Recombinant Rabies Virus Overexpressing OX40-Ligand Enhances Humoral Immune Responses by Increasing T Follicular Helper Cells and Germinal Center B Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells, Viruses, Antibodies, and Animals
2.2. Construction and Rescue of rRABV Expressing Murine OX40L
2.3. Virus Titration
2.4. Cell Viability Assay
2.5. OX40L Concentration Determination through ELISA
2.6. Mouse Immunization and Challenge Test
2.7. Pathogenicity Studies
2.8. Detection of Virus-Neutralizing Antibody (VNA)
2.9. RABV-Specific Antibody Isotype Test
2.10. Flow Cytometric Assay
2.11. Statistical Analysis
3. Results
3.1. Characterization of rRABV Expressing OX40L
3.2. Pathogenicity of rRABV Expressing OX40L
3.3. Generation of Tfh Cells in Mice Immunized with LBNSE-OX40L
3.4. Generation of GC B Cells in Mice Immunized with LBNSE-OX40L
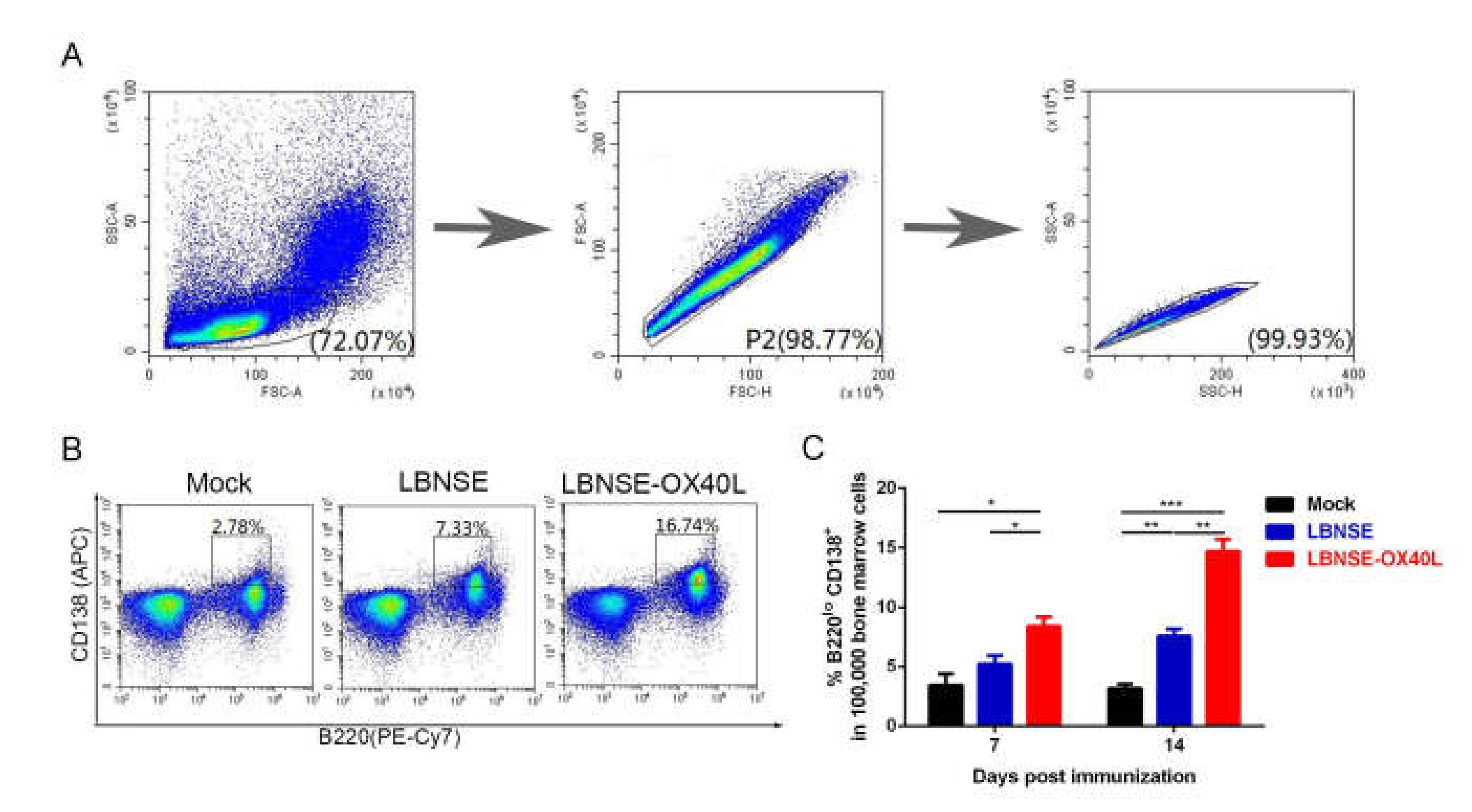
3.5. Generation of PCs in Mice Immunized with LBNSE-OX40L
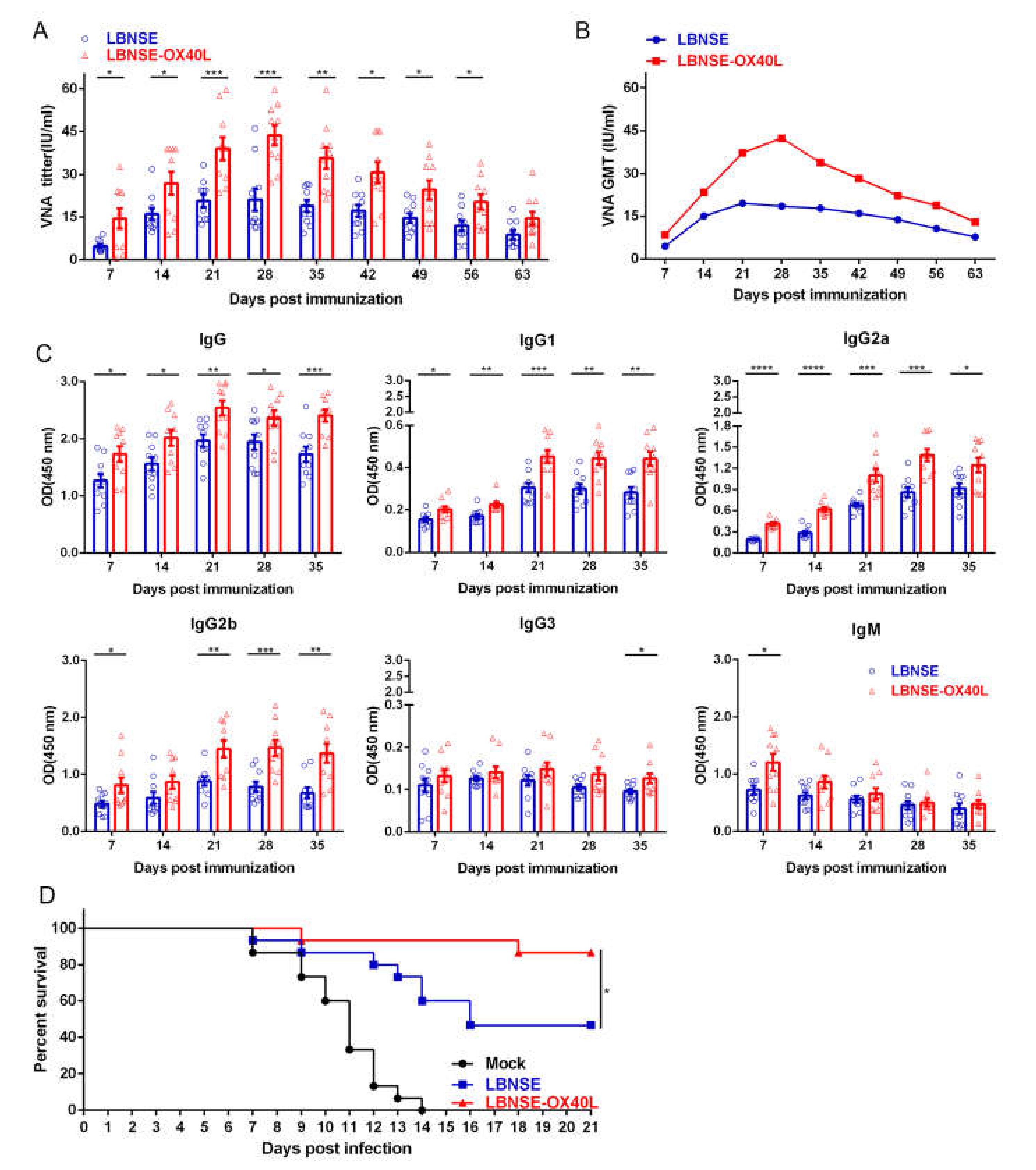
3.6. Antibody Responses and Protection in Mice Immunized with LBNSE-OX40L
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Barkhouse, D.A.; Faber, M.; Hooper, D.C. Pre- and post-exposure safety and efficacy of attenuated rabies virus vaccines are enhanced by their expression of IFNgamma. Virology 2015, 474, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Mazeri, S.; Gibson, A.D.; Meunier, N.; Bronsvoort, B.M.D.; Gamble, L. Barriers of attendance to dog rabies static point vaccination clinics in Blantyre, Malawi. PLoS Negl. Trop. Dis. 2018, 12, e0006159. [Google Scholar] [CrossRef]
- Tarantola, A.; Blanchi, S.; Cappelle, J.; Ly, S.; Chan, M.; In, S.; Peng, Y.; Hing, C.; Taing, C.N.; Ly, S.; et al. Rabies Postexposure Prophylaxis Noncompletion After Dog Bites: Estimating the Unseen to Meet the Needs of the Underserved. Am. J. Epidemiol. 2018, 187, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Ertl, H.C.J. New Rabies Vaccines for Use in Humans. Vaccines 2019, 7, 54. [Google Scholar] [CrossRef] [PubMed]
- Franka, R.; Smith, T.G.; Dyer, J.L.; Wu, X.; Niezgoda, M.; Rupprecht, C.E. Current and future tools for global canine rabies elimination. Antivir. Res. 2013, 100, 220–225. [Google Scholar] [CrossRef]
- Schutsky, K.; Curtis, D.; Bongiorno, E.K.; Barkhouse, D.A.; Kean, R.B.; Dietzschold, B.; Hooper, D.C.; Faber, M. Intramuscular Inoculation of Mice with the Live-Attenuated Recombinant Rabies Virus TriGAS Results in a Transient Infection of the Draining Lymph Nodes and a Robust, Long-Lasting Protective Immune Response against Rabies. J. Virol. 2013, 87, 1834–1841. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, K.; Shoji, Y.; Inoue, S. Characterization of P gene-deficient rabies virus: Propagation, pathogenicity and antigenicity. Virus Res. 2005, 111, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Ito, N.; Sugiyama, M.; Yamada, K.; Shimizu, K.; Takayama-Ito, M.; Hosokawa, J.; Minamoto, N. Characterization of M gene-deficient rabies virus with advantages of effective immunization and safety as a vaccine strain. Microbiol. Immunol. 2005, 49, 971–979. [Google Scholar] [CrossRef]
- Zhao, L.; Toriumi, H.; Wang, H.; Kuang, Y.; Guo, X.; Morimoto, K.; Fu, Z.F. Expression of MIP-1alpha (CCL3) by a recombinant rabies virus enhances its immunogenicity by inducing innate immunity and recruiting dendritic cells and B cells. J. Virol. 2010, 84, 9642–9648. [Google Scholar] [CrossRef]
- Zhou, M.; Zhang, G.; Ren, G.; Gnanadurai, C.W.; Li, Z.; Chai, Q.; Yang, Y.; Leyson, C.M.; Wu, W.; Cui, M.; et al. Recombinant rabies viruses expressing GM-CSF or flagellin are effective vaccines for both intramuscular and oral immunizations. PLoS ONE 2013, 8, e63384. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, M.; Luo, Z.; Zhang, Y.; Cui, M.; Chen, H.; Fu, Z.F.; Zhao, L. Overexpression of Interleukin-7 Extends the Humoral Immune Response Induced by Rabies Vaccination. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed]
- Linton, P.J.; Bautista, B.; Biederman, E.; Bradley, E.S.; Harbertson, J.; Kondrack, R.M.; Padrick, R.C.; Bradley, L.M. Costimulation via OX40L Expressed by B Cells Is Sufficient to Determine the Extent of Primary CD4 Cell Expansion and Th2 Cytokine Secretion In Vivo. J. Exp. Med. 2003, 197, 875–883. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, S.J.; Perona-Wright, G.; Worsley, A.G.F.; Ishii, N.; MacDonald, A.S. Dendritic Cell Expression of OX40 Ligand Acts as a Costimulatory, Not Polarizing, Signal for Optimal Th2 Priming and Memory Induction In Vivo. J. Immunol. 2007, 179, 3515–3523. [Google Scholar] [CrossRef]
- Webb, G.J.; Hirschfield, G.M.; Lane, P.J.L. OX40, OX40L and Autoimmunity: A Comprehensive Review. Clin. Rev. Allergy Immunol. 2016, 50, 312–332. [Google Scholar] [CrossRef]
- Jacquemin, C.; Schmitt, N.; Contin-Bordes, C.; Liu, Y.; Blanco, P. OX40 Ligand Contributes to Human Lupus Pathogenesis by Promoting T Follicular Helper Response. Immunity 2015, 42, 1159–1170. [Google Scholar] [CrossRef] [PubMed]
- Eckhard Stiiber, W.S. The T cell-B cell interaction via OX40-OX40L is necessary for the T cell-dependent humoral immune response. J. Exp. Med. 1996, 183, 979–989. [Google Scholar]
- Tahiliani, V.; Hutchinson, T.E.; Abboud, G.; Croft, M.; Salek-Ardakani, S. OX40 Cooperates with ICOS To Amplify Follicular Th Cell Development and Germinal Center Reactions during Infection. J. Immunol. 2016, 198, 218–228. [Google Scholar] [CrossRef]
- Du, X.; Kang, Y.; Wang, X.; Wang, J.; Zhao, G.; Wang, B. Adjuvant effect of plasmid vector-expressed OX40L on candidate DNA vaccine against type B heptatitis. Acta Microbiol. Sin. 2009, 49, 357–362. [Google Scholar]
- Wen, Y.J.; Wang, H.L.; Wu, H.; Yang, F.H.; Tripp, R.A.; Hogan, R.J.; Fu, Z.F. Rabies Virus Expressing Dendritic Cell-Activating Molecules Enhances the Innate and Adaptive Immune Response to Vaccination. J. Virol. 2011, 85, 1634–1644. [Google Scholar] [CrossRef]
- Cliquet, F.; Aubert, M.; Sagne, L. Development of a fluorescent antibody virus neutralisation test (FAVN test) for the quantitation of rabies-neutralising antibody. J. Immunol. Methods 1998, 212, 79–87. [Google Scholar] [CrossRef]
- Bedard, E.L.; Inculet, R.I.; Malthaner, R.A.; Brecevic, E.; Vincent, M.; Dar, R. The role of surgery and postoperative chemoradiation therapy in patients with lymph node positive esophageal carcinoma. Cancer 2001, 91, 2423–2430. [Google Scholar] [CrossRef]
- Lee, S.K.; Rigby, R.J.; Zotos, D.; Tsai, L.M.; Kawamoto, S.; Marshall, J.L.; Ramiscal, R.R.; Chan, T.D.; Gatto, D.; Brink, R.; et al. B cell priming for extrafollicular antibody responses requires Bcl-6 expression by T cells. J. Exp. Med. 2011, 208, 1377–1388. [Google Scholar] [CrossRef] [PubMed]
- Linterman, M.A.; Hill, D.L. Can follicular helper T cells be targeted to improve vaccine efficacy? F1000Research 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Murillo, P.; Pramanik, L.; Sundling, C.; Hultenby, K.; Wretenberg, P.; Spangberg, M.; Karlsson Hedestam, G.B. CD138 and CD31 Double-Positive Cells Comprise the Functional Antibody-Secreting Plasma Cell Compartment in Primate Bone Marrow. Front. Immunol. 2016, 7, 242. [Google Scholar] [CrossRef]
- Ramakrishna, C.; Stohlman, S.A.; Atkinson, R.D.; Shlomchik, M.J.; Bergmann, C.C. Mechanisms of central nervous system viral persistence: The critical role of antibody and B cells. J. Immunol. 2002, 168, 1204–1211. [Google Scholar] [CrossRef] [PubMed]
- Dorfmeier, C.L.; Tzvetkov, E.P.; Gatt, A.; McGettigan, J.P. Investigating the role for IL-21 in rabies virus vaccine-induced immunity. PLoS Negl. Trop. Dis. 2013, 7, e2129. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, J.; Li, M.; Cui, M.; Fu, Z.F.; Zhao, L.; Zhou, M. A Recombinant Rabies Virus Expressing Fms-like Tyrosine Kinase 3 Ligand (Flt3L) Induces Enhanced Immunogenicity in Mice. Virol. Sin. 2019, 34, 662–672. [Google Scholar] [CrossRef]
- Liu, J.; Ngai, N.; Stone, G.W.; Yue, F.Y.; Ostrowski, M.A. The adjuvancy of OX40 ligand (CD252) on an HIV-1 canarypox vaccine. Vaccine 2009, 27, 5077–5084. [Google Scholar] [CrossRef]
- Gopisetty, A.; Bhattacharya, P.; Haddad, C.; Bruno, J.C.; Vasu, C.; Miele, L.; Prabhakar, B.S. OX40L/Jagged1 Cosignaling by GM-CSF-Induced Bone Marrow-Derived Dendritic Cells Is Required for the Expansion of Functional Regulatory T Cells. J. Immunol. 2013, 190, 5516–5525. [Google Scholar] [CrossRef]
- Grosenbach, D.W.; Schlom, J.; Gritz, L.; Gomez Yafal, A.; Hodge, J.W. A recombinant vector expressing transgenes for four T-cell costimulatory molecules (OX40L, B7-1, ICAM-1, LFA-3) induces sustained CD4+ and CD8+ T-cell activation, protection from apoptosis, and enhanced cytokine production. Cell. Immunol. 2003, 222, 45–57. [Google Scholar] [CrossRef]
- Brocker, T.; Gulbranson-Judge, A.; Flynn, S.; Riedinger, M.; Raykundalia, C.; Lane, P. CD4 T cell traffic control: In vivo evidence that ligation of OX40 on CD4 T cells by OX40-ligand expressed on dendritic cells leads to the accumulation of CD4 T cells in B follicles. Eur. J. Immunol. 1999, 29, 1610–1616. [Google Scholar] [CrossRef]
- Soroosh, P.; Ine, S.; Sugamura, K.; Ishii, N. OX40-OX40 Ligand Interaction through T Cell-T Cell Contact Contributes to CD4 T Cell Longevity. J. Immunol. 2006, 176, 5975–5987. [Google Scholar] [CrossRef] [PubMed]
- Christofolini, D.M.; Amaro, A.; Mafra, F.; Sonnewend, A.; Bianco, B.; Barbosa, C.P. CYP2C19 polymorphism increases the risk of endometriosis. J. Assist. Reprod. Genet. 2015, 32, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Crotty, S. T follicular helper cell differentiation, function, and roles in disease. Immunity 2014, 41, 529–542. [Google Scholar] [CrossRef]
- Baumjohann, D.; Preite, S.; Reboldi, A.; Ronchi, F.; Ansel, K.M.; Lanzavecchia, A.; Sallusto, F. Persistent antigen and germinal center B cells sustain T follicular helper cell responses and phenotype. Immunity 2013, 38, 596–605. [Google Scholar] [CrossRef]
- Krautler, N.J.; Suan, D.; Butt, D.; Bourne, K.; Hermes, J.R.; Chan, T.D.; Sundling, C.; Kaplan, W.; Schofield, P.; Jackson, J.; et al. Differentiation of germinal center B cells into plasma cells is initiated by high-affinity antigen and completed by Tfh cells. J. Exp. Med. 2017, 214, 1259–1267. [Google Scholar] [CrossRef]
- Vinuesa, C.G.; Linterman, M.A.; Goodnow, C.C.; Randall, K.L. T cells and follicular dendritic cells in germinal center B-cell formation and selection. Immunol. Rev. 2010, 237, 72–89. [Google Scholar] [CrossRef]
- Qiu, L.; Zhou, Y.; Yu, Q.; Zheng, S.; Wang, Z.; Huang, Q. Elevated levels of follicular T helper cells and their association with therapeutic effects in patients with chronic lymphocytic leukaemia. Immunol. Lett. 2018, 197, 15–28. [Google Scholar] [CrossRef]
- Nutt, S.L.; Tarlinton, D.M. Germinal center B and follicular helper T cells: Siblings, cousins or just good friends? Nat. Immunol. 2011, 12, 472–477. [Google Scholar] [CrossRef]
- O’Connor, B.P.; Gleeson, M.W.; Noelle, R.J.; Erickson, L.D. The rise and fall of long-lived humoral immunity: Terminal differentiation of plasma cells in health and disease. Immunol. Rev. 2003, 194, 61–76. [Google Scholar] [CrossRef]
- Salek-Ardakani, S.; Moutaftsi, M.; Sette, A.; Croft, M. Targeting OX40 promotes lung-resident memory CD8 T cell populations that protect against respiratory poxvirus infection. J. Virol. 2011, 85, 9051–9059. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Jin, H.; Hu, Y.; Kang, Y.; Wang, J.; Du, X.; Yang, Y.; She, R.; Wang, B. Enhanced protective efficacy and reduced viral load of foot-and-mouth disease DNA vaccine with co-stimulatory molecules as the molecular adjuvants. Antivir. Res. 2007, 76, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Goulding, J.; Tahiliani, V.; Salek-Ardakani, S. OX40:OX40L axis: Emerging targets for improving poxvirus-based CD8(+) T-cell vaccines against respiratory viruses. Immunol. Rev. 2011, 244, 149–168. [Google Scholar] [CrossRef] [PubMed]
- Rasalingam, P.; Rossiter, J.P.; Mebatsion, T.; Jackson, A.C. Comparative pathogenesis of the SAD-L16 strain of rabies virus and a mutant modifying the dynein light chain binding site of the rabies virus phosphoprotein in young mice. Virus Res. 2005, 111, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Faber, M.; Faber, M.L.; Li, J.; Preuss, M.A.; Schnell, M.J.; Dietzschold, B. Dominance of a nonpathogenic glycoprotein gene over a pathogenic glycoprotein gene in rabies virus. J. Virol. 2007, 81, 7041–7047. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Zhao, L.; Sui, B.; Luo, Z.; Zhang, Y.; Wang, Y. Recombinant Rabies Virus Overexpressing OX40-Ligand Enhances Humoral Immune Responses by Increasing T Follicular Helper Cells and Germinal Center B Cells. Vaccines 2020, 8, 144. https://doi.org/10.3390/vaccines8010144
Li Y, Zhao L, Sui B, Luo Z, Zhang Y, Wang Y. Recombinant Rabies Virus Overexpressing OX40-Ligand Enhances Humoral Immune Responses by Increasing T Follicular Helper Cells and Germinal Center B Cells. Vaccines. 2020; 8(1):144. https://doi.org/10.3390/vaccines8010144
Chicago/Turabian StyleLi, Yingying, Ling Zhao, Baokui Sui, Zhaochen Luo, Yachun Zhang, and Yong Wang. 2020. "Recombinant Rabies Virus Overexpressing OX40-Ligand Enhances Humoral Immune Responses by Increasing T Follicular Helper Cells and Germinal Center B Cells" Vaccines 8, no. 1: 144. https://doi.org/10.3390/vaccines8010144
APA StyleLi, Y., Zhao, L., Sui, B., Luo, Z., Zhang, Y., & Wang, Y. (2020). Recombinant Rabies Virus Overexpressing OX40-Ligand Enhances Humoral Immune Responses by Increasing T Follicular Helper Cells and Germinal Center B Cells. Vaccines, 8(1), 144. https://doi.org/10.3390/vaccines8010144