
Destabilising Effect of Class B CpG Adjuvants on Different Proteins and Vaccine Candidates

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Recombinant Protein Expression and Purification
2.3. Thermal Stress Studies
2.4. SPR Binding Studies
2.5. NanoDSF Studies
2.6. Proteolysis Studies
2.7. Thermal Unfolding as a Function of pH
2.8. Thermal Unfolding as a Function of Salt Concentration
2.9. Thermal Reversibility
2.10. Statistical Analysis
3. Results
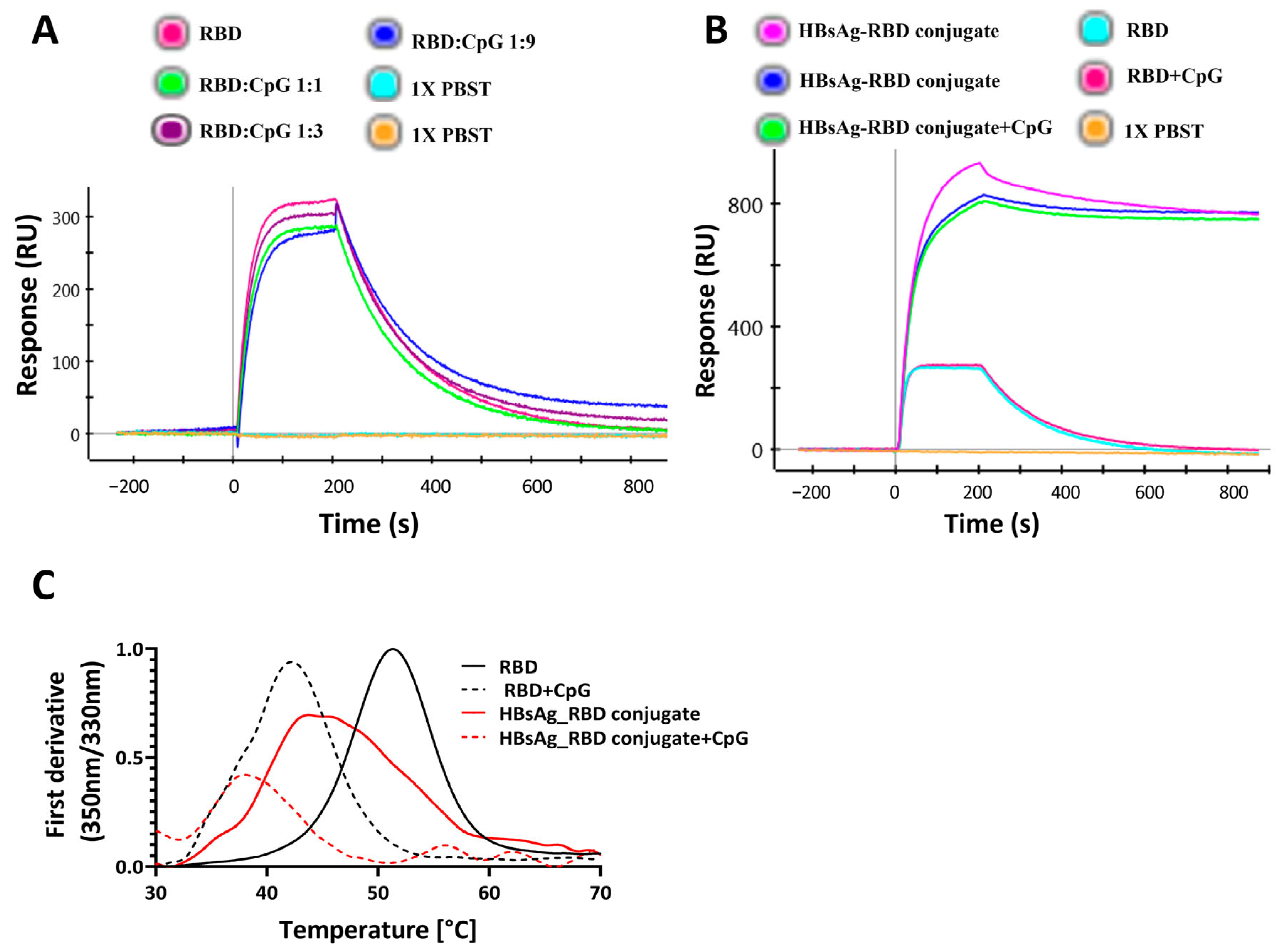
3.1. CpG Alters Protein Thermal Stability with No Observed Effect on Ligand Binding
3.2. CpG Increased Protein Sensitivity to Proteolytic Cleavage
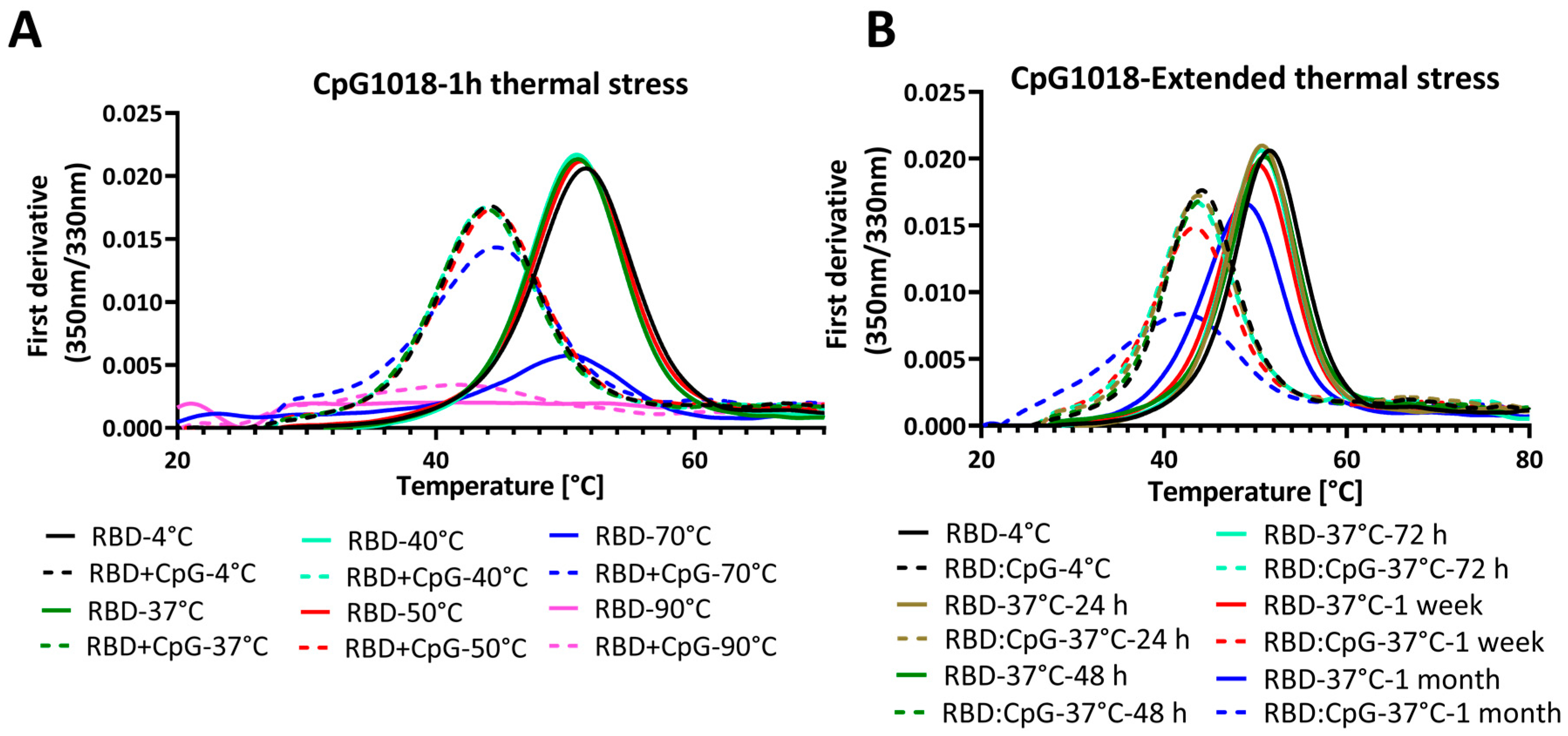
3.3. The Destabilising Effect of CpG on Protein: Ligand Binding Is Enhanced with Prolonged Incubation
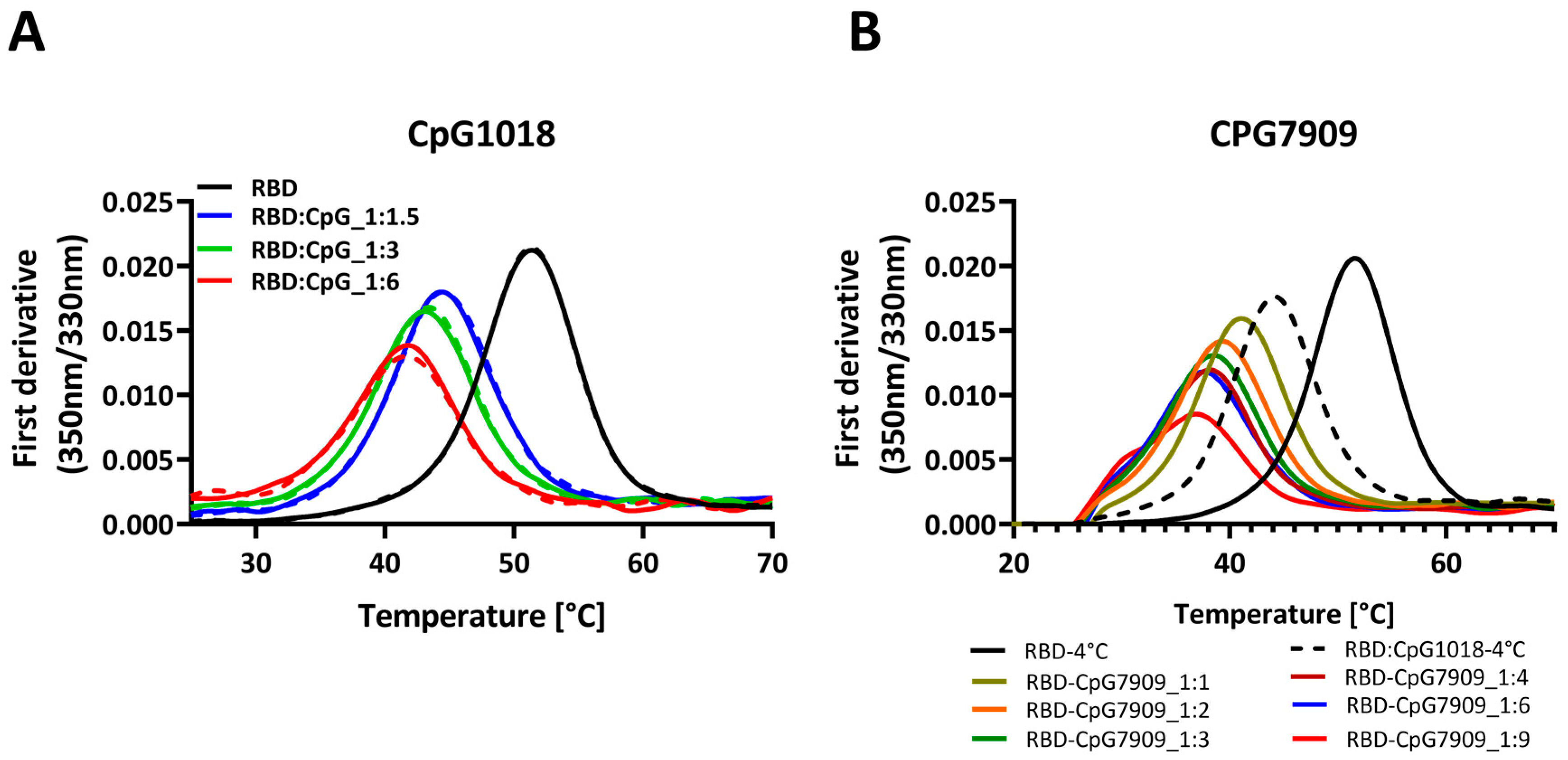
3.4. The Destabilising Effect of CpG Is Not Limited to RBD
3.5. The Effect of CpG on RBD Thermal Stability Is pH-Independent and Salt-Concentration-Dependent
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pulendran, B.; Arunachalam, P.S.; O’Hagan, D.T. Emerging Concepts in the Science of Vaccine Adjuvants. Nat. Rev. Drug Discov. 2021, 20, 454–475. [Google Scholar] [CrossRef]
- Bode, C.; Zhao, G.; Steinhagen, F.; Kinjo, T.; Klinman, D.M. CpG DNA as a Vaccine Adjuvant. Expert Rev. Vaccines 2011, 10, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Toussi, D.; Massari, P. Immune Adjuvant Effect of Molecularly-Defined Toll-Like Receptor Ligands. Vaccines 2014, 2, 323–353. [Google Scholar] [CrossRef]
- Krug, A.; Rothenfusser, S.; Hornung, V.; Jahrsdörfer, B.; Blackwell, S.; Ballas, Z.K.; Endres, S.; Krieg, A.M.; Hartmann, G. Identification of CpG Oligonucleotide Sequences with High Induction of IFN-α/β in Plasmacytoid Dendritic Cells. Eur. J. Immunol. 2001, 31, 2154–2163. [Google Scholar] [CrossRef] [PubMed]
- Kranzer, K.; Bauer, M.; Lipford, G.B.; Heeg, K.; Wagner, H.; Lang, R. CpG-Oligodeoxynucleotides Enhance T-Cell Receptor-Triggered Interferon-γ Production and up-Regulation of CD69 via Induction of Antigen-Presenting Cell-Derived Interferon Type I and Interleukin-12. Immunology 2000, 99, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.D. Development of the CpG Adjuvant 1018: A Case Study. Methods Mol. Biol. 2017, 1494, 15–27. [Google Scholar] [CrossRef]
- Cooper, C.L.; Davis, H.L.; Morris, M.L.; Efler, S.M.; Al Adhami, M.; Krieg, A.M.; Cameron, D.W.; Heathcote, J. CPG 7909, an Immunostimulatory TLR9 Agonist Oligodeoxynucleotide, as Adjuvant to Engerix-B® HBV Vaccine in Healthy Adults: A Double-Blind Phase I/II Study. J. Clin. Immunol. 2004, 24, 693–701. [Google Scholar] [CrossRef]
- Yang, J.; Li, B.; Yang, D.; Wu, J.; Yang, A.; Wang, W.; Lin, F.; Wan, X.; Li, Y.W.; Chen, Z.; et al. The Immunogenicity of Alum+CpG Adjuvant SARS-CoV-2 Inactivated Vaccine in Mice. Vaccine 2023, 41, 6064–6071. [Google Scholar] [CrossRef]
- Sen, G.; Chen, Q.; Snapper, C.M. Immunization of Aged Mice with a Pneumococcal Conjugate Vaccine Combined with an Unmethylated CpG-Containing Oligodeoxynucleotide Restores Defective Immunoglobulin G Antipolysaccharide Responses and Specific CD4 + -T-Cell Priming to Young Adult Levels. Infect. Immun. 2006, 74, 2177–2186. [Google Scholar] [CrossRef]
- Qin, W.; Jiang, J.; Chen, Q.; Yang, N.; Wang, Y.; Wei, X.; Ou, R. CpG ODN Enhances Immunization Effects of Hepatitis B Vaccine in Aged Mice. Cell. Mol. Immunol. 2004, 1, 148–152. [Google Scholar]
- Ming, F.; Yang, J.; Chu, P.; Ma, M.; Shi, J.; Cai, H.; Huang, C.; Li, H.; Jiang, Z.; Wang, H.; et al. Immunization of Aged Pigs with Attenuated Pseudorabies Virus Vaccine Combined with CpG Oligodeoxynucleotide Restores Defective Th1 Immune Responses. PLoS ONE 2013, 8, e65536. [Google Scholar] [CrossRef]
- Manning, B. CpG DNA Functions as an Effective Adjuvant for the Induction of Immune Responses in Aged Mice. Exp. Gerontol. 2001, 37, 107–126. [Google Scholar] [CrossRef] [PubMed]
- Maletto, B.A.; Rópolo, A.S.; Liscovsky, M.V.; Alignani, D.O.; Glocker, M.; Pistoresi-Palencia, M.C. CpG Oligodeoxinucleotides Functions as an Effective Adjuvant in Aged BALB/c Mice. Clin. Immunol. 2005, 117, 251–261. [Google Scholar] [CrossRef]
- Nanishi, E.; Borriello, F.; O’Meara, T.R.; McGrath, M.E.; Saito, Y.; Haupt, R.E.; Seo, H.-S.; van Haren, S.D.; Cavazzoni, C.B.; Brook, B.; et al. An Aluminum Hydroxide:CpG Adjuvant Enhances Protection Elicited by a SARS-CoV-2 Receptor Binding Domain Vaccine in Aged Mice. Sci. Transl. Med. 2022, 14, eabj5305. [Google Scholar] [CrossRef] [PubMed]
- Janssen, J.M.; Jackson, S.; Heyward, W.L.; Janssen, R.S. Immunogenicity of an Investigational Hepatitis B Vaccine with a Toll-like Receptor 9 Agonist Adjuvant (HBsAg-1018) Compared with a Licensed Hepatitis B Vaccine in Subpopulations of Healthy Adults 18-70 Years of Age. Vaccine 2015, 33, 3614–3618. [Google Scholar] [CrossRef] [PubMed]
- Sablan, B.P.; Kim, D.J.; Barzaga, N.G.; Chow, W.C.; Cho, M.; Ahn, S.H.; Hwang, S.G.; Lee, J.H.; Namini, H.; Heyward, W.L. Demonstration of Safety and Enhanced Seroprotection against Hepatitis B with Investigational HBsAg-1018 ISS Vaccine Compared to a Licensed Hepatitis B Vaccine. Vaccine 2012, 30, 2689–2696. [Google Scholar] [CrossRef]
- Richmond, P.; Hatchuel, L.; Dong, M.; Ma, B.; Hu, B.; Smolenov, I.; Li, P.; Liang, P.; Han, H.H.; Liang, J.; et al. Safety and Immunogenicity of S-Trimer (SCB-2019), a Protein Subunit Vaccine Candidate for COVID-19 in Healthy Adults: A Phase 1, Randomised, Double-Blind, Placebo-Controlled Trial. Lancet 2021, 397, 682–694. [Google Scholar] [CrossRef]
- Lee, G.H.; Lim, S.G. CpG-Adjuvanted Hepatitis B Vaccine (HEPLISAV-B®) Update. Expert Rev. Vaccines 2021, 20, 487–495. [Google Scholar] [CrossRef]
- Thuluva, S.; Paradkar, V.; Gunneri, S.; Yerroju, V.; Mogulla, R.; Suneetha, P.V.; Turaga, K.; Kyasani, M.; Manoharan, S.K.; Adabala, S.; et al. Immunogenicity and Safety of Biological E’s CORBEVAXTM Vaccine Compared to COVISHIELDTM (ChAdOx1 NCoV-19) Vaccine Studied in a Phase-3, Single Blind, Multicentre, Randomized Clinical Trial. Hum. Vaccines Immunother. 2023, 19, 2203632. [Google Scholar] [CrossRef]
- Thuluva, S.; Paradkar, V.; Gunneri, S.R.; Yerroju, V.; Mogulla, R.R.; Suneetha, P.V.; Turaga, K.; Kyasani, M.; Manoharan, S.K.; Adabala, S.; et al. Safety, Tolerability and Immunogenicity of Biological E’s CORBEVAXTM Vaccine in Children and Adolescents: A Prospective, Randomised, Double-Blind, Placebo Controlled, Phase-2/3 Study. Vaccine 2022, 40, 7130–7140. [Google Scholar] [CrossRef]
- Krug, A.; Towarowski, A.; Britsch, S.; Rothenfusser, S.; Hornung, V.; Bals, R.; Giese, T.; Engelmann, H.; Endres, S.; Krieg, A.M.; et al. Toll-like Receptor Expression Reveals CpG DNA as a Unique Microbial Stimulus for Plasmacytoid Dendritic Cells Which Synergizes with CD40 Ligand to Induce High Amounts of IL-12. Eur. J. Immunol. 2001, 31, 3026–3037. [Google Scholar] [CrossRef] [PubMed]
- Verthelyi, D.; Ishii, K.J.; Gursel, M.; Takeshita, F.; Klinman, D.M. Human Peripheral Blood Cells Differentially Recognize and Respond to Two Distinct CPG Motifs. J. Immunol. 2001, 166, 2372–2377. [Google Scholar] [CrossRef] [PubMed]
- Hanagata, N. Structure-Dependent Immunostimulatory Effect of CpG Oligodeoxynucleotides and Their Delivery System. Int. J. Nanomed. 2012, 7, 2181–2195. [Google Scholar] [CrossRef]
- Hartmann, G.; Battiany, J.; Poeck, H.; Wagner, M.; Kerkmann, M.; Lubenow, N.; Rothenfusser, S.; Endres, S. Rational Design of New CpG Oligonucleotides That Combine B Cell Activation with High IFN-Alpha Induction in Plasmacytoid Dendritic Cells. Eur. J. Immunol. 2003, 33, 1633–1641. [Google Scholar] [CrossRef] [PubMed]
- Marshall, J.D.; Fearon, K.; Abbate, C.; Subramanian, S.; Yee, P.; Gregorio, J.; Coffman, R.L.; Van Nest, G. Identification of a Novel CpG DNA Class and Motif That Optimally Stimulate B Cell and Plasmacytoid Dendritic Cell Functions. J. Leukoc. Biol. 2003, 73, 781–792. [Google Scholar] [CrossRef]
- Vollmer, J.; Weeratna, R.; Payette, P.; Jurk, M.; Schetter, C.; Laucht, M.; Wader, T.; Tluk, S.; Liu, M.; Davis, H.L.; et al. Characterization of Three CpG Oligodeoxynucleotide Classes with Distinct Immunostimulatory Activities. Eur. J. Immunol. 2004, 34, 251–262. [Google Scholar] [CrossRef]
- Vollmer, J.; Krieg, A.M. Immunotherapeutic Applications of CpG Oligodeoxynucleotide TLR9 Agonists. Adv. Drug Deliv. Rev. 2009, 61, 195–204. [Google Scholar] [CrossRef]
- Samulowitz, U.; Weber, M.; Weeratna, R.; Uhlmann, E.; Noll, B.; Krieg, A.M.; Vollmer, J. A Novel Class of Immune-Stimulatory CpG Oligodeoxynucleotides Unifies High Potency in Type I Interferon Induction with Preferred Structural Properties. Oligonucleotides 2010, 20, 93–101. [Google Scholar] [CrossRef]
- Kuo, T.-Y.; Lin, M.-Y.; Coffman, R.L.; Campbell, J.D.; Traquina, P.; Lin, Y.-J.; Liu, L.T.-C.; Cheng, J.; Wu, Y.-C.; Wu, C.-C.; et al. Development of CpG-Adjuvanted Stable Prefusion SARS-CoV-2 Spike Antigen as a Subunit Vaccine against COVID-19. Sci. Rep. 2020, 10, 20085. [Google Scholar] [CrossRef]
- Li, Y.; Chen, X. CpG 1018 Is an Effective Adjuvant for Influenza Nucleoprotein. Vaccines 2023, 11, 649. [Google Scholar] [CrossRef]
- Hyer, R.N.; Janssen, R.S. Immunogenicity and Safety of a 2-Dose Hepatitis B Vaccine, HBsAg/CpG 1018, in Persons with Diabetes Mellitus Aged 60–70 Years. Vaccine 2019, 37, 5854–5861. [Google Scholar] [CrossRef]
- Schillie, S.; Harris, A.; Link-Gelles, R.; Romero, J.; Ward, J.; Nelson, N. Recommendations of the Advisory Committee on Immunization Practices for Use of a Hepatitis B Vaccine with a Novel Adjuvant. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 455–458. [Google Scholar] [CrossRef] [PubMed]
- Barry, M.; Cooper, C. Review of Hepatitis B Surface Antigen-1018 ISS Adjuvant-Containing Vaccine Safety and Efficacy. Expert Opin. Biol. Ther. 2007, 7, 1731–1737. [Google Scholar] [CrossRef] [PubMed]
- Nagar, G.; Jain, S.; Rajurkar, M.; Lothe, R.; Rao, H.; Majumdar, S.; Gautam, M.; Rodriguez-Aponte, S.A.; Crowell, L.E.; Love, J.C.; et al. Large-Scale Purification and Characterization of Recombinant Receptor-Binding Domain (RBD) of SARS-CoV-2 Spike Protein Expressed in Yeast. Vaccines 2023, 11, 1602. [Google Scholar] [CrossRef] [PubMed]
- Shirbaghaee, Z.; Bolhassani, A. Different Applications of Virus-like Particles in Biology and Medicine: Vaccination and Delivery Systems. Biopolymers 2016, 105, 113–132. [Google Scholar] [CrossRef]
- Zürcher, P.; Sokolov, M.; Brühlmann, D.; Ducommun, R.; Stettler, M.; Souquet, J.; Jordan, M.; Broly, H.; Morbidelli, M.; Butté, A. Cell Culture Process Metabolomics Together with Multivariate Data Analysis Tools Opens New Routes for Bioprocess Development and Glycosylation Prediction. Biotechnol. Prog. 2020, 36, e3012. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, H.; Liu, M.; Zhang, T.; Zhang, J.; Wang, X.; Xiang, W. Rational Development of a Serum-Free Medium and Fed-Batch Process for a GS-CHO Cell Line Expressing Recombinant Antibody. Cytotechnology 2013, 65, 363–378. [Google Scholar] [CrossRef]
- Brune, K.D.; Leneghan, D.B.; Brian, I.J.; Ishizuka, A.S.; Bachmann, M.F.; Draper, S.J.; Biswas, S.; Howarth, M. Plug-and-Display: Decoration of Virus-Like Particles via Isopeptide Bonds for Modular Immunization. Sci. Rep. 2016, 6, 19234. [Google Scholar] [CrossRef]
- Marini, A.; Zhou, Y.; Li, Y.; Taylor, I.J.; Leneghan, D.B.; Jin, J.; Zaric, M.; Mekhaiel, D.; Long, C.A.; Miura, K.; et al. A Universal Plug-and-Display Vaccine Carrier Based on HBsAg VLP to Maximize Effective Antibody Response. Front. Immunol. 2019, 10, 2931. [Google Scholar] [CrossRef]
- Mittal, N.; Kumar, S.; Rajmani, R.S.; Singh, R.; Lemoine, C.; Jakob, V.; Bj, S.; Jagannath, N.; Bhat, M.; Chakraborty, D.; et al. Enhanced Protective Efficacy of a Thermostable RBD-S2 Vaccine Formulation against SARS-CoV-2 and Its Variants. npj Vaccines 2023, 8, 161. [Google Scholar] [CrossRef]
- Kanjo, K.; Chattopadhyay, G.; Malladi, S.K.; Singh, R.; Jayatheertha, S.; Varadarajan, R. Biophysical Correlates of Enhanced Immunogenicity of a Stabilized Variant of the Receptor Binding Domain of SARS-CoV-2. J. Phys. Chem. B 2023, 127, 1704–1714. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, G.; Varadarajan, R. Facile Measurement of Protein Stability and Folding Kinetics Using a Nano Differential Scanning Fluorimeter. Protein Sci. 2019, 28, 1127–1134. [Google Scholar] [CrossRef]
- Chiba, S.; Kong, H.; Neumann, G.; Kawaoka, Y. Influenza H3 Hemagglutinin Vaccine with Scrambled Immunodominant Epitopes Elicits Antibodies Directed toward Immunosubdominant Head Epitopes. MBio 2023, 14, e0062223. [Google Scholar] [CrossRef]
- Lenart, K.; Arcoverde Cerveira, R.; Hellgren, F.; Ols, S.; Sheward, D.J.; Kim, C.; Cagigi, A.; Gagne, M.; Davis, B.; Germosen, D.; et al. Three Immunizations with Novavax’s Protein Vaccines Increase Antibody Breadth and Provide Durable Protection from SARS-CoV-2. npj Vaccines 2024, 9, 17. [Google Scholar] [CrossRef]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the MRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Malladi, S.K.; Singh, R.; Pandey, S.; Gayathri, S.; Kanjo, K.; Ahmed, S.; Khan, M.S.; Kalita, P.; Girish, N.; Upadhyaya, A.; et al. Design of a Highly Thermotolerant, Immunogenic SARS-CoV-2 Spike Fragment. J. Biol. Chem. 2021, 296, 100025. [Google Scholar] [CrossRef]
- Reed, S.G.; Orr, M.T.; Fox, C.B. Key Roles of Adjuvants in Modern Vaccines. Nat. Med. 2013, 19, 1597–1608. [Google Scholar] [CrossRef]
- Klaschik, S.; Tross, D.; Klinman, D.M. Inductive and Suppressive Networks Regulate TLR9-Dependent Gene Expression in Vivo. J. Leukoc. Biol. 2009, 85, 788–795. [Google Scholar] [CrossRef]
- Kobayashi, H.; Horner, A.A.; Takabayashi, K.; Nguyen, M.-D.; Huang, E.; Cinman, N.; Raz, E. Immunostimulatory DNA Prepriming: A Novel Approach for Prolonged Th1-Biased Immunity. Cell. Immunol. 1999, 198, 69–75. [Google Scholar] [CrossRef]
- Ozorowski, G.; Cupo, A.; Golabek, M.; LoPiccolo, M.; Ketas, T.A.; Cavallary, M.; Cottrell, C.A.; Klasse, P.J.; Ward, A.B.; Moore, J.P. Effects of Adjuvants on HIV-1 Envelope Glycoprotein SOSIP Trimers In Vitro. J. Virol. 2018, 92, 381–399. [Google Scholar] [CrossRef]
- Bajoria, S.; Kaur, K.; Kumru, O.S.; Van Slyke, G.; Doering, J.; Novak, H.; Rodriguez Aponte, S.A.; Dalvie, N.C.; Naranjo, C.A.; Johnston, R.S.; et al. Antigen-Adjuvant Interactions, Stability, and Immunogenicity Profiles of a SARS-CoV-2 Receptor-Binding Domain (RBD) Antigen Formulated with Aluminum Salt and CpG Adjuvants. Hum. Vaccines Immunother. 2022, 18, 2079346. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kanjo, K.; Lothe, R.; Nagar, G.; Rajurkar, M.; Rao, H.; Batwal, S.; Shaligram, U.; Varadarajan, R. Destabilising Effect of Class B CpG Adjuvants on Different Proteins and Vaccine Candidates. Vaccines 2025, 13, 395. https://doi.org/10.3390/vaccines13040395
Kanjo K, Lothe R, Nagar G, Rajurkar M, Rao H, Batwal S, Shaligram U, Varadarajan R. Destabilising Effect of Class B CpG Adjuvants on Different Proteins and Vaccine Candidates. Vaccines. 2025; 13(4):395. https://doi.org/10.3390/vaccines13040395
Chicago/Turabian StyleKanjo, Kawkab, Rakesh Lothe, Gaurav Nagar, Meghraj Rajurkar, Harish Rao, Saurabh Batwal, Umesh Shaligram, and Raghavan Varadarajan. 2025. "Destabilising Effect of Class B CpG Adjuvants on Different Proteins and Vaccine Candidates" Vaccines 13, no. 4: 395. https://doi.org/10.3390/vaccines13040395
APA StyleKanjo, K., Lothe, R., Nagar, G., Rajurkar, M., Rao, H., Batwal, S., Shaligram, U., & Varadarajan, R. (2025). Destabilising Effect of Class B CpG Adjuvants on Different Proteins and Vaccine Candidates. Vaccines, 13(4), 395. https://doi.org/10.3390/vaccines13040395