Xanthohumol and Gossypol Are Promising Inhibitors against Babesia microti by In Vitro Culture via High-Throughput Screening of 133 Natural Products

Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Parasites and Mice
2.3. Preparation of Infected Erythrocytes
2.4. In Vitro Culture of B. microti
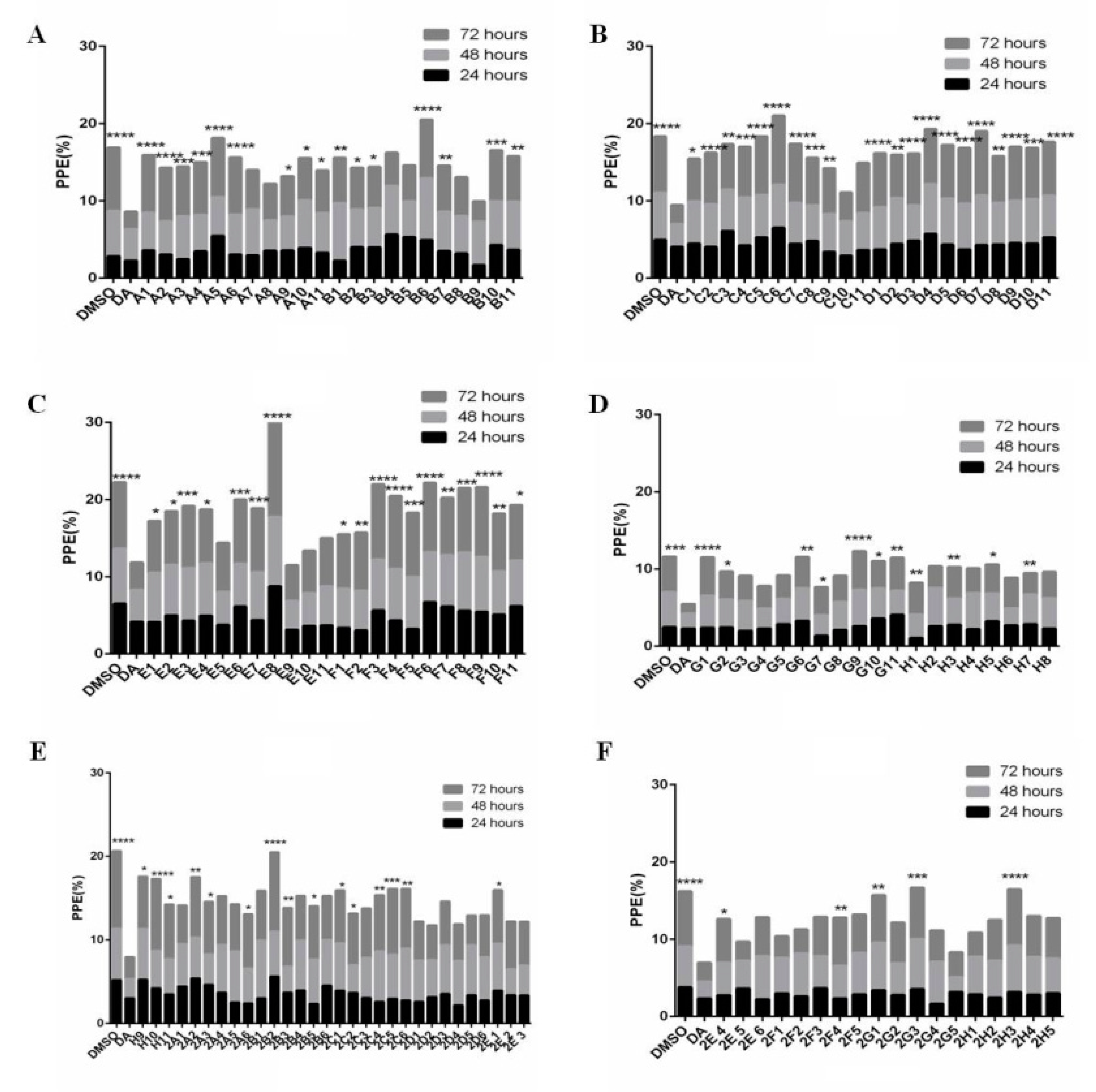
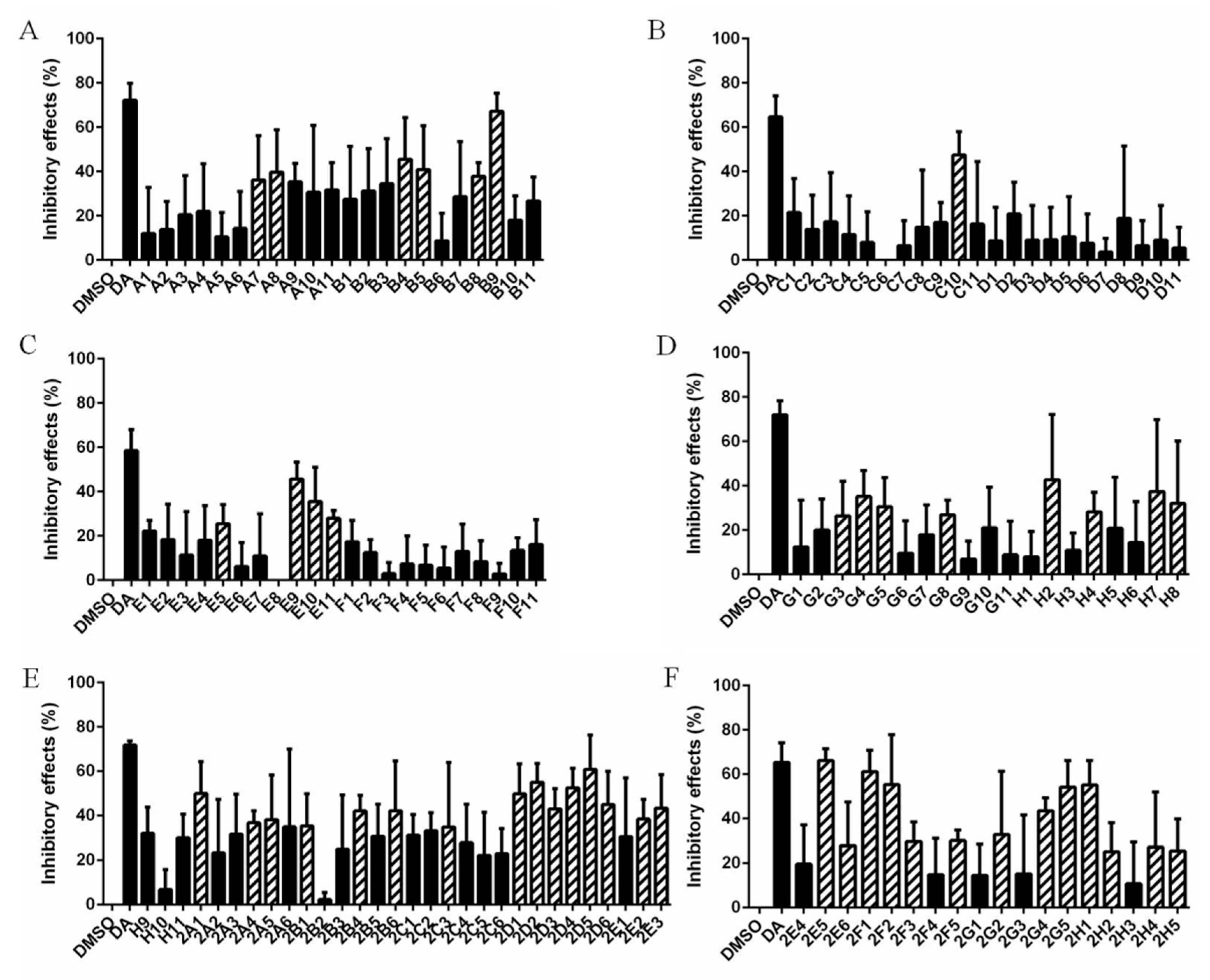
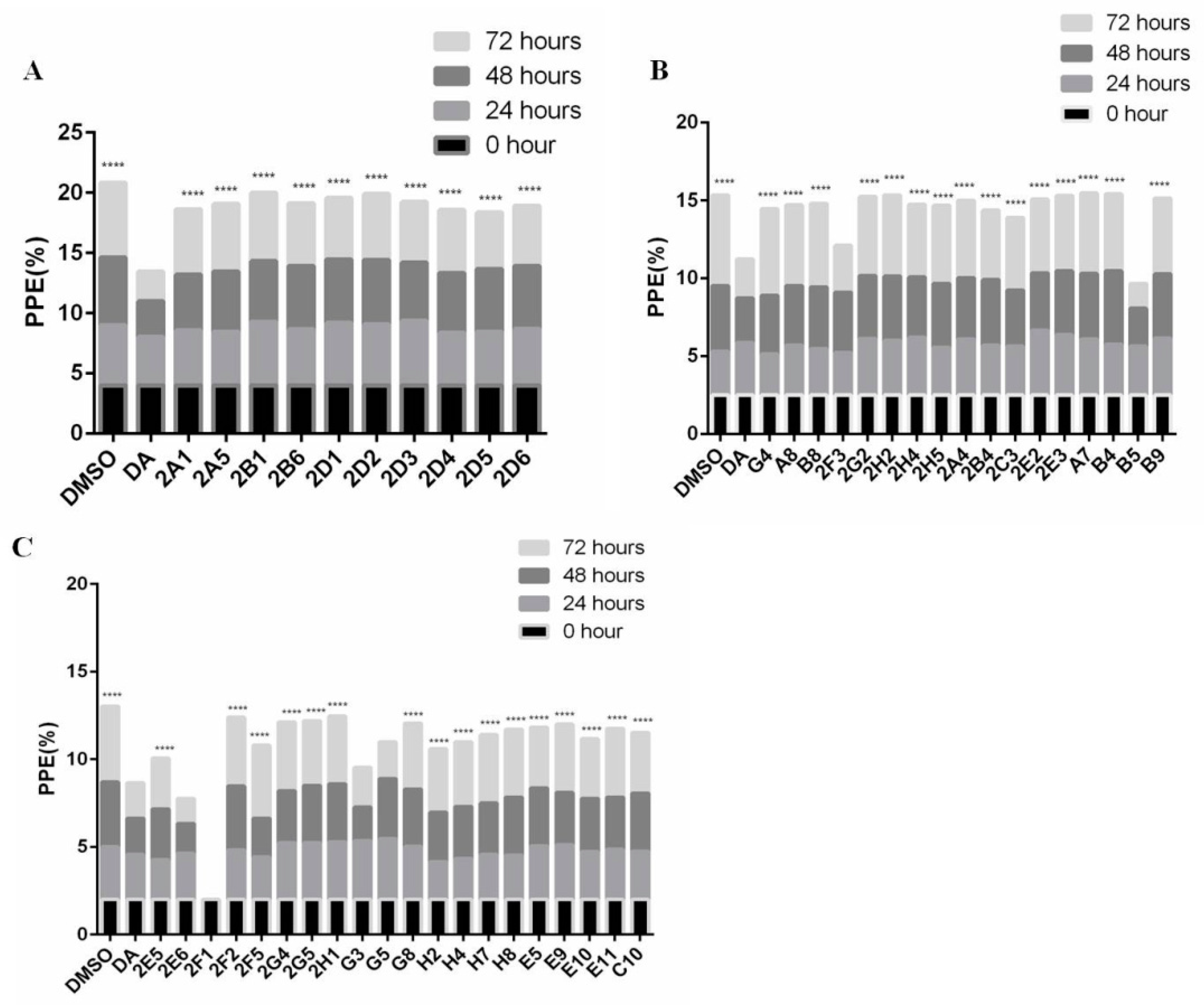
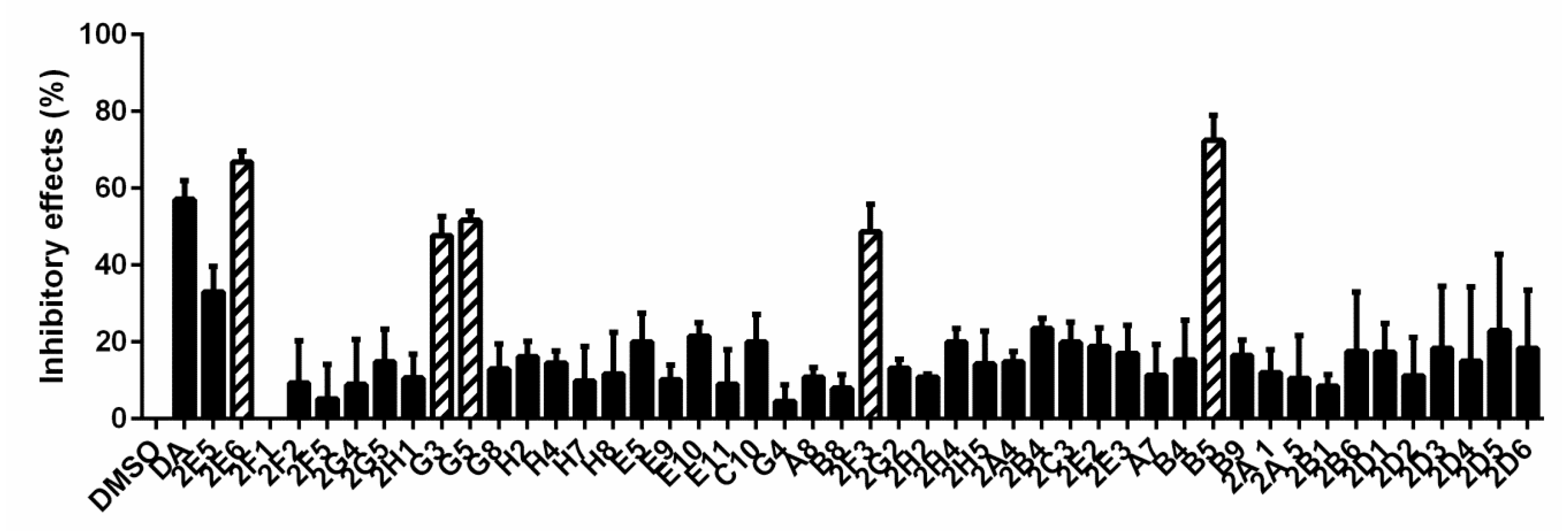
2.5. Preliminary Screening and Rescreening of Natural Products
2.6. Combination of Gp and Xn on In Vitro Culture
2.7. Cell Toxicity Assay
2.8. In Vivo Inhibition Assay
3. Results
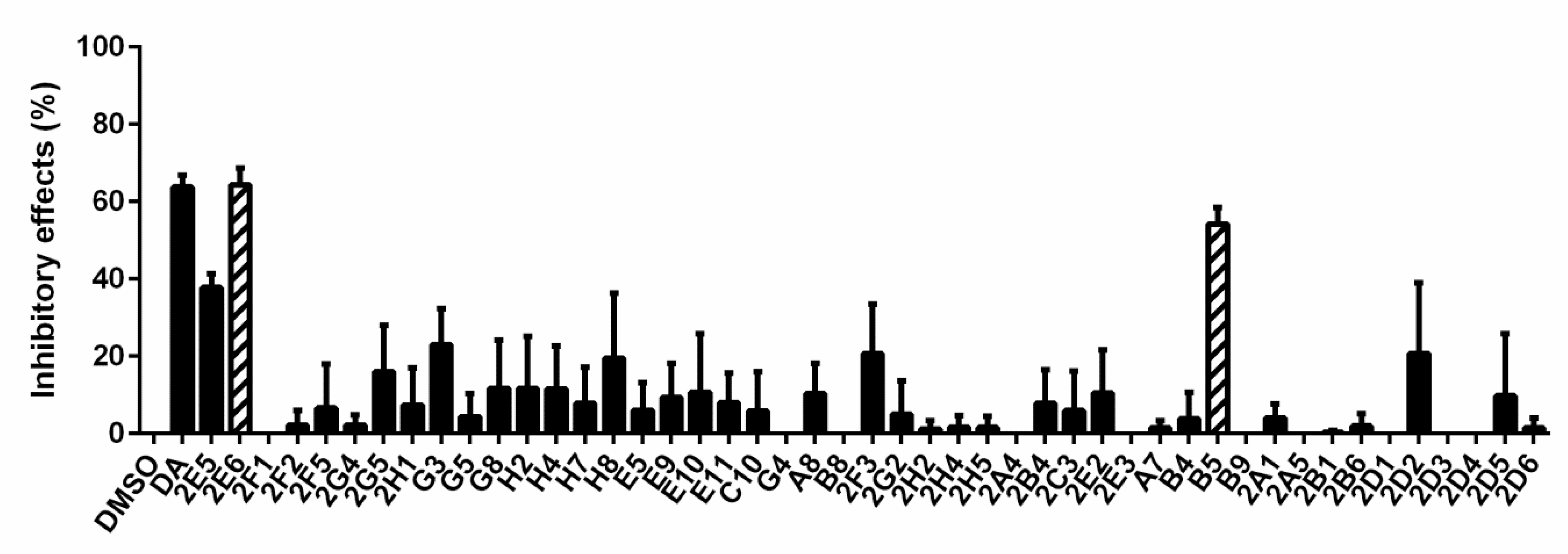
3.1. Forty-Seven Natural Products Showed Significant Inhibitory Effects on B. microti in Preliminary Screening
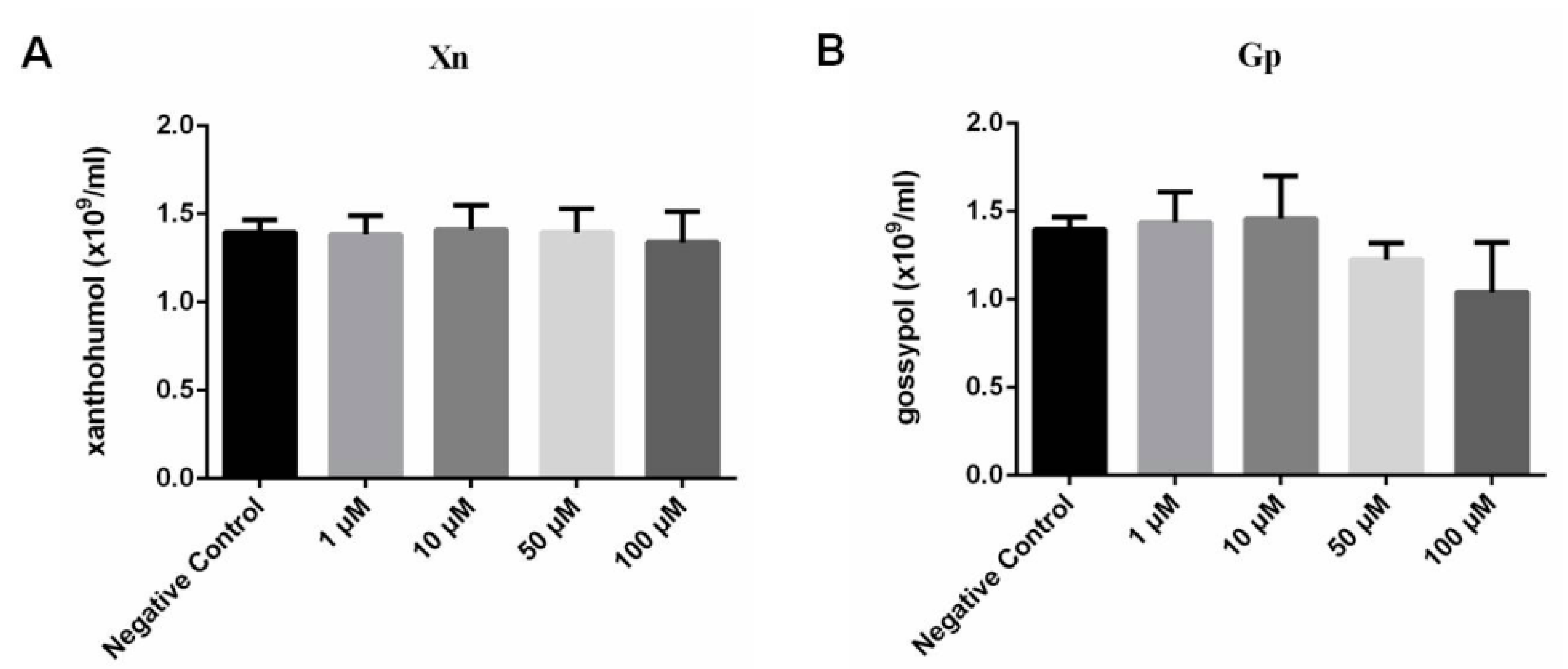
3.2. Cytotoxicity of Gp and Xn
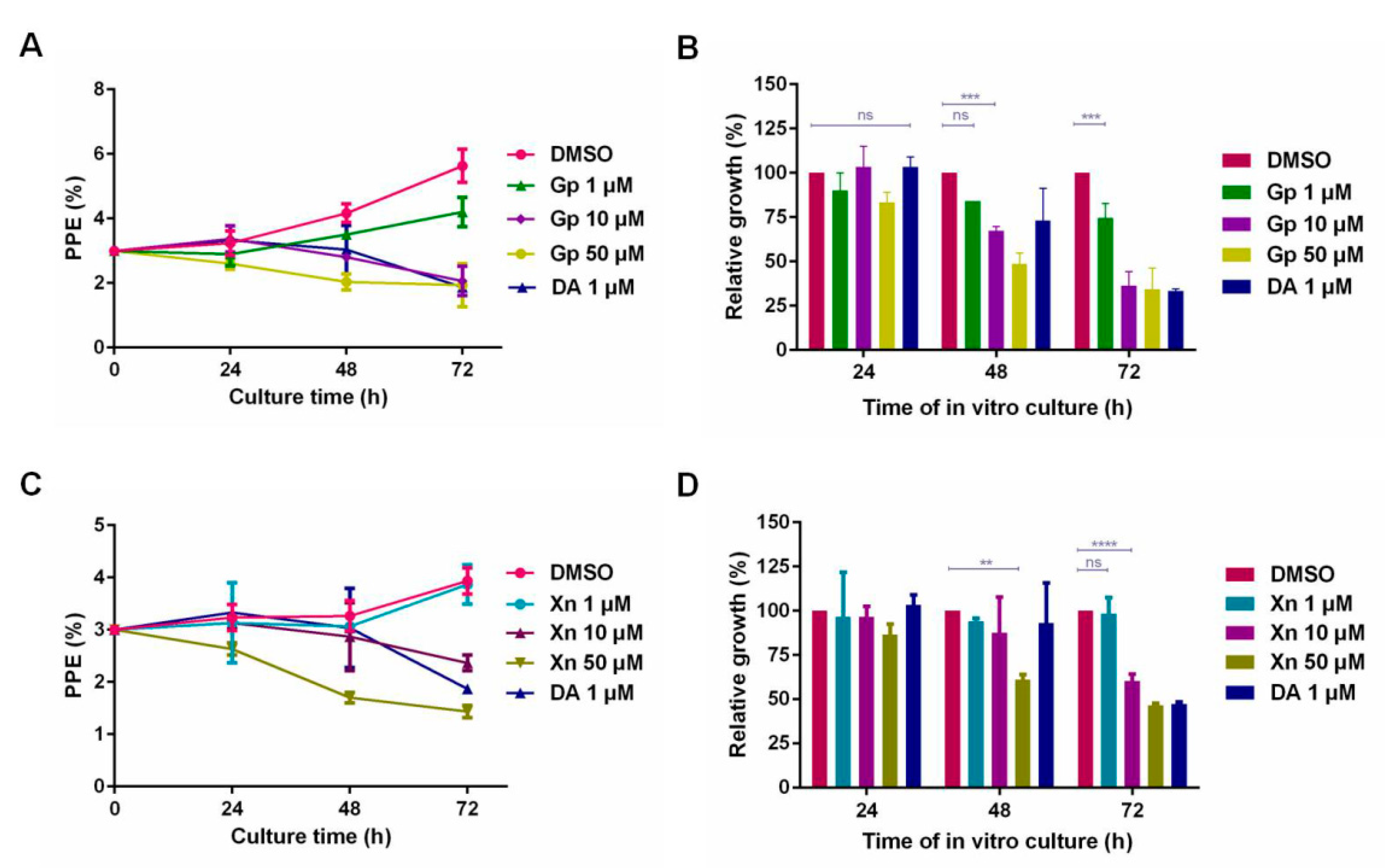
3.3. The Inhibitory Effect of Xn and Gp on the In Vitro Culture of B. microti
3.4. The Inhibitory Effect of Xn and Gp Combination on the In Vitro Culture of B. microti
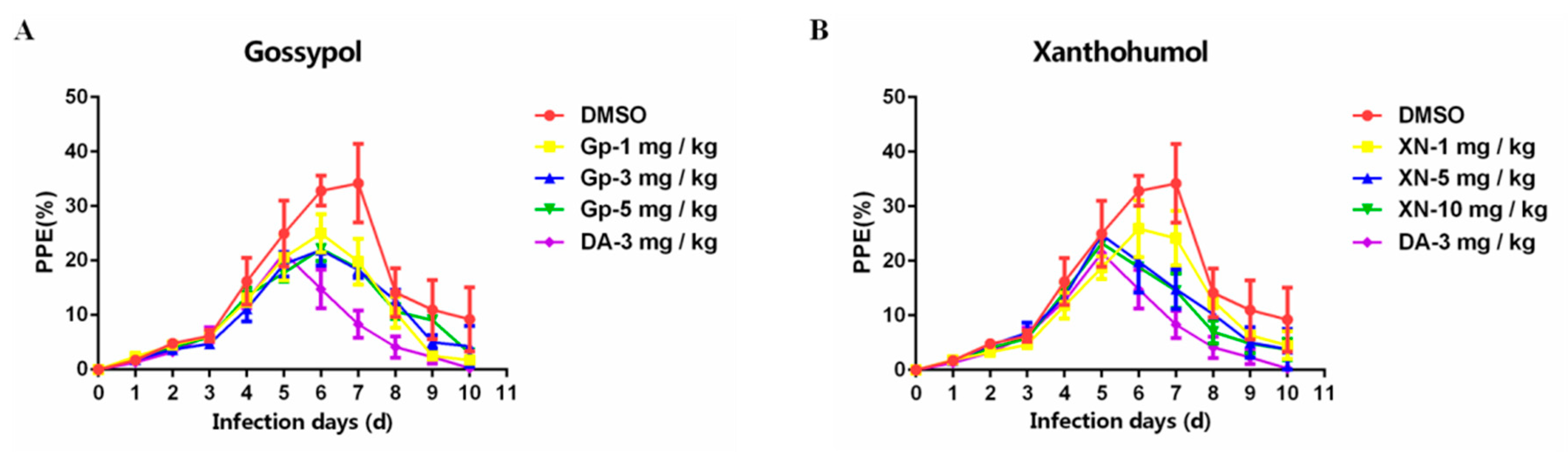
3.5. Inhibitory Effect of Xn and Gp on the In Vivo Culture of B. microti
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Rathinasamy, V.; Poole, W.A.; Bastos, R.G.; Suarez, C.E.; Cooke, B.M. Babesiosis vaccines: Lessons learned, challenges ahead, and future glimpses. Trends Parasitol. 2019, 35, 622–635. [Google Scholar] [CrossRef]
- Krause, P.J. Human Babesiosis. Int. J. Parasitol. 2019, 49, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, E.; Vannier, E.; Wormser, G.P.; Hu, L.T. Diagnosis, Treatment and prevention of lyme disease, human granulocytic anaplasmosis, and babesiosis. JAMA 2016, 315, 1767–1777. [Google Scholar] [CrossRef] [PubMed]
- Jajosky, R.P.; Jajosky, A.N.; Jajosky, P.G. The centers for disease control and prevention and state health departments should include blood–type variables in their babesiosis case reports. Transfus. Apher. Sci. 2020, 102824. [Google Scholar] [CrossRef] [PubMed]
- Ingram, D.; Crook, T. Rise in babesiosis cases, pennsylvania, USA, 2005–2018. Emerg. Infect. Dis. 2020, 26, 1703–1709. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Xia, S.; Huang, J.L.; Tambo, E.; Zhuge, H.X.; Zhou, X.N. human babesiosis, an emerging tick–borne disease in the people’s Republic Of China. Parasites Vectors 2014, 7, 509. [Google Scholar] [CrossRef]
- Westblade, L.F.; Simon, M.S.; Mathison, B.A.; Kirkman, L.A. Babesia Microti: From mice to ticks to an increasing number of highly susceptible humans. J. Clin. Microbiol. 2017, 55, 2903–2912. [Google Scholar] [CrossRef]
- Chen, Z.; Li, H.; Gao, X.; Bian, A.; Yan, H.; Kong, D.; Liu, X.-Y. Human babesiosis in china: A systematic review. Parasitol. Res. 2019, 118, 1103–1112. [Google Scholar] [CrossRef]
- Hildebrandt, A.; Gray, J.S.; Hunfeld, K.-P. Human babesiosis in europe: What clinicians need to know. Infection 2013, 41, 1057–1072. [Google Scholar] [CrossRef]
- Peniche-Lara, G.; Balmaceda, L.; Perez-Osorio, C.; Munoz-Zanzi, C. Human Babesiosis, Yucatán State, Mexico, 2015. Emerg. Infect. Dis. 2018, 24, 2061–2062. [Google Scholar] [CrossRef]
- Yamasaki, M.; Watanabe, N.; Idaka, N.; Yamamori, T.; Otsuguro, K.-I.; Uchida, N.; Iguchi, A.; Ohta, H.; Takiguchi, M. Intracellular diminazene aceturate content and adenosine incorporation in diminazene aceturate-resistant babesia gibsoni isolate in vitro. Exp. Parasitol. 2017, 183, 92–98. [Google Scholar] [CrossRef]
- Vial, H.J.; Gorenflot, A. Chemotherapy against babesiosis. Veter. Parasitol. 2006, 138, 147–160. [Google Scholar] [CrossRef]
- Oguejiofor, C.F.; Ochiogu, I.; Umeoduagu, C. Increasing doses of diminazene aceturate: Adverse reproductive effects in female Wistar rats. Asian Pac. J. Trop. Med. 2010, 3, 887–889. [Google Scholar] [CrossRef]
- Checa, R.; Montoya, A.; Ortega, N.; González-Fraga, J.L.; Bartolomé, A.; Gálvez, R.; Marino, V.; Miró, G. Efficacy, safety and tolerance of imidocarb dipropionate versus atovaquone or buparvaquone plus azithromycin used to treat sick dogs naturally infected with the babesia microti-like piroplasm. Par. Vect. 2017, 10, 145. [Google Scholar] [CrossRef] [PubMed]
- Schwint, O.N.; Ueti, M.W.; Palmer, G.H.; Kappmeyer, L.S.; Hines, M.T.; Cordes, R.T.; Knowles, D.P.; Scoles, G.A. Imidocarb Dipropionate Clears Persistent Babesia Caballi Infection with Elimination of Transmission Potential. Antimicrob. Agents Chemother. 2009, 53, 4327–4332. [Google Scholar] [CrossRef]
- Wijaya, A.; Wulansari, R.; Ano, H.; Makimura, S. Effect of clindamycin therapy on phagocytic and oxidative activity profiles of spleen mononuclear cells in babesia rodhaini-infected mice. J. Veter. Med. Sci. 2001, 63, 563–566. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Raju, M.; Salazar, J.C.; Leopold, H.; Krause, P.J. Atovaquone and azithromycin treatment for babesiosis in an infant. Pediatr. Infect. Dis. J. 2007, 26, 181–183. [Google Scholar] [CrossRef] [PubMed]
- Krause, P.J.; Lepore, T.; Sikand, V.K.; Gadbaw, J.; Burke, G.; Telford, S.R.; Brassard, P.; Pearl, D.; Azlanzadeh, J.; Christianson, D.; et al. Atovaquone and azithromycin for the treatment of babesiosis. N. Engl. J. Med. 2000, 343, 1454–1458. [Google Scholar] [CrossRef]
- Smith, R.P.; Hunfeld, K.-P.; Krause, P.J. Management strategies for human babesiosis. Expert Rev. Anti-Infect. Ther. 2020, 18, 625–636. [Google Scholar] [CrossRef]
- Yuan, R.; Hou, Y.; Sun, W.; Yu, J.; Liu, X.; Niu, Y.; Lu, J.-J.; Chen, X. Natural Products To Prevent Drug Resistance In Cancer Chemotherapy: A Review. Ann. N. Y. Acad. Sci. 2017, 1401, 19–27. [Google Scholar] [CrossRef]
- Harvey, A.L.; Edrada-Ebel, R.; Quinn, R.J. The re-emergence of natural products for drug discovery in the genomics Era. Nat. Rev. Drug Discov. 2015, 14, 111–129. [Google Scholar] [CrossRef] [PubMed]
- Marrero, F.E. Natural products for drug discovery. Toxicol. Lett. 2016, 259, S15. [Google Scholar] [CrossRef]
- Moloney, M.G. Natural products as a source for novel antibiotics. Trends Pharmacol. Sci. 2016, 37, 689–701. [Google Scholar] [CrossRef] [PubMed]
- Tayebwa, D.S.; Tuvshintulga, B.; Guswanto, A.; Nugraha, A.B.; Batiha, G.E.-S.; Gantuya, S.; Rizk, M.A.; Vudriko, P.; Sivakumar, T.; Yokoyama, N.; et al. The Effects of Nitidine Chloride and Camptothecin on the growth of babesia and theileria parasites. Ticks Tick-Borne Dis. 2018, 9, 1192–1201. [Google Scholar] [CrossRef]
- Halasi, M.; Hitchinson, B.; Shah, B.N.; Váraljai, R.; Khan, I.; Benevolenskaya, E.V.; Gaponenko, V.; Arbiser, J.L.; Gartel, A.L. Honokiol Is a foxm1 antagonist. Cell Death Dis. 2018, 9, 84. [Google Scholar] [CrossRef]
- Jiang, X.; Chen, L.; Zheng, Z.; Chen, Y.; Weng, X.; Guo, Y.; Li, K.; Yang, T.; Qu, S.; Liu, H.; et al. Synergistic effect of combined artesunate and tetramethylpyrazine in experimental cerebral malaria. ACS Infect. Dis. 2020, 6, 2400–2409. [Google Scholar] [CrossRef] [PubMed]
- Kyei-Baffour, K.; Davis, D.C.; Boskovic, Z.; Kato, N.; Dai, M. Natural product-inspired aryl isonitriles as a new class of antimalarial compounds against drug-resistant parasites. Bioorg. Med. Chem. 2020, 28, 115678. [Google Scholar] [CrossRef]
- Goo, Y.-K.; Terkawi, M.A.; Jia, H.; Aboge, G.O.; Ooka, H.; Nelson, B.; Kim, S.; Sunaga, F.; Namikawa, K.; Igarashi, I.; et al. Artesunate, A potential drug for treatment of babesia infection. Parasitol. Int. 2010, 59, 481–486. [Google Scholar] [CrossRef]
- Li, M.; Ao, Y.; Guo, J.; Nie, Z.; Liu, Q.; Yu, L.; Luo, X.; Zhan, X.; Zhao, Y.; Wang, S.; et al. Surface Antigen 1 Is a crucial secreted protein that mediates babesia microti invasion into host cells. Front. Microbiol. 2020, 10, 3046. [Google Scholar] [CrossRef]
- Chou, T.-C. Theoretical Basis, Experimental Design, and Computerized Simulation of Synergism and Antagonism in Drug Combination Studies. Pharmacol. Rev. 2006, 58, 621–681. [Google Scholar] [CrossRef]
- Zhang, B.; Duan, D.; Ge, C.; Yao, J.; Liu, Y.; Li, X.; Fang, J. Synthesis of xanthohumol analogues and discovery of potent thioredoxin reductase inhibitor as potential anticancer agent. J. Med. Chem. 2015, 58, 1795–1805. [Google Scholar] [CrossRef] [PubMed]
- Logan, I.E.; Miranda, C.L.; Lowry, M.B.; Maier, C.S.; Stevens, J.F.; Gombart, A.F. Antiproliferative and cytotoxic activity of xanthohumol and its non-estrogenic derivatives in colon and hepatocellular carcinoma cell lines. Int. J. Mol. Sci. 2019, 20, 1203. [Google Scholar] [CrossRef]
- Zhang, L.; Jiang, H.; Cao, X.; Zhao, H.; Wang, F.; Cui, Y.; Jiang, B. Chiral gossypol derivatives: Evaluation of their anticancer activity and molecular modeling. Eur. J. Med. Chem. 2009, 44, 3961–3972. [Google Scholar] [CrossRef] [PubMed]
- Hussong, R.; Frank, N.; Knauft, J.; Ittrich, C.; Owen, R.; Becker, H.; Gerhäuser, C. A safety study of oral xanthohumol administration and its influence on fertility in sprague dawley rats. Mol. Nutr. Food Res. 2005, 49, 861–867. [Google Scholar] [CrossRef]
- Simon, M.S.; Westblade, L.F.; Dziedziech, A.; Visone, J.E.; Furman, R.R.; Jenkins, S.G.; Schuetz, A.N.; Kirkman, L.A. Clinical and molecular evidence of atovaquone and azithromycin resistance in relapsed babesia microti infection associated with rituximab and chronic lymphocytic leukemia. Clin. Infect. Dis. 2017, 65, 1222–1225. [Google Scholar] [CrossRef]
- Nonaka, M.; Murata, Y.; Takano, R.; Han, Y.; Bin Kabir, H.; Kato, K. Screening of a library of traditional chinese medicines to identify anti-malarial compounds and extracts. Malar. J. 2018, 17, 244. [Google Scholar] [CrossRef]
- Haj, E.; Losev, Y.; Krishnakumar, V.G.; Pichinuk, E.; Engel, H.; Raveh, A.; Gazit, E.; Segal, D. integrating in vitro and in silico approaches to evaluate the “dual functionality” of palmatine chloride in inhibiting and disassembling tau-derived peptide fibrils. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2018, 1862, 1565–1575. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-A.; Kang, O.-H.; Kwon, D.-Y. Cryptotanshinone induces cell cycle arrest and apoptosis of nsclc cells through the pi3k/akt/gsk-3β Pathway. Int. J. Mol. Sci. 2018, 19, 2739. [Google Scholar] [CrossRef]
- Bloch, E.M.; Lee, T.H.; Krause, P.J.; Telford, S.R.; Montalvo, L.; Chafets, D.; Usmani-Brown, S.; Lepore, T.J.; Busch, M.P. development of a real-time polymerase chain reaction assay for sensitive detection and quantitation of babesia microti infection. Transfusion 2013, 53, 2299–2306. [Google Scholar]
- Guswanto, A.; Sivakumar, T.; Rizk, M.A.; Elsayed, S.A.E.; Youssef, M.A.; Elsaid, E.E.S.; Yokoyama, N.; Igarashi, I. Evaluation of a fluorescence-based method for antibabesial drug screening. Antimicrob. Agents Chemother. 2014, 58, 4713–4717. [Google Scholar] [CrossRef]
- Rizk, M.A.; El-Sayed, S.A.E.-S.; Aboulaila, M.; Eltaysh, R.; Yokoyama, N.; Igarashi, I. Performance and consistency of a fluorescence-based high-throughput screening assay for use in babesia drug screening in mice. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Teal, A.E.; Habura, A.; Ennis, J.; Keithly, J.S.; Madison-Antenucci, S. A New Real-Time PCR assay for improved detection of the parasite babesia microti. J. Clin. Microbiol. 2011, 50, 903–908. [Google Scholar] [CrossRef]
- Augoff, K.; Hryniewicz-Jankowska, A.; Tabola, R. Lactate dehydrogenase 5: An old friend and a new hope in the war on cancer. Cancer Lett. 2015, 358, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Dando, C.; Schroeder, E.R.; Hunsaker, L.A.; Deck, L.M.; Royer, R.E.; Zhou, X.; Parmley, S.F.; Jagt, D.L.V. The kinetic properties and sensitivities to inhibitors of lactate dehydrogenases (ldh1 and ldh2) from toxoplasma gondii: Comparisons with pldh from plasmodium falciparum. Mol. Biochem. Parasitol. 2001, 118, 23–32. [Google Scholar] [CrossRef]
- Bork, S.; Okamura, M.; Boonchit, S.; Hirata, H.; Yokoyama, N.; Igarashi, I. Identification of babesia bovis l-lactate dehydrogenase as a potential chemotherapeutical target against bovine babesiosis. Mol. Biochem. Parasitol. 2004, 136, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Razakantoanina, V.; Phung, N.K.P.; Jaureguiberry, G. Antimalarial activity of new gossypol derivatives. Parasitol. Res. 2000, 86, 665–668. [Google Scholar] [CrossRef]
- Balakrishnan, K.; Wierda, W.G.; Keating, M.J.; Gandhi, V. Gossypol, A BH3 mimetic, induces apoptosis in chronic lymphocytic leukemia cells. Blood 2008, 112, 1971–1980. [Google Scholar] [CrossRef]
- Shaaban, W.F.; Taha, T.; El-Nouty, F.D.; El-Mahdy, A.R.; Salem, M.H. Reproductive toxicologic effects of gossypol on male rabbits: Biochemical, enzymatic, and electrolytic properties of seminal plasma. Fertil. Steril. 2008, 89, 1585–1593. [Google Scholar] [CrossRef]
- El-Sharaky, A.; Newairy, A.; Elguindy, N.; Elwafa, A. Spermatotoxicity, biochemical changes and histological alteration induced by gossypol in testicular and hepatic tissues of male rats. Food Chem. Toxicol. 2010, 48, 3354–3361. [Google Scholar] [CrossRef]
- Sharma, S.; Kumar, M.; Goyal, R.; Manivannan, B.; Lohiya, N. Reversible Antispermatogenic effect of gossypol in langur monkeys (presbytis entellus entellus). Adv. Contracept. 1999, 15, 15–27. [Google Scholar] [CrossRef]
- Romualdo, G.S.; Klinefelter, G.R.; De, K. Postweaning exposure to gossypol results in epididymis-specific effects throughout puberty and adulthood in rats. J. Androl. 2002, 23, 220–228. [Google Scholar]
- Basini, G.; Bussolati, S.; Baioni, L.; Grasselli, F. Gossypol, A polyphenolic aldehyde from cotton plant, interferes with swine granulosa cell function. Domest. Anim. Endocrinol. 2009, 37, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Monzote, L.; Lackova, A.; Staniek, K.; Steinbauer, S.; Pichler, G.; Jäger, U.-P.D.W.; Gille, L. The antileishmanial activity of xanthohumol is mediated by mitochondrial inhibition. Parasitology 2016, 144, 747–759. [Google Scholar] [CrossRef] [PubMed]
- Gerhauser, C. Broad spectrum anti-infective potential of xanthohumol from hop (humulus lupulus l.) in comparison with activities of other hop constituents and xanthohumol metabolites. Mol. Nutr. Food. Res. 2005, 49, 827–831. [Google Scholar] [CrossRef] [PubMed]
- Gerhäuser, C.; Alt, A.; Heiss, E.; Gamal-Eldeen, A.; Klimo, K.; Knauft, J.; Neumann, I.; Scherf, H.-R.; Frank, N.; Bartsch, H.; et al. Cancer chemopreventive activity of xanthohumol, a natural product derived from hop. Mol. Cancer Ther. 2002, 1, 959–969. [Google Scholar] [PubMed]
- Stevens, J.F.; Page, J.E. Xanthohumol and related prenylflavonoids from hops and beer: To your good health! Phytochemistry 2004, 65, 1317–1330. [Google Scholar] [CrossRef]
- Dorn, C.; Bataille, F.; Gaebele, E.; Heilmann, J.; Hellerbrand, C. Xanthohumol Feeding does not impair organ function and homoeostasis in Mice. Food Chem. Toxicol. 2010, 48, 1890–1897. [Google Scholar] [CrossRef]
- Van Breemen, R.B.; Yuan, Y.; Banuvar, S.; Shulman, L.P.; Qiu, X.; Alvarenga, R.F.R.; Chen, S.-N.; Dietz, B.M.; Bolton, J.L.; Pauli, G.F.; et al. Pharmacokinetics of Prenylated Hop Phenols in Women Following Oral Administration of a standardized extract of hops. Mol. Nutr. Food Res. 2014, 58, 1962–1969. [Google Scholar] [CrossRef]
- Tronina, T.; Strugała, P.; Popłoński, J.; Włoch, A.; Sordon, S.; Bartmańska, A.; Huszcza, E. The influence of glycosylation of natural and synthetic prenylated flavonoids on binding to human serum albumin and inhibition of cyclooxygenases cox-1 and cox-2. Molecules 2017, 22, 1230. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nature Products | Organism/Cell Type | IC50 | Selectivity Index a |
|---|---|---|---|
| Gossypol | erythrocyte of KM | >100 μm | >11.8 |
| chronic lymphocytic leukemia cells | 30 μm | 3.50 | |
| epithelial cervix cancer cells (HeLa) | 31.3 μm | 3.70 | |
| brain malignant glioma cells (U87) | 59.6 μm | 7.00 | |
| gastric cancer cells (M85) | 39.7 μm | 4.70 | |
| Xanthohumol | erythrocyte of KM | >100 μm | >4.7 |
| human colon adenocarcinoma cell lines (HT29) | 50.2 μm | 2.30 | |
| human colon adenocarcinoma cell lines (HCT116) | 40.8 μm | 1.90 | |
| human hepatocellular carcinoma cell lines (Huh7) | 37.2 μm | 1.70 | |
| hypotriploid alveolar basal epithelial cells (A549) | 30.5 μm | 1.40 | |
| epithelial cervix cancer cells (HeLa) | 40.4 μm | 1.90 | |
| human hepatocellular carcinoma cell lines (HepG2) | 35 μm | 1.60 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, J.; Luo, X.; Wang, S.; He, L.; Zhao, J. Xanthohumol and Gossypol Are Promising Inhibitors against Babesia microti by In Vitro Culture via High-Throughput Screening of 133 Natural Products. Vaccines 2020, 8, 613. https://doi.org/10.3390/vaccines8040613
Guo J, Luo X, Wang S, He L, Zhao J. Xanthohumol and Gossypol Are Promising Inhibitors against Babesia microti by In Vitro Culture via High-Throughput Screening of 133 Natural Products. Vaccines. 2020; 8(4):613. https://doi.org/10.3390/vaccines8040613
Chicago/Turabian StyleGuo, Jiaying, Xiaoying Luo, Sen Wang, Lan He, and Junlong Zhao. 2020. "Xanthohumol and Gossypol Are Promising Inhibitors against Babesia microti by In Vitro Culture via High-Throughput Screening of 133 Natural Products" Vaccines 8, no. 4: 613. https://doi.org/10.3390/vaccines8040613
APA StyleGuo, J., Luo, X., Wang, S., He, L., & Zhao, J. (2020). Xanthohumol and Gossypol Are Promising Inhibitors against Babesia microti by In Vitro Culture via High-Throughput Screening of 133 Natural Products. Vaccines, 8(4), 613. https://doi.org/10.3390/vaccines8040613