
Ethnic Comparisons of Spike-Specific CD4+ T Cells, Serological Responses, and Neutralizing Antibody Titers Against SARS-CoV-2 Variants



Abstract
1. Introduction
2. Materials and Methods
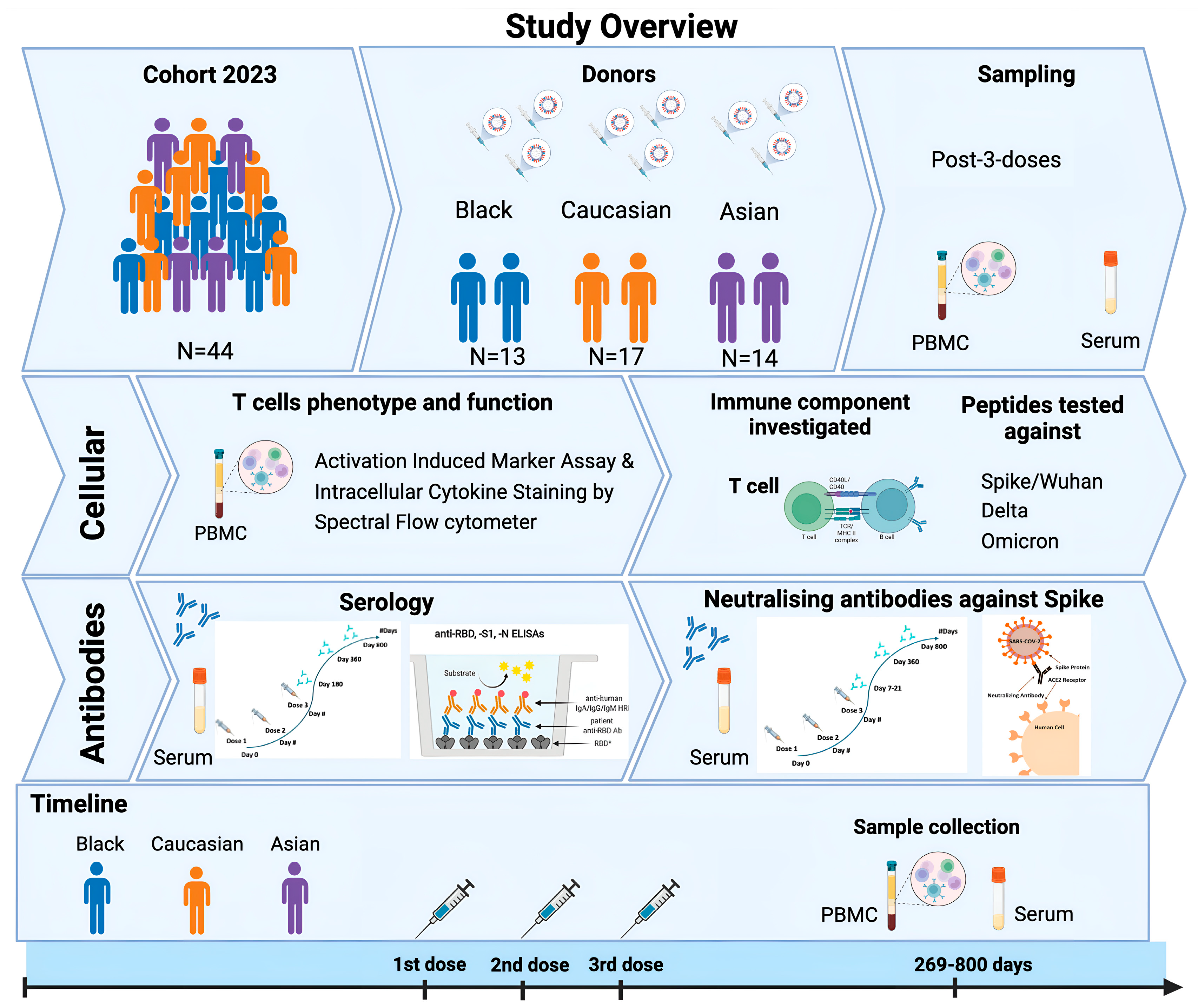
2.1. Study Participants
2.2. Whole Blood Processing and Enzyme-Linked Immunosorbent Assay (ELISA)
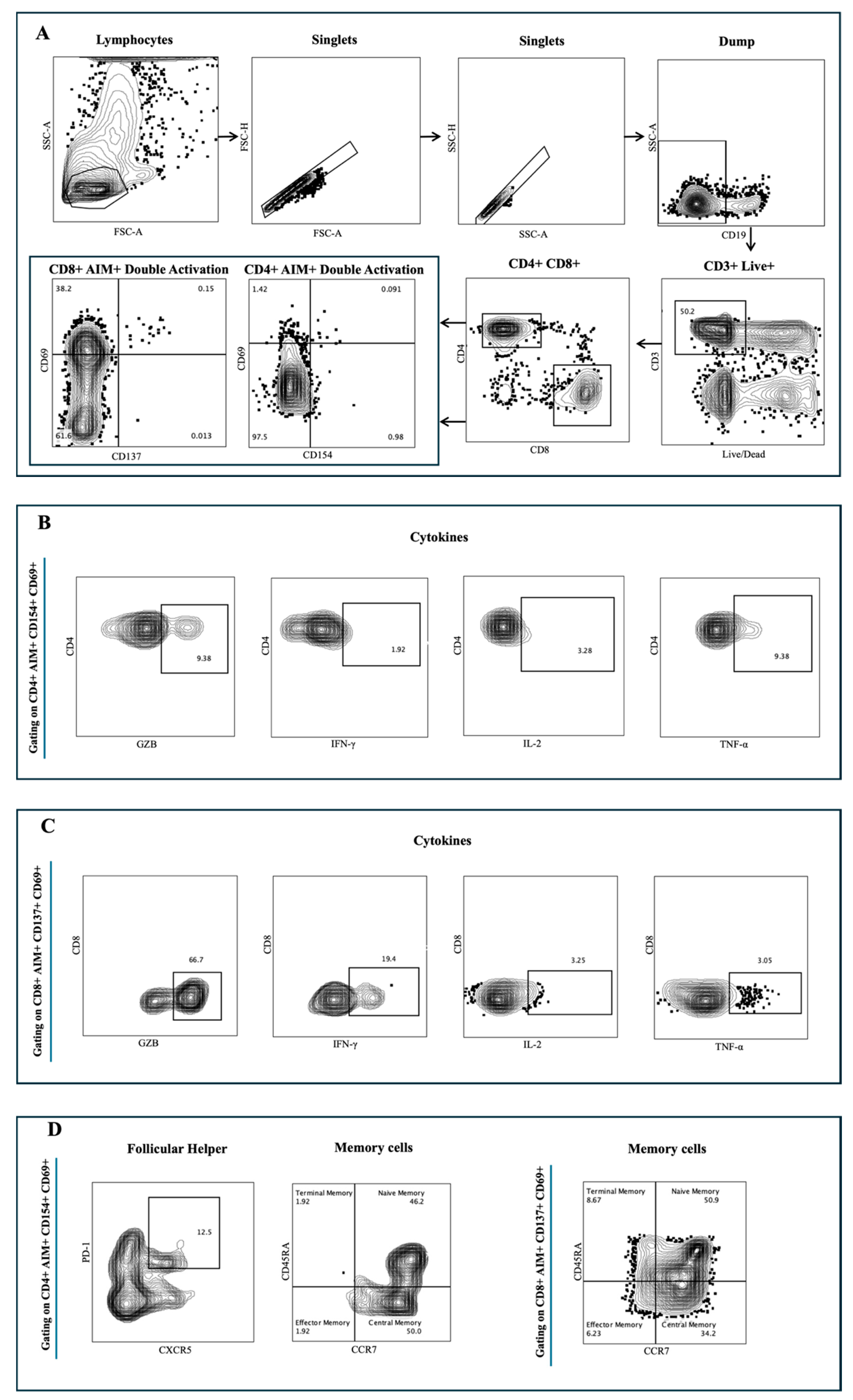
2.3. Flow Cytometry
2.4. SARS-CoV-2 Pseudolentivirus and Neutralization Assay
2.5. Statistical Analysis
2.6. Ethical Approval
3. Results
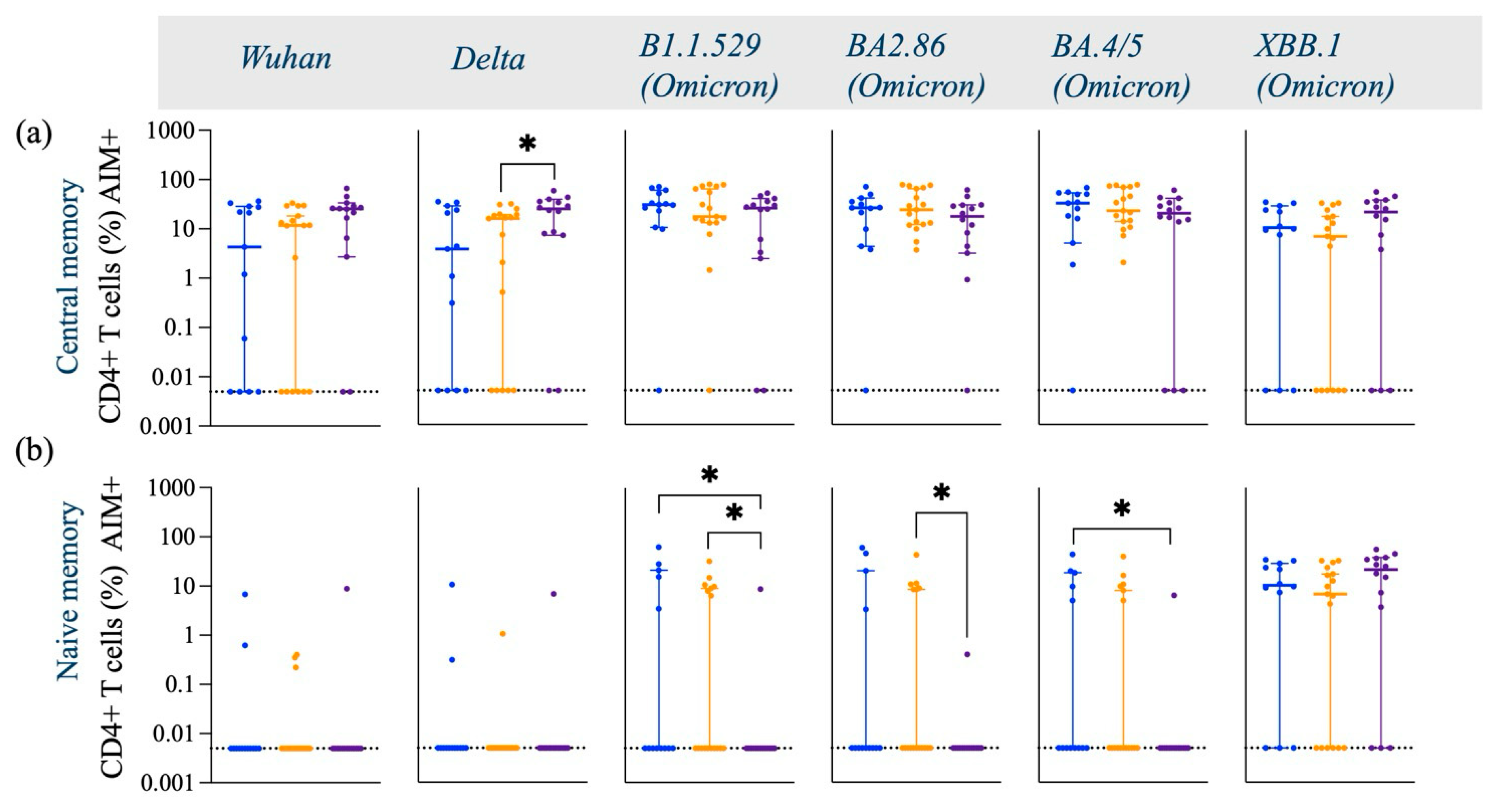
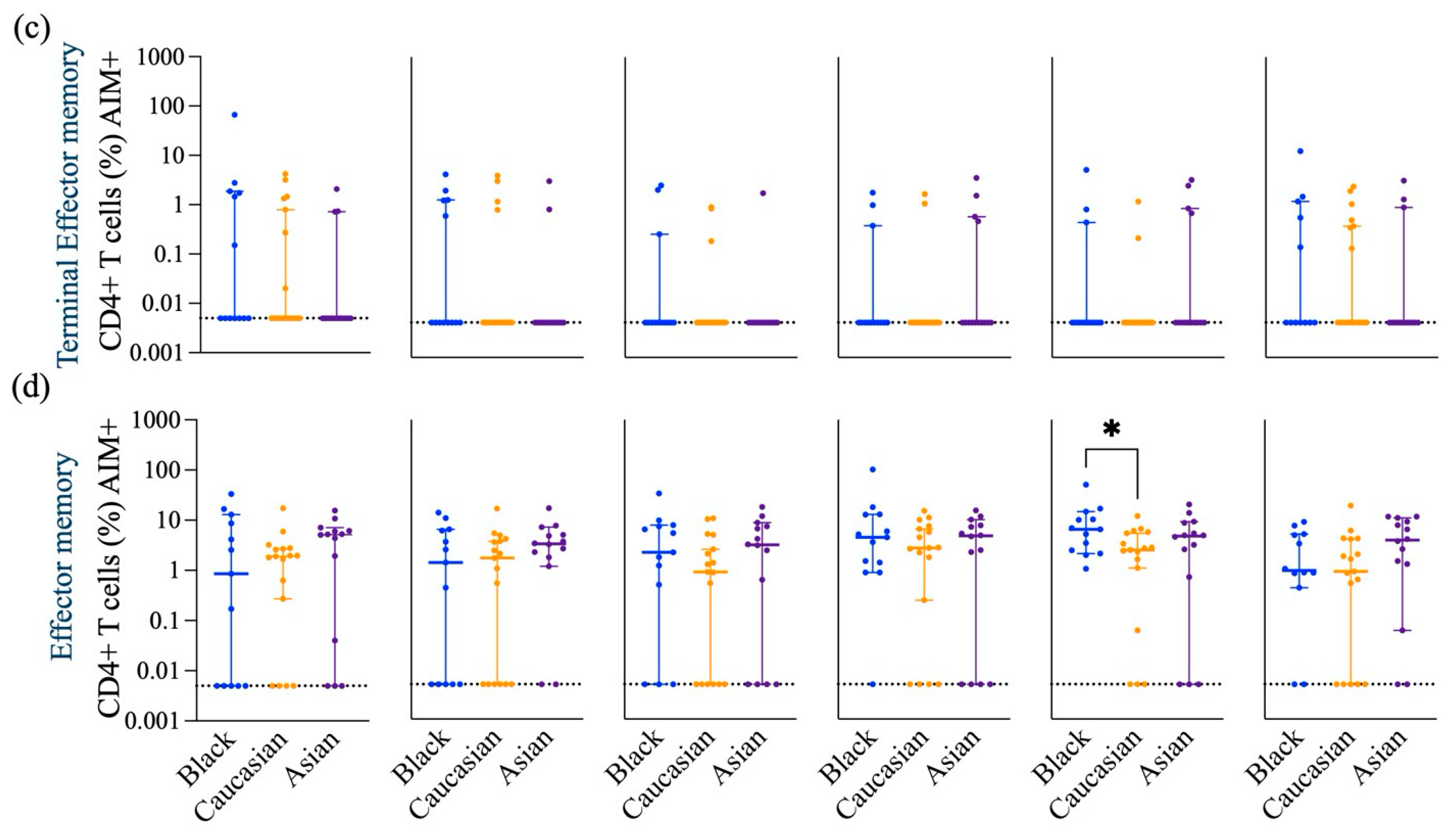
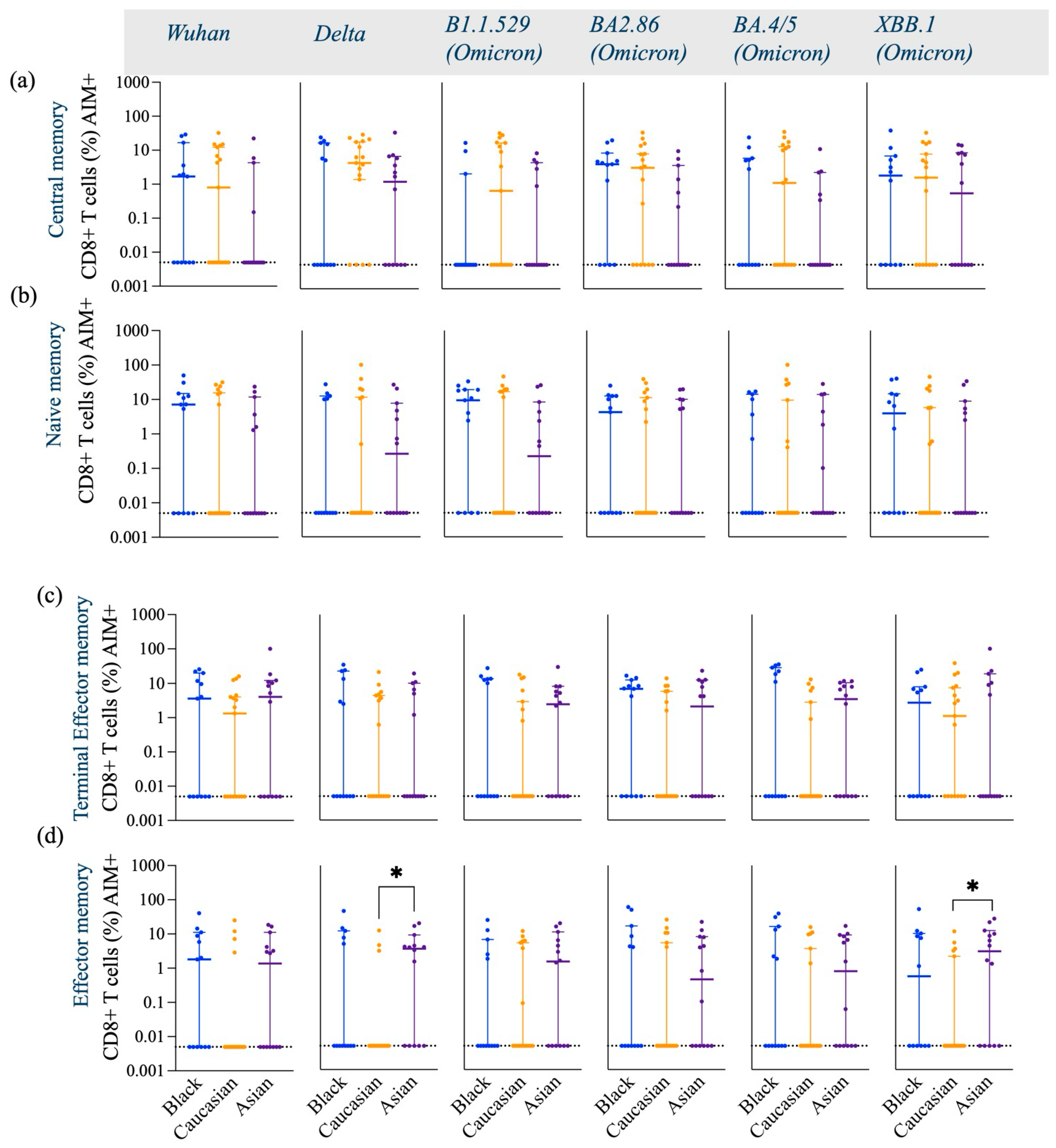
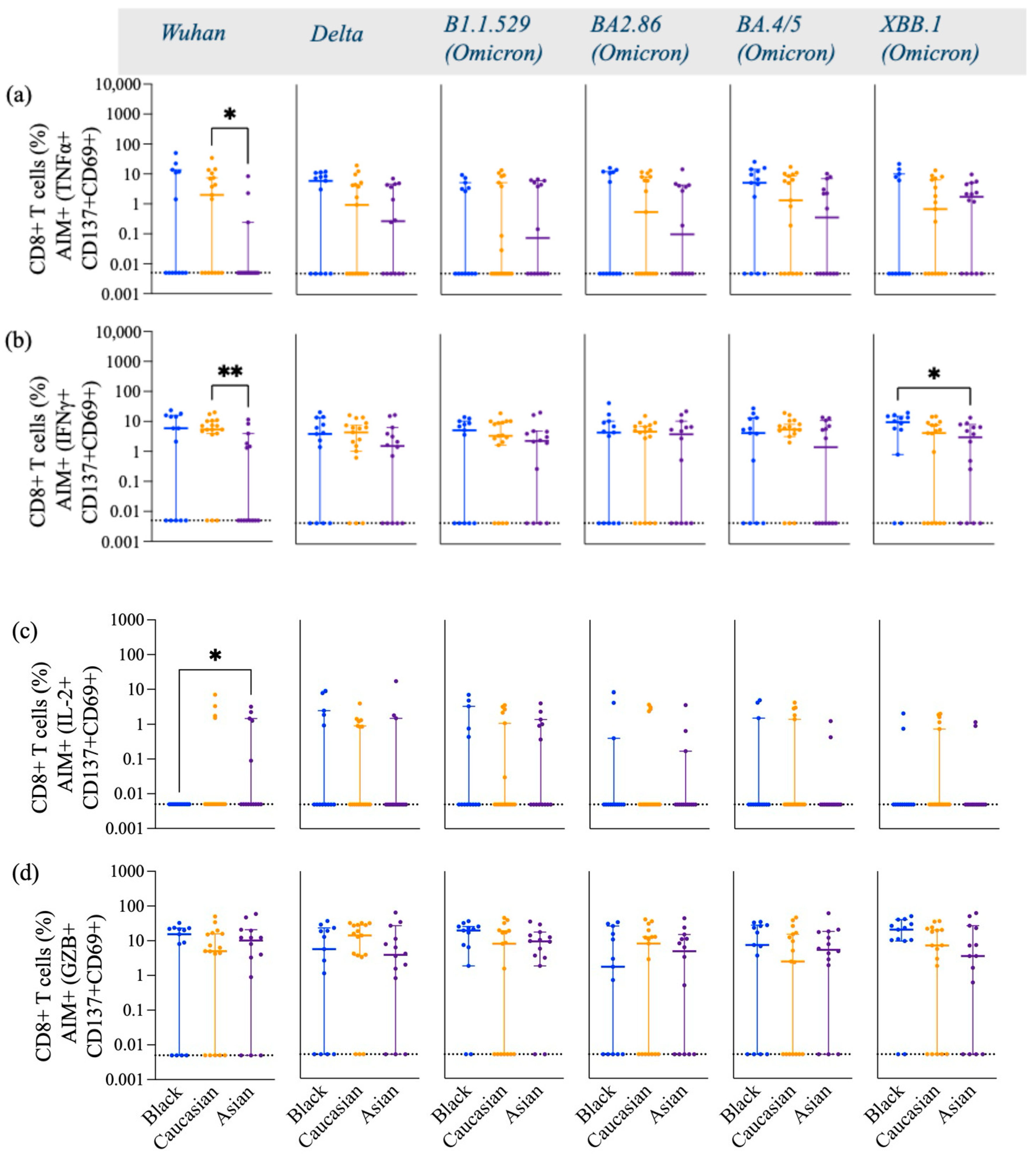
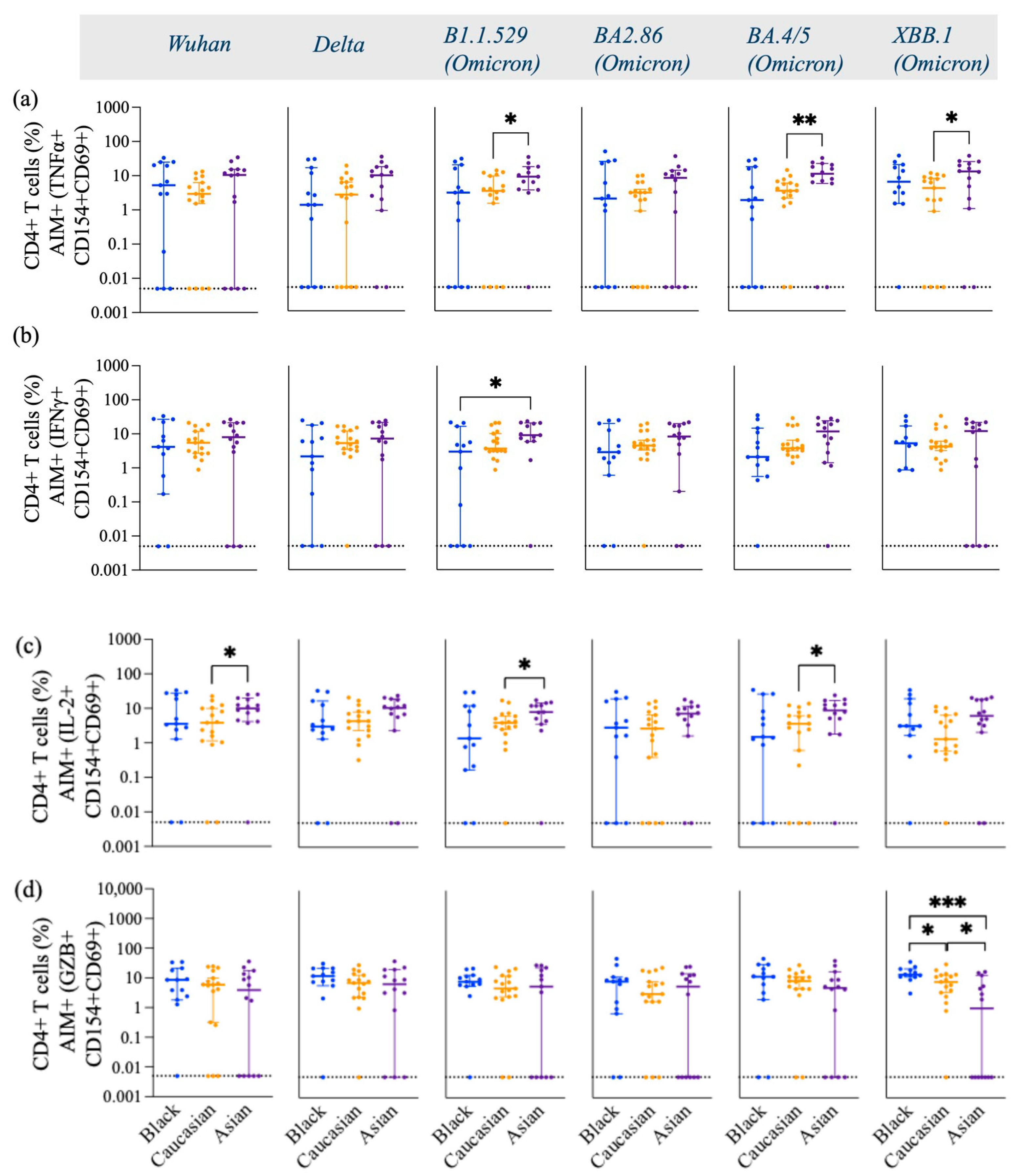
3.1. SARS-CoV-2 Spike-Specific AIM+ CD4+ and CD8+ T Cell Responses
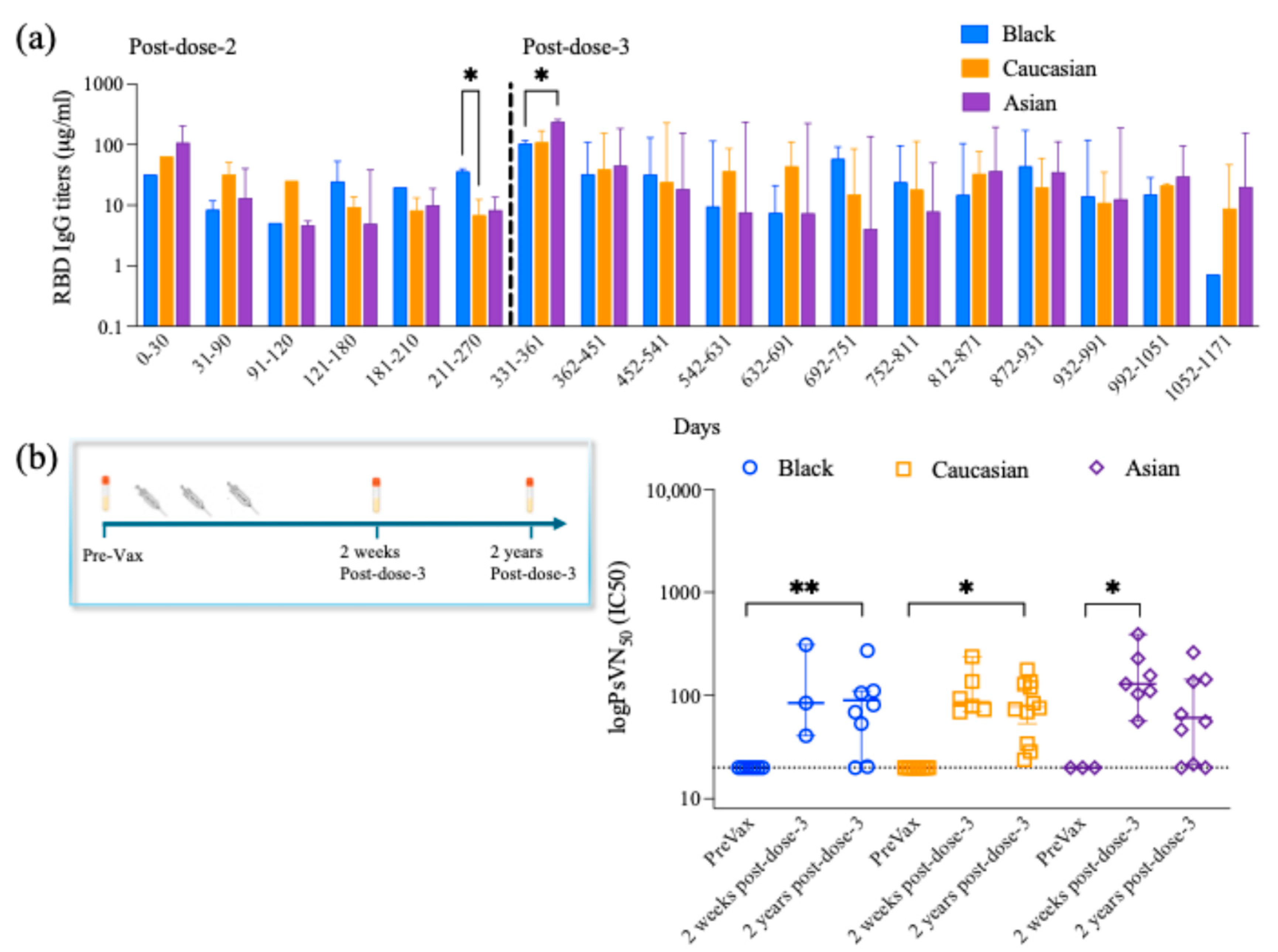
3.2. Longitudinal Anti-RBD IgG Antibody Titers
3.3. Long-Lived Protective Neutralizing Antibody Titers
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| RBD | Receptor-binding domain |
| Th1 | T helper type 1 |
| AIM | Activation-induced marker assays |
| ICS | Intracellular cytokine staining |
| PBMCs | Peripheral Blood Mononuclear Cells |
| cTfh | T follicular helper cells |
| GZB | Granzyme B |
| IFNγ | Interferon-gamma |
| TNFα | Tumor necrosis factor alpha |
| IL-2 | Interleukin 2 |
| DMEM | Dulbecco’s Modified Eagle Medium |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Race | Age | Gender | BTI | Pre-Immunity | Days Post-3-Dose | Vaccine Dose 1 | Vaccine Dose 2 | Vaccine Dose 3 |
|---|---|---|---|---|---|---|---|---|---|
| CCT-014 | Asian | 51 | F | no | no | 663 | Pfizer | Pfizer | Pfizer |
| CCT-008 | Asian | 34 | M | yes | no | 559 | Pfizer | Pfizer | Moderna |
| CVI-739 | Asian | 18 | F | no | no | 666 | Pfizer | Pfizer | Pfizer |
| CVI-753 | Asian | 20 | M | no | no | 599 | Pfizer | Pfizer | Pfizer |
| CVI-633 | Asian | 25 | F | yes | no | 597 | Pfizer | Pfizer | Pfizer |
| CVI-1019 | Asian | 27 | F | yes | no | 485 | Pfizer | Pfizer | Pfizer |
| CVI-914 | Asian | 29 | M | no | no | 408 | Astrazeneca | Astrazeneca | Astrazeneca |
| CVI-778 | Asian | 30 | F | no | no | 522 | Astrazeneca | Astrazeneca | Pfizer |
| CVI-224 | Asian | 34 | F | no | yes | 616 | Moderna | Moderna | Moderna |
| CVI-684 | Asian | 23 | F | yes | no | 549 | Pfizer | Pfizer | Pfizer |
| CVI-628 | Asian | 24 | M | yes | no | 800 | Pfizer | Pfizer | Pfizer |
| CVI-504 | Asian | 27 | F | no | no | 728 | Pfizer | Pfizer | Pfizer |
| STM-002 | Asian | 36 | F | yes | no | 665 | Pfizer | Pfizer | Pfizer |
| CVI-678 | Asian | 39 | F | no | no | 777 | Sinopharm CNBC bibp | Sinopharm CNBC bibp | Sinopharm CNBC bibp |
| CVI-1001 | Black | 45 | F | no | no | 436 | Pfizer | Pfizer | Pfizer |
| CVI-772 | Black | 21 | F | yes | yes | 583 | Pfizer | Pfizer | Pfizer |
| CVI-961 | Black | 66 | M | no | no | 446 | Pfizer | Pfizer | Pfizer |
| CTR-375 | Black | 20 | F | no | no | 534 | Moderna | Moderna | Moderna |
| CCT-025 | Black | 42 | M | no | no | 626 | Moderna | Moderna | Moderna |
| CVI-807 | Black | 55 | M | yes | no | 255 | Pfizer | Pfizer | Moderna |
| CVI-666 | Black | 30 | F | no | no | 269 | Pfizer | Pfizer | Pfizer Bivalent |
| CTR-365 | Black | 70 | F | no | no | 674 | Pfizer | Pfizer | Pfizer |
| CVI-987 | Black | 28 | F | no | no | 596 | Astrazeneca | Astrazeneca | Pfizer |
| CVI-1032 | Black | 33 | M | yes | no | 347 | Pfizer | Pfizer | Pfizer |
| LNC-003 | Black | 43 | F | yes | no | 504 | Moderna | Moderna | Moderna |
| P-201 | Black | 46 | F | no | no | >500 | Pfizer | Pfizer | Pfizer |
| CVI-996 | Black | 36 | F | yes | no | 683 | Pfizer | Pfizer | Pfizer |
| CCT-060 | Caucasian | 33 | M | no | no | 697 | Pfizer | Pfizer | Pfizer |
| CCT-052 | Caucasian | 34 | F | yes | no | 652 | Pfizer | Pfizer | Pfizer |
| CVI-926 | Caucasian | 19 | F | yes | no | 625 | Pfizer | Pfizer | Pfizer |
| CVI-540 | Caucasian | 27 | F | yes | no | 678 | Pfizer | Pfizer | Pfizer |
| CTR-257 | Caucasian | 25 | M | no | no | 642 | Pfizer | Pfizer | Pfizer |
| CVI-145 | Caucasian | 29 | M | yes | no | 670 | Moderna | Moderna | Pfizer |
| CCT-078 | Caucasian | 50 | F | yes | no | 397 | Pfizer | Pfizer | Pfizer |
| CVI-845 | Caucasian | 25 | F | yes | no | 620 | Moderna | Moderna | Pfizer |
| CVI-820 | Caucasian | 25 | F | no | no | 494 | Pfizer | Pfizer | Pfizer |
| CTR-257 | Caucasian | 25 | M | no | no | 558 | Pfizer | Pfizer | Pfizer |
| CCT-031 | Caucasian | 48 | F | no | no | 746 | Pfizer | Pfizer | Pfizer |
| CVI-157 | Caucasian | 29 | F | no | yes | 495 | Pfizer | Pfizer | Pfizer |
| CCT-050 | Caucasian | 76 | M | yes | yes | 698 | Moderna | Moderna | Moderna |
| CCT-002 | Caucasian | 25 | M | no | no | 675 | Moderna | Moderna | Moderna |
| CCT-132 | Caucasian | 32 | M | yes | no | 658 | Moderna | Moderna | Pfizer |
| CCT-031 | Caucasian | 48 | F | no | no | 700 | Pfizer | Pfizer | Pfizer |
| CCT-085 | Caucasian | 48 | F | yes | no | 506 | Moderna | Moderna | Moderna |
| CCF1-020 | Caucasian | 50 | F | no | no | 707 | Pfizer | Pfizer | Pfizer |
| Antigen | Clone | Fluorophore | Company | Panel | Catalogue Nb. | Dilution |
|---|---|---|---|---|---|---|
| Viability dye | - | Zombie NIR | Biolegend | surface | 423105 | 1:1,250 |
| CD8 | SK1 | SB550 | Biolegend | surface | 344760 | 1:125 |
| PD1 | EH12.1 | BB515 | BD Bioscience | surface | 564494 | 1:10 |
| CD45RA | HI100 | PerCP | Biolegend | surface | 304156 | 1:25 |
| CD154 (CD40L) | 24-31 | APC | Biolegend | surface | 310810 | 1:50 |
| CCR7 | 2-L1-A | APCR700 | Biolegend | surface | 566766 | 1:25 |
| CD19 | HIB19 | APCEF780 | Invitrogen | surface | 50-161-52 | 1:25 |
| CD14 | 61D3 | APCEF780 | Invitrogen | surface | 47-0149-42 | 1:10 |
| CD3 | UCHT1 | BV570 | Biolegend | surface | 300436 | 1:10 |
| CXCR5 | J252D4 | PE-DAZZLE | Biolegend | surface | 356928 | 1:50 |
| CD4 | SK3 | cFluor YG 584 | Cytek | surface | R7-20041 | 1:100 |
| CD134 (OX40) | Ber-ACT35 | PECY7 | Biolegend | surface | 350012 | 1:250 |
| IL2 | MQ1-17H12 | PERCP-EF 710 | Invitrogen | intracellular | 50-161-00 | 1:50 |
| IFNG | 4SB3 | BV711 | Biolegend | intracellular | 502540 | 1:25 |
| TNF | MAB11 | BV785 | Biolegend | intracellular | 502948 | 1:25 |
| GZB | QA16A02 | PE-CY5 | Biolegend | intracellular | 372226 | 1:100 |
| CD137 | 4B4-1 | BV750 | Biolegend | intracellular | 309844 | 1:50 |
| CD69 | FN50 | BV510 | Biolegend | intracellular | 310936 | 1:25 |
References
- Zhang, B.; Fong, Y.; Fintzi, J.; Chu, E.; Janes, H.E.; Kenny, A.; Carone, M.; Benkeser, D.; van der Laan, L.W.P.; Deng, W.; et al. Omicron COVID-19 immune correlates analysis of a third dose of mRNA-1273 in the COVE trial. Nat. Commun. 2024, 15, 7954. [Google Scholar] [CrossRef] [PubMed]
- Tarke, A.; Coelho, C.H.; Zhang, Z.; Dan, J.M.; Yu, E.D.; Methot, N.; Bloom, N.I.; Goodwin, B.; Phillips, E.; Mallal, S.; et al. SARS-CoV-2 vaccination induces immunological T cell memory able to cross-recognize variants from Alpha to Omicron. Cell 2022, 185, 847–859 e11. [Google Scholar] [CrossRef]
- Wang, L.; Nicols, A.; Turtle, L.; Richter, A.; Duncan, C.J.; Dunachie, S.J.; Klenerman, P.; Payne, R.P. T cell immune memory after covid-19 and vaccination. BMJ Med. 2023, 2, e000468. [Google Scholar] [CrossRef]
- Calder, P.C.; Carr, A.C.; Gombart, A.F.; Eggersdorfer, M. Optimal Nutritional Status for a Well-Functioning Immune System Is an Important Factor to Protect against Viral Infections. Nutrients 2020, 12, 1181. [Google Scholar] [CrossRef]
- Maggini, S.; Pierre, A.; Calder, P.C. Immune Function and Micronutrient Requirements Change over the Life Course. Nutrients 2018, 10, 1531. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.A.; Nazareth, J.; Jarkhi, A.; Pan, D.; Das, M.; Logan, N.; Scott, S.; Bryant, L.; Abeywickrama, N.; Adeoye, O.; et al. Ethnic differences in cellular and humoral immune responses to SARS-CoV-2 vaccination in UK healthcare workers: A cross-sectional analysis. eClinicalMedicine 2023, 58, 101926. [Google Scholar] [CrossRef]
- Magesh, S.; John, D.; Li, W.T.; Li, Y.; Mattingly-App, A.; Jain, S.; Chang, E.Y.; Ongkeko, W.M. Disparities in COVID-19 Outcomes by Race, Ethnicity, and Socioeconomic Status: A Systematic-Review and Meta-analysis. JAMA Netw Open. 2021, 4, e2134147. [Google Scholar] [CrossRef]
- Center FDA. FDA Approves and Authorizes Updated mRNA COVID-19 Vaccines to Better Protect Against Currently Circulating Variants: FDA News Release. 2024. Available online: https://www.fda.gov/news-events/press-announcements/fda-approves-and-authorizes-updated-mrna-covid-19-vaccines-better-protect-against-currently (accessed on 4 May 2025).
- CDC. Center for Disease Control and Prevention: COVID Data Tracker Atlanta, GA: U.S. Department of Health and Human Services; 2024 [updated 2024, December 27]. Available online: https://covid.cdc.gov/covid-data-tracker (accessed on 4 May 2025).
- Wahl, I.; Wardemann, H. Sterilizing immunity: Understanding COVID-19. Immunity 2022, 55, 2231–2235. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Zhang, Q.; Gu, X.; Ren, L.; Huang, T.; Li, Y.; Zhang, H.; Liu, Y.; Zhong, J.; Wang, X.; et al. Durability and cross-reactive immune memory to SARS-CoV-2 in individuals 2 years after recovery from COVID-19: A longitudinal cohort study. Lancet Microbe 2024, 5, e24–e33. [Google Scholar] [CrossRef]
- Moss, P. The T cell immune response against SARS-CoV-2. Nat. Immunol. 2022, 23, 186–193. [Google Scholar] [CrossRef]
- Sette, A.; Sidney, J.; Crotty, S. T Cell Responses to SARS-CoV-2. Annu. Rev. Immunol. 2023, 41, 343–373. [Google Scholar] [CrossRef]
- Forgacs, D.; Jang, H.; Abreu, R.B.; Hanley, H.B.; Gattiker, J.L.; Jefferson, A.M.; Ross, T.M. SARS-CoV-2 mRNA Vaccines Elicit Different Responses in Immunologically Naive and Pre-Immune Humans. Front. Immunol. 2021, 12, 728021. [Google Scholar] [CrossRef] [PubMed]
- Forgacs, D.; Silva-Moraes, V.; Sautto, G.A.; Hanley, H.B.; Gattiker, J.L.; Jefferson, A.M.; Kolhe, R.; Ross, T.M. The Effect of Waning on Antibody Levels and Memory B Cell Recall following SARS-CoV-2 Infection or Vaccination. Vaccines 2022, 10, 696. [Google Scholar] [CrossRef] [PubMed]
- Silva-Moraes, V.; Souquette, A.; Sautto, G.A.; Paciello, I.; Antonelli, G.; Andreano, E.; Rappuoli, R.; Teixeira-Carvalho, A.; Ross, T.M. Prior SARS-CoV-2 Infection Enhances Initial mRNA Vaccine Response with a Lower Impact on Long-Term Immunity. Immunohorizons 2023, 7, 635–651. [Google Scholar] [CrossRef]
- Ecker, J.W.; Kirchenbaum, G.A.; Pierce, S.R.; Skarlupka, A.L.; Abreu, R.B.; Cooper, R.E.; Taylor-Mulneix, D.; Ross, T.M.; Sautto, G.A. High-Yield Expression and Purification of Recombinant Influenza Virus Proteins from Stably-Transfected Mammalian Cell Lines. Vaccines 2020, 8, 462. [Google Scholar] [CrossRef] [PubMed]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501 e15. [Google Scholar] [CrossRef]
- Mateus, J.; Grifoni, A.; Tarke, A.; Sidney, J.; Ramirez, S.I.; Dan, J.M.; Burger, Z.C.; Rawlings, S.A.; Smith, D.M.; Phillips, E.; et al. Selective and cross-reactive SARS-CoV-2 T cell epitopes in unexposed humans. Science 2020, 370, 89–94. [Google Scholar] [CrossRef]
- Rydyznski Moderbacher, C.; Ramirez, S.I.; Dan, J.M.; Grifoni, A.; Hastie, K.M.; Weiskopf, D.; Belanger, S.; Abbott, R.K.; Kim, C.; Choi, J.; et al. Antigen-Specific Adaptive Immunity to SARS-CoV-2 in Acute COVID-19 and Associations with Age and Disease Severity. Cell 2020, 183, 996–1012 e19. [Google Scholar] [CrossRef]
- Tarke, A.; Ramezani-Rad, P.; Alves Pereira Neto, T.; Lee, Y.; Silva-Moraes, V.; Goodwin, B.; Bloom, N.; Siddiqui, L.; Avalos, L.; Frazier, A.; et al. SARS-CoV-2 breakthrough infections enhance T cell response magnitude, breadth, and epitope repertoire. Cell Rep. Med. 2024, 5, 101583. [Google Scholar] [CrossRef]
- Ferrara, F.; Temperton, N. Pseudotype Neutralization Assays: From Laboratory Bench to Data Analysis. Methods Protoc. 2018, 1, 8. [Google Scholar] [CrossRef]
- Bellusci, L.; Grubbs, G.; Zahra, F.T.; Forgacs, D.; Golding, H.; Ross, T.M.; Khurana, S. Antibody affinity and cross-variant neutralization of SARS-CoV-2 Omicron BA.1, BA.2 and BA.3 following third mRNA vaccination. Nat. Commun. 2022, 13, 4617. [Google Scholar] [CrossRef] [PubMed]
- Binayke, A.; Zaheer, A.; Vishwakarma, S.; Singh, S.; Sharma, P.; Chandwaskar, R.; Gosain, M.; Raghavan, S.; Murugesan, D.R.; Kshetrapal, P.; et al. A quest for universal anti-SARS-CoV-2 T cell assay: Systematic review, meta-analysis, and experimental validation. NPJ Vaccines. 2024, 9, 3. [Google Scholar] [CrossRef]
- Piccoli, L.; Park, Y.J.; Tortorici, M.A.; Czudnochowski, N.; Walls, A.C.; Beltramello, M.; Silacci-Fregni, C.; Pinto, D.; Rosen, L.E.; Bowen, J.E.; et al. Mapping Neutralizing and Immunodominant Sites on the SARS-CoV-2 Spike Receptor-Binding Domain by Structure-Guided High-Resolution Serology. Cell 2020, 183, 1024–1042 e21. [Google Scholar] [CrossRef] [PubMed]
- Wec, A.Z.; Wrapp, D.; Herbert, A.S.; Maurer, D.P.; Haslwanter, D.; Sakharkar, M.; Jangra, R.K.; Dieterle, M.E.; Lilov, A.; Huang, D.; et al. Broad neutralization of SARS-related viruses by human monoclonal antibodies. Science 2020, 369, 731–736. [Google Scholar] [CrossRef]
- Wrapp, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.L.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 2020, 367, 1260–1263. [Google Scholar] [CrossRef]
- Chen, X.; Meng, X.; Wu, Q.; Lim, W.W.; Xin, Q.; Cowling, B.J.; Meng, W.; Yu, H.; Covasa, D.T. Assessment of neutralizing antibody response as a correlate of protection against symptomatic SARS-CoV-2 infections after administration of two doses of the CoronaVac inactivated COVID-19 vaccine: A phase III randomized controlled trial. J. Infect. 2024, 89, 106315. [Google Scholar] [CrossRef]
- Almendro-Vázquez, P.; Laguna-Goya, R.; Paz-Artal, E. Defending against SARS-CoV-2: The T cell perspective. Front. Immunol. 2023, 14, 1107803. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.Y.; Tang, Y.Q.; Zhang, Z.B.; Liu, J.; Zhuang, Y.X.; Li, T. COVID-19: From immune response to clinical intervention. Precis Clin. Med. 2024, 7, pbae015. [Google Scholar] [CrossRef]
- Snell, L.M.; Osokine, I.; Yamada, D.H.; De la Fuente, J.R.; Elsaesser, H.J.; Brooks, D.G. Overcoming CD4 Th1 Cell Fate Restrictions to Sustain Antiviral CD8 T Cells and Control Persistent Virus Infection. Cell Rep. 2016, 16, 3286–3296. [Google Scholar] [CrossRef]
- Mehta, A.K.; Gracias, D.T.; Croft, M. TNF activity and T cells. Cytokine 2018, 101, 14–18. [Google Scholar] [CrossRef]
- Soudja, S.M.; Chandrabos, C.; Yakob, E.; Veenstra, M.; Palliser, D.; Lauvau, G. Memory-T-cell-derived interferon-gamma instructs potent innate cell activation for protective immunity. Immunity 2014, 40, 974–988. [Google Scholar] [CrossRef] [PubMed]
- Ekkens, M.J.; Shedlock, D.J.; Jung, E.; Troy, A.; Pearce, E.L.; Shen, H.; Pearce, E.J. Th1 and Th2 cells help CD8 T-cell responses. Infect. Immun. 2007, 75, 2291–2296. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, P.A.S.; Assone, T.; Prates, G.; Tedeschi, M.R.M.; Fonseca, L.A.M.; Casseb, J. The role of IFN-gamma production during retroviral infections: An important cytokine involved in chronic inflammation and pathogenesis. Rev. Inst. Med. Trop. Sao. Paulo. 2022, 64, e64. [Google Scholar] [CrossRef]
- Hawerkamp, H.C.; Dyer, A.H.; Patil, N.D.; McElheron, M.; O’dowd, N.; O’doherty, L.; Mhaonaigh, A.U.; George, A.M.; O’halloran, A.M.; Reddy, C.; et al. Characterisation of the pro-inflammatory cytokine signature in severe COVID-19. Front. Immunol. 2023, 14, 1170012. [Google Scholar] [CrossRef]
- Grimm, L.; Onyeukwu, C.; Kenny, G.; Parent, D.; Fu, J.; Dhingra, S.; Yang, E.; Moy, J.; Utz, P.; Tracy, R.; et al. Immune Dysregulation in Acute SARS-CoV-2 Infection. Pathog. Immun. 2022, 7, 143–170. [Google Scholar] [CrossRef]
- Zanza, C.; Romenskaya, T.; Manetti, A.C.; Franceschi, F.; La Russa, R.; Bertozzi, G.; Maiese, A.; Savioli, G.; Volonnino, G.; Longhitano, Y. Cytokine Storm in COVID-19: Immunopathogenesis and Therapy. Medicina 2022, 58, 144. [Google Scholar] [CrossRef] [PubMed]
- Belik, M.; Liedes, O.; Vara, S.; Haveri, A.; Pöysti, S.; Kolehmainen, P.; Maljanen, S.; Huttunen, M.; Reinholm, A.; Lundberg, R.; et al. Persistent T cell-mediated immune responses against Omicron variants after the third COVID-19 mRNA vaccine dose. Front. Immunol. 2023, 14, 1099246. [Google Scholar] [CrossRef]
- Sereejav, E.; Sandagdorj, A.; Bazarjav, P.; Ganbold, S.; Enkhtuvshin, A.; Tsedenbal, N.; Namuuntsetseg, B.; Chimedregzen, K.; Badarch, D.; Otgonbayar, D.; et al. Antibody responses to mRNA versus non-mRNA COVID vaccines among the Mongolian population. IJID Reg. 2023, 8, 1–8. [Google Scholar] [CrossRef]
- Maher, S.; El Assaly, N.M.; Aly, D.M.; Atta, S.; Fteah, A.M.; Badawi, H.; Zahran, M.Y.; Kamel, M. Comparative study of neutralizing antibodies titers in response to different types of COVID-19 vaccines among a group of egyptian healthcare workers. Virol. J. 2024, 21, 277. [Google Scholar] [CrossRef]


| Characteristics | Black (N = 13) | Caucasian (N = 17) | Asian (N = 14) |
|---|---|---|---|
| Age (years) | |||
| 18–30 | 4 | 8 | 9 |
| 31–50 | 6 | 8 | 4 |
| >51 | 3 | 1 | 1 |
| Gender | |||
| Males | 4 | 6 | 4 |
| Females | 9 | 11 | 10 |
| Vaccine type | |||
| Pfizer | 10 | 11 | 10 |
| Moderna | 4 | 6 | 1 |
| AstraZeneca | 1 | 0 | 2 |
| BBIBP-CorV (Sinopharm) | 0 | 0 | 1 |
| Pfizer Bivalent | 1 | 0 | 0 |
| Pre-Immunity | |||
| Naïve | 12 | 15 | 13 |
| Pre-Immune | 1 | 2 | 1 |
| Breakthrough Infections | |||
| Positive PCR or self-reported/RBD titer | 5 | 9 | 6 |
| Pre-existing health conditions | |||
| Asthma | 1 | 4 | 0 |
| Hypothyroidism | 0 | 1 | 1 |
| Wolff-Parkinson-White syndrome | 0 | 1 | 0 |
| Chronic Kidney disease | 0 | 1 | 0 |
| Hypertension | 3 | 0 | 0 |
| Hypercholesteraemia | 0 | 0 | 2 |
| Hypertriglyceridemia | 0 | 0 | 1 |
| Prescribed Medications | |||
| Levothyroxine | 0 | 1 | 0 |
| Albuterol | 1 | 0 | 0 |
| Paxlovid | 1 | 0 | 0 |
| Wuhan | Delta | B1.1.529 | BA2.86 | BA4/5 | XBB.1 | ||
|---|---|---|---|---|---|---|---|
| Black participants | |||||||
| RBD IgG Ab titers × cTfh | Spearman’s r | 0.732 | 0.102 | 0.252 | 0.382 | 0.189 | 0.223 |
| p value | 0.0098 ** | 0.757 | 0.430 | 0.221 | 0.557 | 0.506 | |
| RBD IgG Ab titers × CD4+ Activated T cells | Spearman’s r | −0.427 | −0.280 | −0.308 | −0.392 | −0.270 | −0.219 |
| p value | 0.169 | 0.379 | 0.331 | 0.207 | 0.394 | 0.515 | |
| Neutralizing Ab titers × RBD IgG Ab titers | Spearman’s r | 0.741 | - | - | - | - | - |
| p value | 0.0078 ** | - | - | - | - | - | |
| Neutralizing Ab titers × cTfh | Spearman’s r | 0.644 | −0.345 | −0.005 | 0.406 | 0.341 | 0.220 |
| p value | 0.021 * | 0.248 | 0.993 | 0.170 | 0.252 | 0.487 | |
| Neutralizing Ab titers × CD4+ Activated T cells | Spearman’s r | −0.291 | −0.137 | −0.275 | −0.160 | −0.198 | −0.053 |
| p value | 0.334 | 0.656 | 0.363 | 0.600 | 0.513 | 0.873 | |
| Caucasian participants | |||||||
| RBD IgG Ab titers × cTfh | Spearman’s r | 0.312 | −0.030 | 0.167 | 0.213 | 0.275 | 0.289 |
| p value | 0.208 | 0.906 | 0.507 | 0.396 | 0.270 | 0.245 | |
| RBD IgG Ab titers × CD4+ Activated T cells | Spearman’s r | 0.174 | 0.103 | 0.140 | 0.108 | 0.159 | 0.061 |
| p value | 0.503 | 0.694 | 0.592 | 0.678 | 0.540 | 0.817 | |
| Neutralizing Ab titers × RBD IgG Ab titers | Spearman’s r | 0.800 | - | - | - | - | - |
| p value | 0.0006 *** | - | - | - | - | - | |
| Neutralizing Ab titers × cTfh | Spearman’s r | 0.371 | 0.093 | 0.416 | 0.371 | 0.384 | 0.330 |
| p value | 0.173 | 0.742 | 0.123 | 0.175 | 0.158 | 0.229 | |
| Neutralizing Ab titers × CD4+ Activated T cells | Spearman’s r | −0.071 | −0.168 | −0.111 | −0.229 | −0.218 | −0.168 |
| p value | 0.803 | 0.549 | 0.695 | 0.409 | 0.434 | 0.549 | |
| Asian participants | |||||||
| RBD IgG Ab titers × cTfh | Spearman’s r | 0.319 | 0.247 | 0.332 | 0.327 | −0.189 | −0.024 |
| p value | 0.246 | 0.372 | 0.246 | 0.233 | 0.498 | 0.934 | |
| RBD IgG Ab titers × CD4+ Activated T cells | Spearman’s r | 0.304 | 0.206 | 0.370 | 0.202 | 0.302 | 0.075 |
| p value | 0.271 | 0.459 | 0.193 | 0.468 | 0.271 | 0.790 | |
| Neutralizing Ab titers × RBD IgG Ab titers | Spearman’s r | 0.792 | - | - | - | - | - |
| p value | 0.0012 ** | - | - | - | - | - | |
| Neutralizing Ab titers × cTfh | Spearman’s r | 0.298 | 0.093 | 0.363 | 0.138 | −0.018 | −0.081 |
| p value | 0.297 | 0.748 | 0.221 | 0.634 | 0.954 | 0.782 | |
| Neutralizing Ab titers × CD4+ Activated T cells | Spearman’s r | 0.425 | 0.351 | 0.432 | 0.256 | 0.256 | 0.167 |
| p value | 0.131 | 0.217 | 0.141 | 0.374 | 0.373 | 0.565 |
| Parameter | Black Group | Caucasian Group | Asian Group |
|---|---|---|---|
| Activated CD4+ T Cells | ↑ (comparable to other groups) | ↑ (comparable to other groups) | ↓ as compared to Caucasian (p < 0.05) in response to Omicron subvariants |
| Activated CD8+ T Cells | ↑ (comparable to other groups) | ↑ (comparable to other groups) | ↑ (comparable to other groups) |
| TNF-α secreting Activated CD4+ T Cells | ↑ (comparable to other groups) | ↓ as compared to Asian (p < 0.05) in response to Omicron subvariant | ↑ as compared to Caucasian (p < 0.05, p < 0.01) in response to Omicron subvariants |
| IFN-γ secreting Activated CD4+ T Cells | ↓ as compared to Asian (p < 0.05) in response to Omicron subvariant | ↑ (comparable to other groups) | ↑ as compared to Black (p < 0.05) in response to Omicron subvariant |
| IL-2 secreting Activated CD4+ T Cells | ↑ (comparable to other groups) | ↓ as compared to Asian (p < 0.05) in response to Omicron subvariants | ↑ as compared to Caucasian (p < 0.05) in response to Omicron subvariants |
| Anti-RBD IgG Levels | ↑ (comparable to other groups) | ↑ (comparable to other groups) | ↑ (comparable to other groups) |
| Neutralizing Antibody Titers | ↑ (comparable to other groups) | ↑ (comparable to other groups) | ↑ (comparable to other groups) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pantouli, F.; Silva-Moraes, V.; Ross, T.M. Ethnic Comparisons of Spike-Specific CD4+ T Cells, Serological Responses, and Neutralizing Antibody Titers Against SARS-CoV-2 Variants. Vaccines 2025, 13, 607. https://doi.org/10.3390/vaccines13060607
Pantouli F, Silva-Moraes V, Ross TM. Ethnic Comparisons of Spike-Specific CD4+ T Cells, Serological Responses, and Neutralizing Antibody Titers Against SARS-CoV-2 Variants. Vaccines. 2025; 13(6):607. https://doi.org/10.3390/vaccines13060607
Chicago/Turabian StylePantouli, Fani, Vanessa Silva-Moraes, and Ted M. Ross. 2025. "Ethnic Comparisons of Spike-Specific CD4+ T Cells, Serological Responses, and Neutralizing Antibody Titers Against SARS-CoV-2 Variants" Vaccines 13, no. 6: 607. https://doi.org/10.3390/vaccines13060607
APA StylePantouli, F., Silva-Moraes, V., & Ross, T. M. (2025). Ethnic Comparisons of Spike-Specific CD4+ T Cells, Serological Responses, and Neutralizing Antibody Titers Against SARS-CoV-2 Variants. Vaccines, 13(6), 607. https://doi.org/10.3390/vaccines13060607