Catestatin Regulates Epithelial Cell Dynamics to Improve Intestinal Inflammation

 , ,
, , 
Abstract
1. Introduction
2. Material and Methods
2.1. Human Subjects
2.2. Mice
2.3. Peptides
2.4. Acute DSS-Induced Colitis
2.5. Assessment of DSS-Induced Colitis
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Caco-2 Epithelial Cell Culture and Treatment with CST
2.8. Quantitative Real-Time Reverse-Transcription Polymerase Chain Reaction
2.9. Western Blotting
2.10. Statistical Analysis
3. Results
3.1. CST Peptide Protein Levels Correlate Positively with mRNA Gene Expression of TJ Proteins and STAT3, and Negatively with IL-8 and IL-18 in Patients with Active UC
3.2. CST Attenuates the Onset and Severity of Acute DSS-Induced Colitis
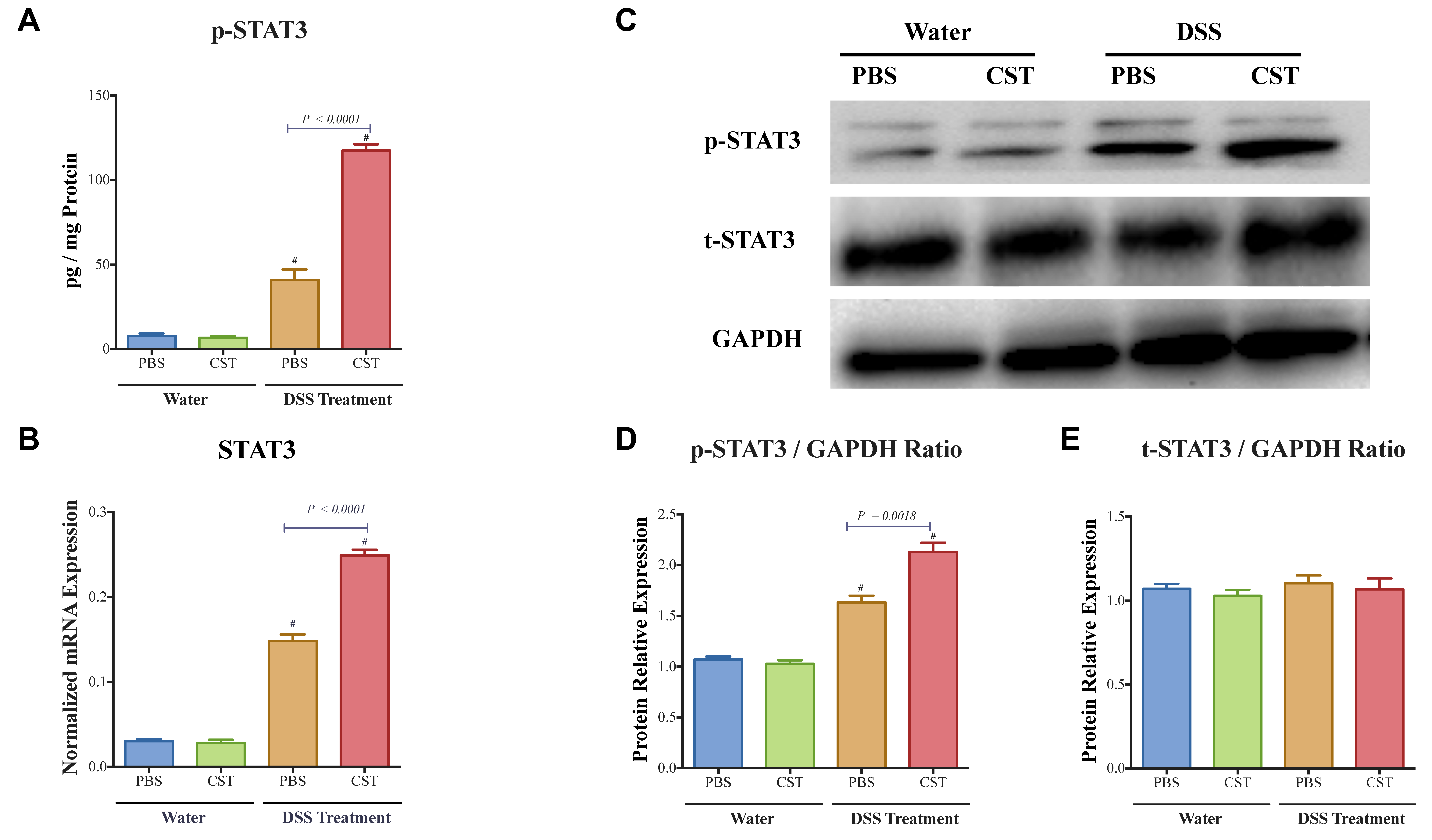
3.3. CST Increases the Activation of STAT3, Maintains Colonic Gene Expression of TJ Proteins and Decreases Colonic IL-18 Release in Acute DSS-Induced Colitis
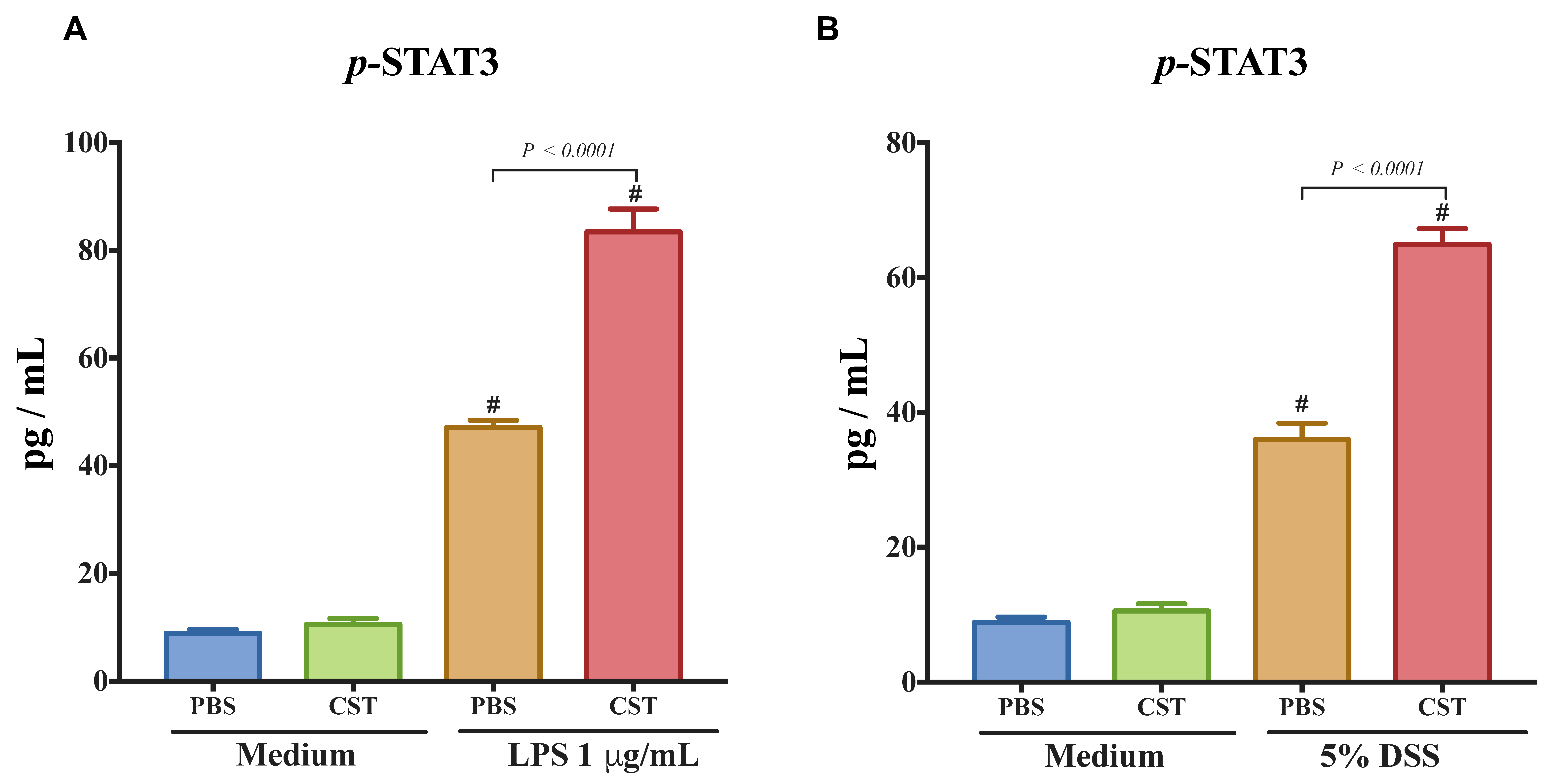
3.4. CST Enhances the Phosphorylated STAT3 (p-STAT3) in LPS- and DSS-Stimulated Caco-2 Epithelial Cells, But Not in Non-Inflammatory Conditions
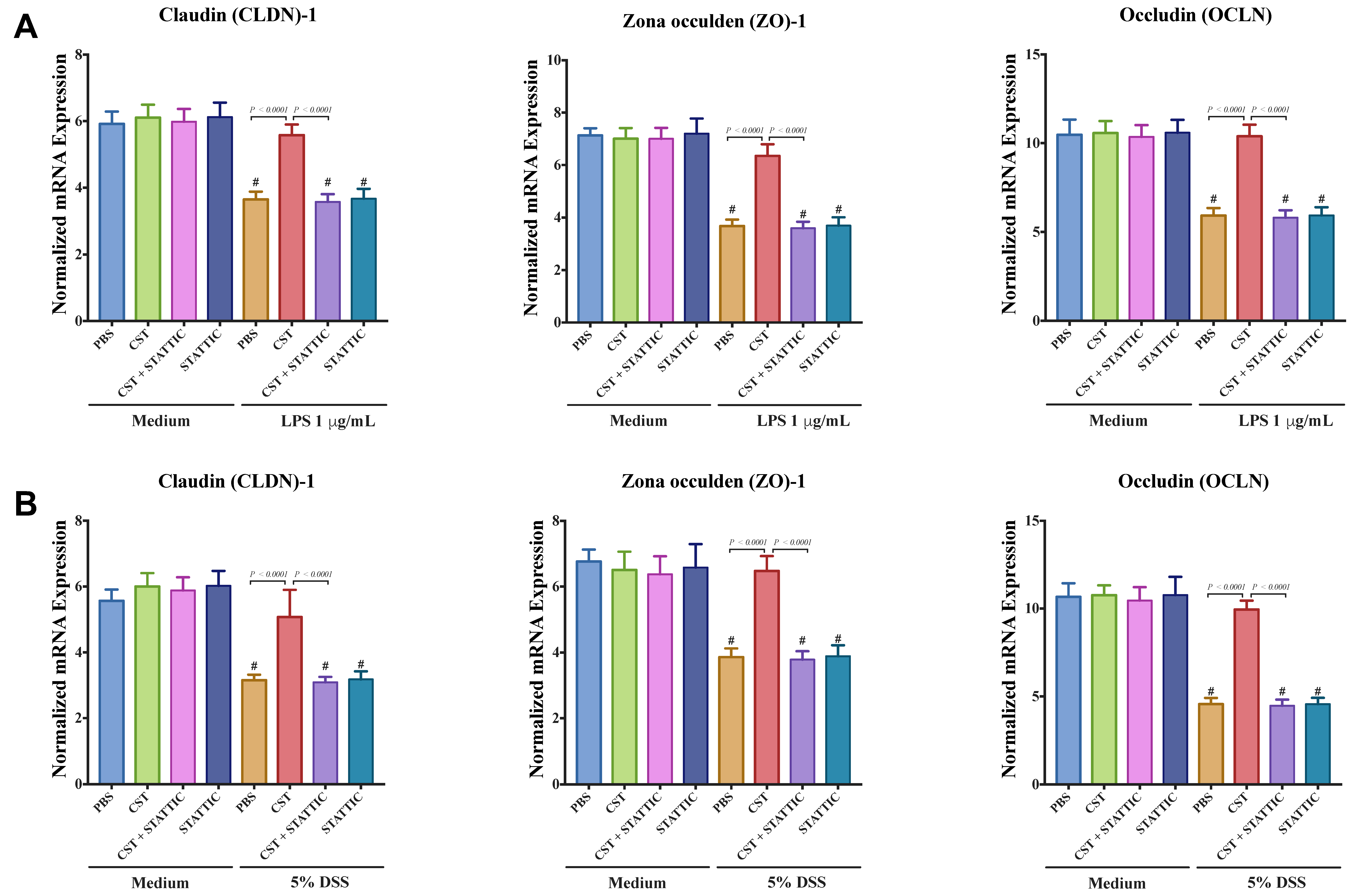
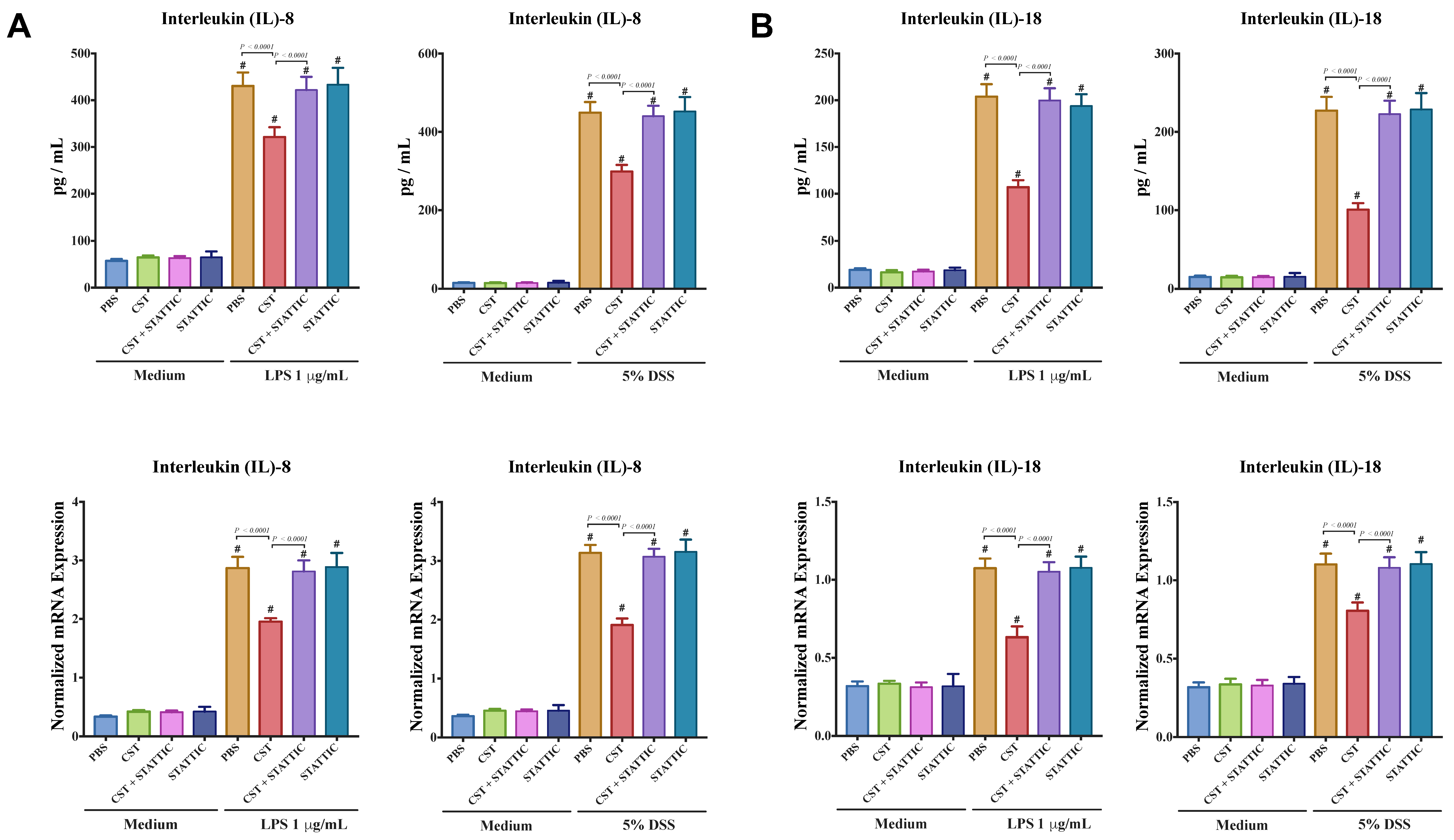
3.5. CST Maintains Gene Expression of TJ Proteins and Reduces IL-8 and IL-18 Release in LPS- and DSS-Stimulated Caco-2 Epithelial Cells through an Undetermined Mechanism Involving STAT3, But Not in Non-Inflammatory Conditions
3.6. CST Enhances the Functional Capacities of Colonic Epithelial Cells Including Epithelial Migration, Proliferation, Viability, & Oxidative Stress Resistance.
3.7. CST Decreases IL-6 Release in Acute DSS-Induced Colitis and DSS-and LPS-Stimulated Caco-2 Epithelial Cells, But Not in Control Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Buckley, A.; Turner, J.R. Cell biology of tight junction barrier regulation and mucosal disease. Cold Spring Harb. Perspect. Boil. 2018, 10, a029314. [Google Scholar] [CrossRef] [PubMed]
- Stio, M.; Retico, L.; Annese, V.; Bonanomi, A.G. Vitamin d regulates the tight-junction protein expression in active ulcerative colitis. Scand. J. Gastroenterol. 2016, 51, 1193–1199. [Google Scholar] [CrossRef] [PubMed]
- Maloy, K.J.; Powrie, F. Intestinal homeostasis and its breakdown in inflammatory bowel disease. Nature 2011, 474, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.Y.; Nighot, P.; Al-Sadi, R. Tight junctions and the intestinal barrier. In Physiology of the Gastrointestinal Tract, 6th ed.; Elsevier: Irvine, CA, USA, 2018; pp. 587–639. [Google Scholar]
- Rabbi, M.F.; Munyaka, P.M.; Eissa, N.; Metz-Boutigue, M.-H.; Khafipour, E.; Ghia, J.E. Human catestatin alters gut microbiota composition in mice. Front. Microbiol. 2017, 7, 2151. [Google Scholar] [CrossRef] [PubMed]
- Eissa, N.; Hussein, H.; Kermarrec, L.; Elgazzar, O.; Metz-Boutigue, M.-H.; Bernstein, C.N.; Ghia, J.-E. Chromofungin (chr: Chga47-66) is downregulated in persons with active ulcerative colitis and suppresses pro-inflammatory macrophage function through the inhibition of NF-κB signalling. Biochem. Pharmacol. 2017, 145, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Martini, E.; Krug, S.M.; Siegmund, B.; Neurath, M.F.; Becker, C. Mend your fences: The epithelial barrier and its relationship with mucosal immunity in inflammatory bowel disease. Cell. Mol. Gastroenterol. Hepatol. 2017, 4, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Waldner, M.J.; Neurath, M.F. Mechanisms of immune signalling in colitis-associated cancer. Cell. Mol. Gastroenterol. Hepatol. 2015, 1, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Neurath, M. New targets for mucosal healing and therapy in inflammatory bowel diseases. Mucosal Immunol. 2014, 7, 6–19. [Google Scholar] [CrossRef] [PubMed]
- Pickert, G.; Neufert, C.; Leppkes, M.; Zheng, Y.; Wittkopf, N.; Warntjen, M.; Lehr, H.-A.; Hirth, S.; Weigmann, B.; Wirtz, S. Stat3 links IL-22 signalling in intestinal epithelial cells to mucosal wound healing. J. Exp. Med. 2009, 206, 1465–1472. [Google Scholar] [CrossRef] [PubMed]
- Eissa, N.; Hussein, H.; Hendy, G.N.; Bernstein, C.N.; Ghia, J.-E. Chromogranin-a and its derived peptides and their pharmacological effects during intestinal inflammation. Biochem. Pharmacol. 2018, 152, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Eissa, N.; Kermarrec, L.; Ghia, J.-E. Neuroimmune mechanisms of cerebellar development and its developmental disorders: Bidirectional link between the immune system and nervous system. In Development of the Cerebellum from Molecular Aspects to Diseases; Springer: Cham, Switzerland, 2017; pp. 255–274. [Google Scholar]
- Eissa, N.; Ghia, J. Immunomodulatory effect of ghrelin in the intestinal mucosa. Neurogastroenterol. Motil. 2015, 27, 1519–1527. [Google Scholar] [CrossRef] [PubMed]
- Eissa, N.; Hussein, H.; Kermarrec, L.; Ali, A.Y.; Marshall, A.; Metz-Boutigue, M.-H.; Hendy, G.N.; Bernstein, C.N.; Ghia, J.-E. Chromogranin-a regulates macrophage function and the apoptotic pathway in murine dss colitis. J. Mol. Med. 2018, 96, 183–198. [Google Scholar] [CrossRef] [PubMed]
- Eissa, N.; Kermarrec, L.; Metz-Boutigue, M.-H.; Hendy, G.N.; Bernstein, C.N.; Ghia, J.-E. Chromofungin treatment promotes alternatively activated macrophages, suppresses classically activated macrophages and improves epithelial cell functions during colitis. Gastroenterology 2017, 152, S143. [Google Scholar] [CrossRef]
- Eissa, N.; Rabbi, M.; Bernstein, C.; Ghia, J. Chromofungin & pancreastatin co-regulate migration and functional plasticity of murine peritoneal macrophages. Neurogastroenterol. Motil. 2016, 28, 103–104. [Google Scholar]
- Rabbi, M.F.; Labis, B.; Metz-Boutigue, M.-H.; Bernstein, C.N.; Ghia, J.-E. Catestatin decreases macrophage function in two mouse models of experimental colitis. Biochem. Pharmacol. 2014, 89, 386–398. [Google Scholar] [CrossRef] [PubMed]
- Rabbi, M.F.; Eissa, N.; Munyaka, P.M.; Kermarrec, L.; Elgazzar, O.; Khafipour, E.; Bernstein, C.N.; Ghia, J.E. Reactivation of intestinal inflammation is suppressed by catestatin in a murine model of colitis via m1 macrophages and not the gut microbiota. Front. Immunol. 2017, 8, 985. [Google Scholar] [CrossRef] [PubMed]
- Eissa, N.; Rabbi, M.F.; Munyaka, P.M.; Khafipour, A.; Bernstein, C.N.; Ghia, J.-E. Mo1929 critical role of chromogranin-a on macrophage intrinsic apoptotic pathway in colitis: Human and animal studies. Gastroenterology 2016, 150, S819. [Google Scholar] [CrossRef]
- Eissa, N.; Hussein, H.; Wang, H.; Rabbi, M.F.; Bernstein, C.N.; Ghia, J.-E. Stability of reference genes for messenger rna quantification by real-time pcr in mouse dextran sodium sulfate experimental colitis. PLoS ONE 2016, 11, e0156289. [Google Scholar] [CrossRef] [PubMed]
- Eissa, N.; Kermarrec, L.; Hussein, H.; Bernstein, C.N.; Ghia, J.-E. Appropriateness of reference genes for normalizing messenger RNA in mouse 2,4-dinitrobenzene sulfonic acid (DNBS)-induced colitis using quantitative real time pcr. Sci. Rep. 2017, 7, 42427. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.Y.; Wu, X.; Eissa, N.; Hou, S.; Ghia, J.-E.; Murooka, T.T.; Banerji, V.; Johnston, J.B.; Lin, F.; Gibson, S.B. Distinct roles for phosphoinositide 3-kinases γ and δ in malignant B cell migration. Leukemia 2018, 32, 1958–1969. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente-Núñez, C.; Mansour, S.C.; Wang, Z.; Jiang, L.; Breidenstein, E.; Elliott, M.; Reffuveille, F.; Speert, D.P.; Reckseidler-Zenteno, S.L.; Shen, Y. Anti-biofilm and immunomodulatory activities of peptides that inhibit biofilms formed by pathogens isolated from cystic fibrosis patients. Antibiotics 2014, 3, 509–526. [Google Scholar] [CrossRef] [PubMed]
- Mansour, S.C.; Pena, O.M.; Hancock, R.E. Host defense peptides: Front-line immunomodulators. Trends Immunol. 2014, 35, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Bimczok, D.; Kao, J.Y.; Zhang, M.; Cochrun, S.; Mannon, P.; Peter, S.; Wilcox, C.M.; Mönkemüller, K.E.; Harris, P.R.; Grams, J.M. Human gastric epithelial cells contribute to gastric immune regulation by providing retinoic acid to dendritic cells. Mucosal Immunol. 2015, 8, 533–544. [Google Scholar] [CrossRef] [PubMed]
- Weber, C.R.; Turner, J.R. Dynamic modeling of the tight junction pore pathway. Ann. N. Y. Acad. Sci. 2017, 1397, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; de Haar, C.; Peppelenbosch, M.P.; van der Woude, J.C. New insights into the role of STAT3 in IBD. Inflamm. Bowel Dis. 2011, 18, 1177–1183. [Google Scholar] [CrossRef] [PubMed]
- Ludwiczek, O.; Kaser, A.; Novick, D.; Dinarello, C.A.; Rubinstein, M.; Tilg, H. Elevated systemic levels of free interleukin-18 (IL-18) in patients with crohn’s disease. Eur. Cytokine Netw. 2005, 16, 27–33. [Google Scholar] [PubMed]
- Eissa, N. The Role of Chromogranin-A in Inflammatory Bowel disease: Clinical & Experimental Approaches. Ph.D. Thesis, University of Manitoba, Winnipeg, MB, Canada, 2018. [Google Scholar]
- Nedoszytko, B.; Sokołowska-Wojdyło, M.; Ruckemann-Dziurdzińska, K.; Roszkiewicz, J.; Nowicki, R.J. Chemokines and cytokines network in the pathogenesis of the inflammatory skin diseases: Atopic dermatitis, psoriasis and skin mastocytosis. Adv. Dermatol. Allergol. Postȩpy Dermatol. I Alergol. 2014, 31, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Ying, W.; Mahata, S.; Bandyopadhyay, G.K.; Zhou, Z.; Wollam, J.; Vu, J.; Mayoral, R.; Chi, N.-W.; Webster, N.J.; Corti, A. Catestatin inhibits obesity-induced macrophage infiltration and inflammation in the liver and suppresses hepatic glucose production, leading to improved insulin sensitivity. Diabetes 2018, 67, 841–848. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.; Novick, D.; Kim, S.; Kaplanski, G. Interleukin-18 and il-18 binding protein. Front. Immunol. 2013, 4, 289. [Google Scholar] [CrossRef] [PubMed]
- Siegmund, B.; Fantuzzi, G.; Rieder, F.; Gamboni-Robertson, F.; Lehr, H.-A.; Hartmann, G.; Dinarello, C.A.; Endres, S.; Eigler, A. Neutralization of interleukin-18 reduces severity in murine colitis and intestinal ifn-γ and TNF-α production. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 281, R1264–R1273. [Google Scholar] [CrossRef] [PubMed]
- Ten Hove, T.; Corbaz, A.; Amitai, H.; Aloni, S.; Belzer, I.; Graber, P.; Drillenburg, P.; Van Deventer, S.J.; Chvatchko, Y.; Te Velde, A.A. Blockade of endogenous il-18 ameliorates tnbs-induced colitis by decreasing local tnf-α production in mice. Gastroenterology 2001, 121, 1372–1379. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Li, H.; Li, Y. Effects of bacillus subtilis on epithelial tight junctions of mice with inflammatory bowel disease. J. Interferon Cytokine Res. 2016, 36, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Maier, T.; Güell, M.; Serrano, L. Correlation of mrna and protein in complex biological samples. FEBS Lett. 2009, 583, 3966–3973. [Google Scholar] [CrossRef] [PubMed]
- Grivennikov, S.; Karin, E.; Terzic, J.; Mucida, D.; Yu, G.-Y.; Vallabhapurapu, S.; Scheller, J.; Rose-John, S.; Cheroutre, H.; Eckmann, L. IL-6 and STAT3 are required for survival of intestinal epithelial cells and development of colitis-associated cancer. Cancer Cell 2009, 15, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Bollrath, J.; Phesse, T.J.; von Burstin, V.A.; Putoczki, T.; Bennecke, M.; Bateman, T.; Nebelsiek, T.; Lundgren-May, T.; Canli, Ö.; Schwitalla, S. GP130-mediated STAT3 activation in enterocytes regulates cell survival and cell-cycle progression during colitis-associated tumorigenesis. Cancer Cell 2009, 15, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Corvinus, F.M.; Orth, C.; Moriggl, R.; Tsareva, S.A.; Wagner, S.; Pfitzner, E.B.; Baus, D.; Kaufman, R.; Huber, L.A.; Zatloukal, K. Persistent STAT3 activation in colon cancer is associated with enhanced cell proliferation and tumor growth. Neoplasia 2005, 7, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Zhao, E.; Zhang, D.; Basak, A.; Trudeau, V.L. New insights into granin-derived peptides: Evolution and endocrine roles. Gen. Comp. Endocrinol. 2009, 164, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Henriques, S.T.; Melo, M.N.; Castanho, M.A. Cell-penetrating peptides and antimicrobial peptides: How different are they? Biochem. J. 2006, 399, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Shooshtarizadeh, P.; Laventie, B.-J.; Colin, D.A.; Chich, J.-F.; Vidic, J.; de Barry, J.; Chasserot-Golaz, S.; Delalande, F.; Van Dorsselaer, A. Two chromogranin a-derived peptides induce calcium entry in human neutrophils by calmodulin-regulated calcium independent phospholipase A2. PLoS ONE 2009, 4, e4501. [Google Scholar] [CrossRef] [PubMed]
- Gry, M.; Rimini, R.; Strömberg, S.; Asplund, A.; Pontén, F.; Uhlén, M.; Nilsson, P. Correlations between rna and protein expression profiles in 23 human cell lines. BMC Genom. 2009, 10, 365. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward | Reverse |
|---|---|---|
| OCLDN | ACAAGCGGTTTTATCCAGAGTC | GTCATCCACAGGCGAAGTTAAT |
| TBP | CCCGAAACGCCGAATATAATCC | AATCAGTGCCGTGGTTCGTG |
| CLDN1 | AGGTGCTATCTGTTCAGTGATG | TGGCTGACTTTCCTTGTGTAG |
| ZO1 | CCAGCCTGCTAAACCTACTAAA | ATCTCTTGCTGCCAAACTATCT |
| IL8 | ACTGAGAGTGATTGAGAGTGGAC | AACCCTCTGCACCCAGTTTTC |
| IL18 | GCGTCACTACACTCAGCTAAT | GCGTCACTACACTCAGCTAAT |
| STAT3 | ACCAGCAGTATAGCCGCTTC | GCCACAATCCGGGCAATCT |
| IL6 | CCTGAACCTTCCAAAGATGGC | TTCACCAGGCAAGTCTCCTCA |
| Gene | Forward | Reverse |
|---|---|---|
| Il18 | GACTCTTGCGTCAACTTCAAGG | CAGGCTGTCTTTTGTCAACGA |
| Eef2 | TGTCAGTCATCGCCCATGTG | CATCCTTGCGAGTGTCAGTGA |
| Ocldn | TTGAAAGTCCACCTCCTTACAGA | CCGGATAAAAAGAGTACGCTGG |
| Cldn1 | GGGGACAACATCGTGACCG | AGGAGTCGAAGACTTTGCACT |
| Zo1 | GCCGCTAAGAGCACAGCAA | TCCCCACTCTGAAAATGAGGA |
| Il6 | TAGTCCTTCCTACCCCAATTTCC | TTGGTCCTTAGCCACTCCTTC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eissa, N.; Hussein, H.; Mesgna, R.; Bonin, S.; Hendy, G.N.; Metz-Boutigue, M.-H.; Bernstein, C.N.; Ghia, J.-E. Catestatin Regulates Epithelial Cell Dynamics to Improve Intestinal Inflammation. Vaccines 2018, 6, 67. https://doi.org/10.3390/vaccines6040067
Eissa N, Hussein H, Mesgna R, Bonin S, Hendy GN, Metz-Boutigue M-H, Bernstein CN, Ghia J-E. Catestatin Regulates Epithelial Cell Dynamics to Improve Intestinal Inflammation. Vaccines. 2018; 6(4):67. https://doi.org/10.3390/vaccines6040067
Chicago/Turabian StyleEissa, Nour, Hayam Hussein, Ruth Mesgna, Sandra Bonin, Geoffrey N. Hendy, Marie-Hélène Metz-Boutigue, Charles N. Bernstein, and Jean-Eric Ghia. 2018. "Catestatin Regulates Epithelial Cell Dynamics to Improve Intestinal Inflammation" Vaccines 6, no. 4: 67. https://doi.org/10.3390/vaccines6040067
APA StyleEissa, N., Hussein, H., Mesgna, R., Bonin, S., Hendy, G. N., Metz-Boutigue, M.-H., Bernstein, C. N., & Ghia, J.-E. (2018). Catestatin Regulates Epithelial Cell Dynamics to Improve Intestinal Inflammation. Vaccines, 6(4), 67. https://doi.org/10.3390/vaccines6040067