Negl. Trop. Dis. 2012, 6, e1935. [Google Scholar] [CrossRef]
- Sankar, S.; Ramamurthy, M.; Nandagopal, B.; Sridharan, G. In Silico Validation of D7 Salivary Protein-derived B- and T-cell Epitopes of Aedes aegypti as Potential Vaccine to Prevent Transmission of Flaviviruses and Togaviruses to Humans. Bioinformation 2017, 13, 366–375. [Google Scholar] [CrossRef]
- McCracken, M.K.; Christofferson, R.C.; Grasperge, B.J.; Calvo, E.; Chisenhall, D.M.; Mores, C.N. Aedes aegypti salivary protein “aegyptin” co-inoculation modulates dengue virus infection in the vertebrate host. Virology 2014, 468–470, 133–139. [Google Scholar] [CrossRef]
- Chisenhall, D.M.; Christofferson, R.C.; McCracken, M.K.; Johnson, A.M.; Londono-Renteria, B.; Mores, C.N. Infection with dengue-2 virus alters proteins in naturally expectorated saliva of Aedes aegypti mosquitoes. Parasites Vectors 2014, 7, 252. [Google Scholar] [CrossRef] [PubMed]
- Chisenhall, D.M.; Londono, B.L.; Christofferson, R.C.; McCracken, M.K.; Mores, C.N. Effect of Dengue-2 Virus Infection on Protein Expression in the Salivary Glands of Aedes aegypti Mosquitoes. Am. J. Trop. Med. Hyg. 2014, 90, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Piesman, J.; Gern, L. Lyme borreliosis in Europe and North America. Parasitology 2004, 129, S191–S220. [Google Scholar] [CrossRef] [PubMed]
- Anguita, J.; Ramamoorthi, N.; Hovius, J.W.; Das, S.; Thomas, V.; Persinski, R.; Conze, D.; Askenase, P.W.; Rincón, M.; Kantor, F.S.; et al. Salp15, an Ixodes scapularis Salivary Protein, Inhibits CD4+ T Cell Activation. Immunity 2002, 16, 849–859. [Google Scholar] [CrossRef]
- Ramamoorthi, N.; Narasimhan, S.; Pal, U.; Bao, F.; Yang, X.F.; Fish, D.; Anguita, J.; Norgard, M.V.; Kantor, F.S.; Anderson, J.F.; et al. The Lyme disease agent exploits a tick protein to infect the mammalian host. Nature 2005, 436, 573–577. [Google Scholar] [CrossRef]
- Rosa, P. Lyme disease agent borrows a practical coat. Nat. Med. 2005, 11, 831–832. [Google Scholar] [CrossRef]
- Dai, J.; Wang, P.; Adusumilli, S.; Booth, C.J.; Narasimhan, S.; Anguita, J.; Fikrig, E. Antibodies against a Tick Protein, Salp15, Protect Mice from the Lyme Disease Agent. Cell Host Microbe 2009, 6, 482–492. [Google Scholar] [CrossRef]
- Das, S.; Banerjee, G.; DePonte, K.; Marcantonio, N.; Kantor, F.S.; Fikrig, E. Salp25D, anIxodes scapularisAntioxidant, is 1 of 14 Immunodominant Antigens in Engorged Tick Salivary Glands. J. Infect. Dis. 2001, 184, 1056–1064. [Google Scholar] [CrossRef]
- Narasimhan, S.; Sukumaran, B.; Bozdogan, U.; Thomas, V.; Liang, X.; DePonte, K.; Marcantonio, N.; Koski, R.A.; Anderson, J.F.; Kantor, F.; et al. A Tick Antioxidant Facilitates the Lyme Disease Agent’s Successful Migration from the Mammalian Host to the Arthropod Vector. Cell Host Microbe 2007, 2, 7–18. [Google Scholar] [CrossRef]
- Wagemakers, A.; Coumou, J.; Schuijt, T.J.; Oei, A.; Nijhof, A.M.; van’t Veer, C.; van der Poll, T.; Bins, A.D.; Hovius, J.W. An Ixodes ricinus Tick Salivary Lectin Pathway Inhibitor Protects Borrelia burgdorferi sensu lato from Human Complement. Vector Borne Zoonotic Dis. 2016, 16, 223–228. [Google Scholar] [CrossRef]
- Dai, J.; Narasimhan, S.; Zhang, L.; Liu, L.; Wang, P.; Fikrig, E. Tick Histamine Release Factor Is Critical for Ixodes scapularis Engorgement and Transmission of the Lyme Disease Agent. PLoS Pathog. 2010, 6, e1001205. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, C.; Lynn, G.E.; Pedra, J.H.F.; Pal, U.; Narasimhan, S.; Fikrig, E. Interactions between Borrelia burgdorferi and ticks. Nat. Rev. Microbiol. 2020, 18, 587–600. [Google Scholar] [CrossRef] [PubMed]
- Tabor, A.E.; Ali, A.; Rehman, G.; Rocha Garcia, G.; Zangirolamo, A.F.; Malardo, T.; Jonsson, N.N. Cattle Tick Rhipicephalus microplus-Host Interface: A Review of Resistant and Susceptible Host Responses. Front. Cell. Infect. Microbiol. 2017, 7, 506. [Google Scholar] [CrossRef]
- Maruyama, S.R.; Garcia, G.R.; Teixeira, F.R.; Brandão, L.G.; Anderson, J.M.; Ribeiro, J.M.C.; Valenzuela, J.G.; Horackova, J.; Veríssimo, C.J.; Katiki, L.M.; et al. Mining a differential sialotranscriptome of Rhipicephalus microplus guides antigen discovery to formulate a vaccine that reduces tick infestations. Parasites Vectors 2017, 10, 206. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, S.R.; Carvalho, B.; González-Porta, M.; Rung, J.; Brazma, A.; Gustavo Gardinassi, L.; Ferreira, B.R.; Banin, T.M.; Veríssimo, C.J.; Katiki, L.M.; et al. Blood transcriptome profile induced by an efficacious vaccine formulated with salivary antigens from cattle ticks. NPJ Vaccines 2019, 4, 53. [Google Scholar] [CrossRef]
- Gold, L.I.; Rahman, M.; Blechman, K.M.; Greives, M.R.; Churgin, S.; Michaels, J.; Callaghan, M.J.; Cardwell, N.L.; Pollins, A.C.; Michalak, M.; et al. Overview of the Role for Calreticulin in the Enhancement of Wound Healing through Multiple Biological Effects. J. Investig. Dermatol. Symp. Proc. 2006, 11, 57–65. [Google Scholar] [CrossRef]
- Cabezón, C.; Cabrera, G.; Paredes, R.; Ferreira, A.; Galanti, N. Echinococcus granulosus calreticulin: Molecular characterization and hydatid cyst localization. Mol. Immunol. 2008, 45, 1431–1438. [Google Scholar] [CrossRef]
- Gao, J.; Luo, J.; Fan, R.; Fingerle, V.; Guan, G.; Liu, Z.; Li, Y.; Zhao, H.; Ma, M.; Liu, J.; et al. Cloning and characterization of a cDNA clone encoding calreticulin from Haemaphysalis qinghaiensis (Acari: Ixodidae). Parasitol. Res. 2008, 102, 737–746. [Google Scholar] [CrossRef]
- Parizi, L.F.; Rech, H.; Ferreira, C.A.; Imamura, S.; Ohashi, K.; Onuma, M.; Masuda, A.; Vaz Ida, S., Jr. Comparative immunogenicity of Haemaphysalis longicornis and Rhipicephalus (Boophilus) microplus calreticulins. Vet. Parasitol. 2009, 164, 282–290. [Google Scholar] [CrossRef]
- Kunz, C.; Hofmann, H.; Heinz, F.X.; Dippe, H. Efficacy of Vaccination Against Tick-Borne Encephalitis. Wien. Klin. Wochenschr. 1980, 92, 809–813. [Google Scholar] [PubMed]
- Rego, R.O.M.; Trentelman, J.J.A.; Anguita, J.; Nijhof, A.M.; Sprong, H.; Klempa, B.; Hajdusek, O.; Tomás-Cortázar, J.; Azagi, T.; Strnad, M.; et al. Counterattacking the tick bite: Towards a rational design of anti-tick vaccines targeting pathogen transmission. Parasites Vectors 2019, 12, 229. [Google Scholar] [CrossRef]
- Labuda, M.; Trimnell, A.R.; Ličková, M.; Kazimírová, M.; Davies, G.M.; Lissina, O.; Hails, R.S.; Nuttall, P.A. An Antivector Vaccine Protects against a Lethal Vector-Borne Pathogen. PLoS Pathog. 2006, 2, e27. [Google Scholar] [CrossRef]
- Trimnell, A.R.; Davies, G.M.; Lissina, O.; Hails, R.S.; Nuttall, P.A. A cross-reactive tick cement antigen is a candidate broad-spectrum tick vaccine. Vaccine 2005, 23, 4329–4341. [Google Scholar] [CrossRef] [PubMed]
- Garg, R.; Juncadella, I.J.; Ramamoorthi, N.; Ananthanarayanan, S.K.; Thomas, V.; Rincón, M.; Anguita, J. Cutting edge: CD4 is the receptor for the tick saliva immunosuppressor, Salp15. J. Immunol. 2006, 177, 6579–6583. [Google Scholar] [CrossRef] [PubMed]
- Schuijt, T.J.; Coumou, J.; Narasimhan, S.; Dai, J.; DePonte, K.; Wouters, D.; Fikrig, E. A Tick Mannose-Binding Lectin Inhibitor Interferes with the Vertebrate Complement Cascade to Enhance Transmission of the Lyme Disease Agent. Cell Host Microbe 2011, 10, 136–146. [Google Scholar] [CrossRef]
- Cotté, V.; Sabatier, L.; Schnell, G.; Carmi-Leroy, A.; Rousselle, J.C.; Arsène-Ploetze, F.; Choumet, V. Differential expression of Ixodes ricinus salivary gland proteins in the presence of the Borrelia burgdorferi sensu lato complex. J. Proteom. 2014, 96, 29–43. [Google Scholar] [CrossRef]
- Kotsyfakis, M.; Sá-Nunes, A.; Francischetti, I.M.; Mather, T.N.; Andersen, J.F.; Ribeiro, J.M. Antiinflammatory and Immunosuppressive Activity of Sialostatin L, a Salivary Cystatin from the Tick Ixodes scapularis. J. Biol. Chem. 2006, 281, 26298–26307. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Shaw, D.K.; Sakhon, O.S.; Snyder, G.A.; Sundberg, E.J.; Santambrogio, L.; Pedra, J.H. The Tick Protein Sialostatin L2 Binds to Annexin A2 and Inhibits NLRC4-Mediated Inflammasome Activation. Infect. Immun. 2016, 84, 1796–1805. [Google Scholar] [CrossRef]
- Penneys, N.S.; Nayar, J.K.; Bernstein, H.; Knight, J.W. Circulating antibody detection in human serum to mosquito salivary gland proteins by the avidin-biotin-peroxidase technique. J. Am. Acad. Dermatol. 1988, 18, 87–92. [Google Scholar] [CrossRef]
- Fontaine, A.; Pascual, A.; Orlandi-Pradines, E.; Diouf, I.; Remoué, F.; Pagès, F.; Fusaï, T.; Rogier, C.; Almeras, L. Relationship between Exposure to Vector Bites and Antibody Responses to Mosquito Salivary Gland Extracts. PLoS ONE 2011, 6, e29107. [Google Scholar] [CrossRef] [PubMed]
- Cardenas, J.C.; Drame, P.M.; Luque-Burgos, K.A.; Berrio, J.D.; Entrena-Mutis, E.; González, M.U.; Carvajal, D.J.; Gutiérrez-Silva, L.Y.; Cardenas, L.D.; Colpitts, T.M.; et al. IgG1 and IgG4 antibodies against Aedes aegypti salivary proteins and risk for dengue infections. PLoS ONE 2019, 14, e0208455. [Google Scholar] [CrossRef] [PubMed]
- Brummer-Korvenkontio, H.; Lappalainen, P.; Reunala, T.; Palosuo, T. Detection of mosquito saliva–specific IgE and IgG4 antibodies by immunoblotting. J. Allergy Clin. Immunol. 1994, 93, 551–555. [Google Scholar] [CrossRef]
- Reunala, T.; Brummer-Korvenkontio, H.; Palosuo, K.; Miyanij, M.; Ruiz-Maldonado, R.; Löve, A.; François, G.; Palosuo, T. Frequent Occurrence of IgE and lgG4 Antibodies against Saliva of Aedes communis and Aedes aegypti Mosquitoes in Children. Int. Arch. Allergy Immunol. 1994, 104, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Maldonado-Ruiz, L.P.; Montenegro-Cadena, L.; Blattner, B.; Menghwar, S.; Zurek, L.; Londono-Renteria, B. Differential Tick Salivary Protein Profiles and Human Immune Responses to Lone Star Ticks (Amblyomma americanum) From the Wild vs. a Laboratory Colony. Front. Immunol. 2019, 10, 1996. [Google Scholar] [CrossRef] [PubMed]
- Londono-Renteria, B.; Cardenas, J.C.; Giovanni, J.E.; Cardenas, L.; Villamizar, P.; Rolon, J.; Chisenhall, D.M.; Christofferson, R.C.; Carvajal, D.J.; Perez, O.G.; et al. Aedes aegypti anti-salivary gland antibody concentration and dengue virus exposure history in healthy individuals living in an endemic area in Colombia. Biomédica 2015, 35, 572–581. [Google Scholar] [CrossRef] [PubMed]
- Londono-Renteria, B.; Cardenas, J.C.; Cardenas, L.D.; Christofferson, R.C.; Chisenhall, D.M.; Wesson, D.M.; McCracken, M.K.; Carvajal, D.; Mores, C.N. Use of Anti-Aedes aegypti Salivary Extract Antibody Concentration to Correlate Risk of Vector Exposure and Dengue Transmission Risk in Colombia. PLoS ONE 2013, 8, e81211. [Google Scholar] [CrossRef]
- Londono-Renteria, B.L.; Eisele, T.P.; Keating, J.; James, M.A.; Wesson, D.M. Antibody Response Against Anopheles albimanus (Diptera: Culicidae) Salivary Protein as a Measure of Mosquito Bite Exposure in Haiti. J. Med. Èntomol. 2010, 47, 1156–1163. [Google Scholar] [CrossRef]
- Rizzo, C.; Lombardo, F.; Ronca, R.; Mangano, V.; Sirima, S.B.; Nèbiè, I.; Fiorentino, G.; Modiano, D.; Arcà, B. Differential antibody response to the Anopheles gambiae gSG6 and cE5 salivary proteins in individuals naturally exposed to bites of malaria vectors. Parasites Vectors 2014, 7, 549. [Google Scholar] [CrossRef]
- Ndille, E.E.; Dubot-Pérès, A.; Doucoure, S.; Mouchet, F.; Cornelie, S.; Sidavong, B.; Fournet, F.; Remoue, F. Human IgG antibody response toAedes aegyptiNterm-34 kDa salivary peptide as an indicator to identify areas at high risk for dengue transmission: A retrospective study in urban settings of Vientiane city, Lao PDR. Trop. Med. Int. Health 2014, 19, 576–580. [Google Scholar] [CrossRef]
- Buezo Montero, S.; Gabrieli, P.; Montarsi, F.; Borean, A.; Capelli, S.; De Silvestro, G.; Forneris, F.; Pombi, M.; Breda, A.; Capelli, G.; et al. IgG Antibody Responses to the Aedes albopictus 34k2 Salivary Protein as Novel Candidate Marker of Human Exposure to the Tiger Mosquito. Front. Cell. Infect. Microbiol. 2020, 10, 377. [Google Scholar] [CrossRef] [PubMed]
- Vannavong, N.; Seidu, R.; Stenström, T.-A.; Dada, N.; Overgaard, H.J. Effects of socio-demographic characteristics and household water management on Aedes aegypti production in suburban and rural villages in Laos and Thailand. Parasites Vectors 2017, 10, 170. [Google Scholar] [CrossRef] [PubMed]


| Pathogen | Protein | Species | Function | Phase | References |
|---|---|---|---|---|---|
| Zika virus | NeST1 | Aedes aegypti | Prevents early changes in inflammatory milieu. | Pre-clinic | [32] |
| Zika virus | AgBR1 | Aedes aegypti | Prevents early changes in inflammatory milieu. | Pre-clinic | [33,34] |
| Zika virus | LTRIN | Aedes aegypti | Binds and inhibits the lymphotoxin-β receptor (LTβR) | Pre-clinic | [35] |
| Dengue virus | CLIPA3 | Aedes aegypti | Disrupts extracellular matrix allowing virus dissemination | Pre-clinic | [36] |
| Dengue virus | D7 | Aedes aegypti | Inhibits DENV infection in vitro and in vivo. | Pre-clinic | [37] |
| Dengue virus | Aegyptin | Aedes aegypti | Blocks collagen-induced platelet aggregation | Pre-clinic | [38] |
| Dengue and Zika virus | AaVA-1 | Aedes aegypti | Increases viral replication in macrophages and dendritic cells. | Pre-clinic | [39] |
| Mosquito transmitted diseases | AGS-v | Anopheles gambiae | Increases vaccine-specific IgG antibodies and cellular responses | Phase 1 | [40] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olajiga, O.; Holguin-Rocha, A.F.; Rippee-Brooks, M.; Eppler, M.; Harris, S.L.; Londono-Renteria, B. Vertebrate Responses against Arthropod Salivary Proteins and Their Therapeutic Potential. Vaccines 2021, 9, 347. https://doi.org/10.3390/vaccines9040347
Olajiga O, Holguin-Rocha AF, Rippee-Brooks M, Eppler M, Harris SL, Londono-Renteria B. Vertebrate Responses against Arthropod Salivary Proteins and Their Therapeutic Potential. Vaccines. 2021; 9(4):347. https://doi.org/10.3390/vaccines9040347
Chicago/Turabian StyleOlajiga, Olayinka, Andrés F. Holguin-Rocha, Meagan Rippee-Brooks, Megan Eppler, Shanice L. Harris, and Berlin Londono-Renteria. 2021. "Vertebrate Responses against Arthropod Salivary Proteins and Their Therapeutic Potential" Vaccines 9, no. 4: 347. https://doi.org/10.3390/vaccines9040347
APA StyleOlajiga, O., Holguin-Rocha, A. F., Rippee-Brooks, M., Eppler, M., Harris, S. L., & Londono-Renteria, B. (2021). Vertebrate Responses against Arthropod Salivary Proteins and Their Therapeutic Potential. Vaccines, 9(4), 347. https://doi.org/10.3390/vaccines9040347