
Evaluation of Long-Term Adaptive Immune Responses Specific to SARS-CoV-2: Effect of Various Vaccination and Omicron Exposure



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
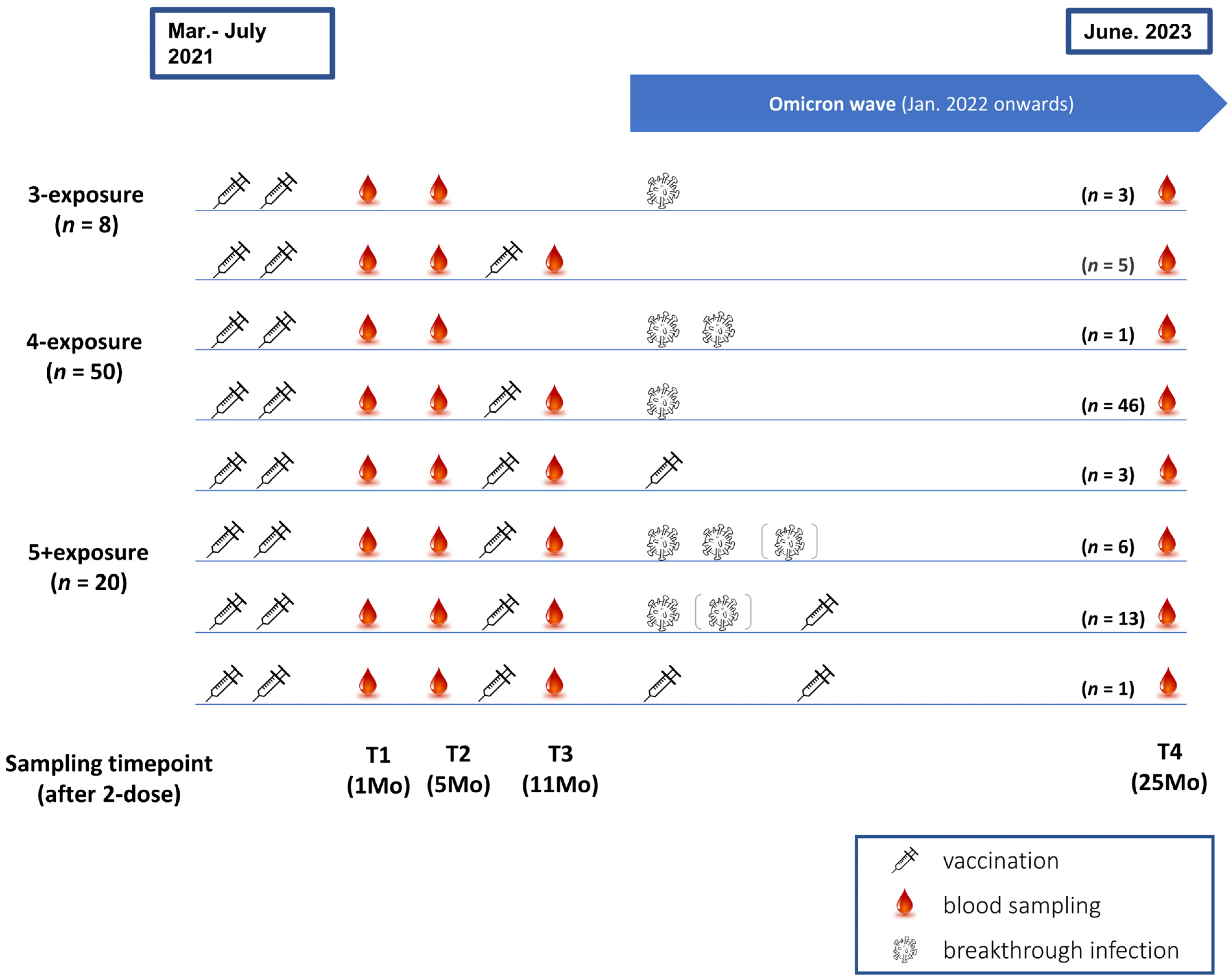
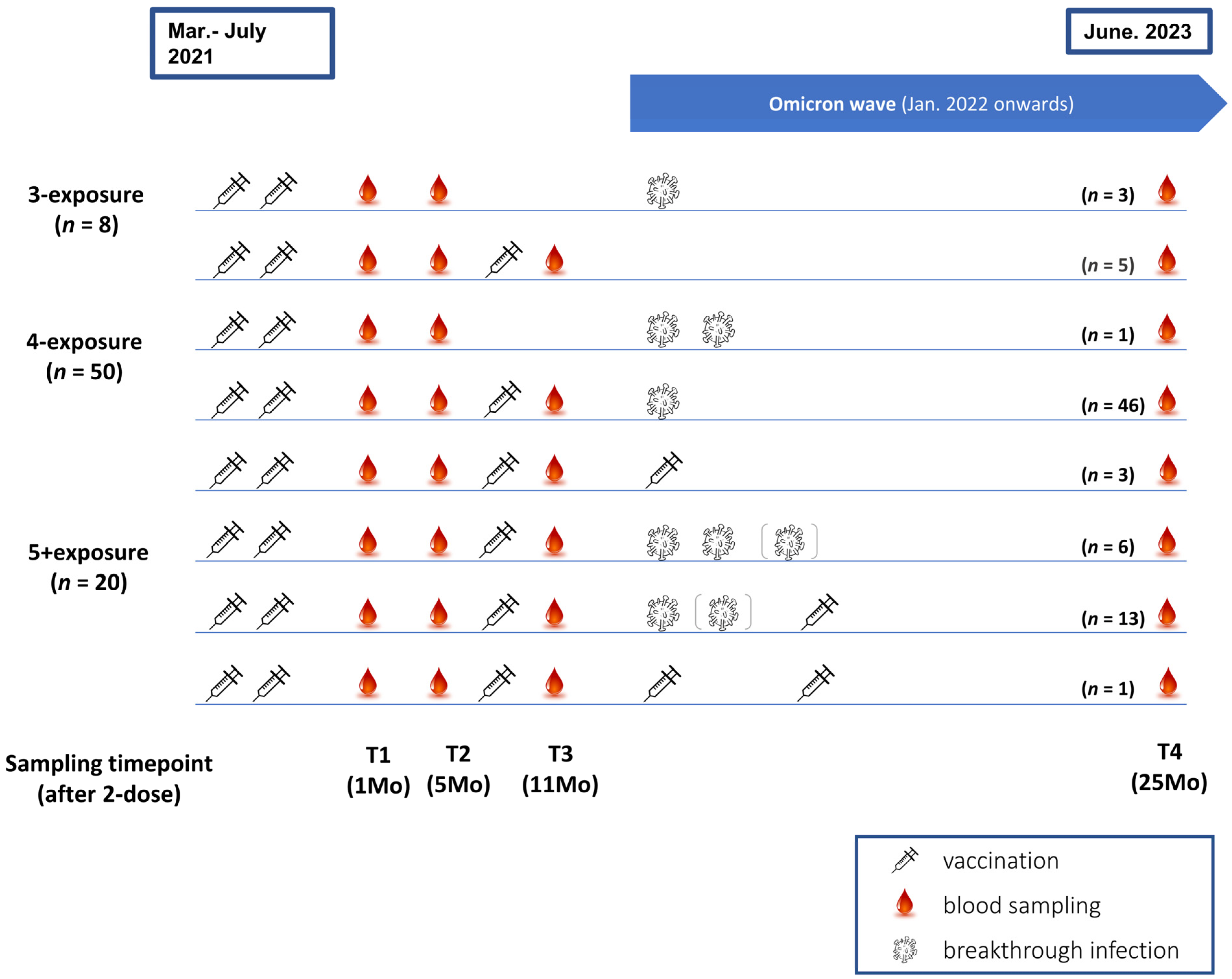
2.1. Enrolment and Sample Collection
2.2. SARS-CoV-2–Specific Humoral Immune Response Assays
2.3. SARS-CoV-2–Specific Cellular Immune Response Assays
2.4. Statistical Analysis
3. Results
3.1. Participant Characteristics
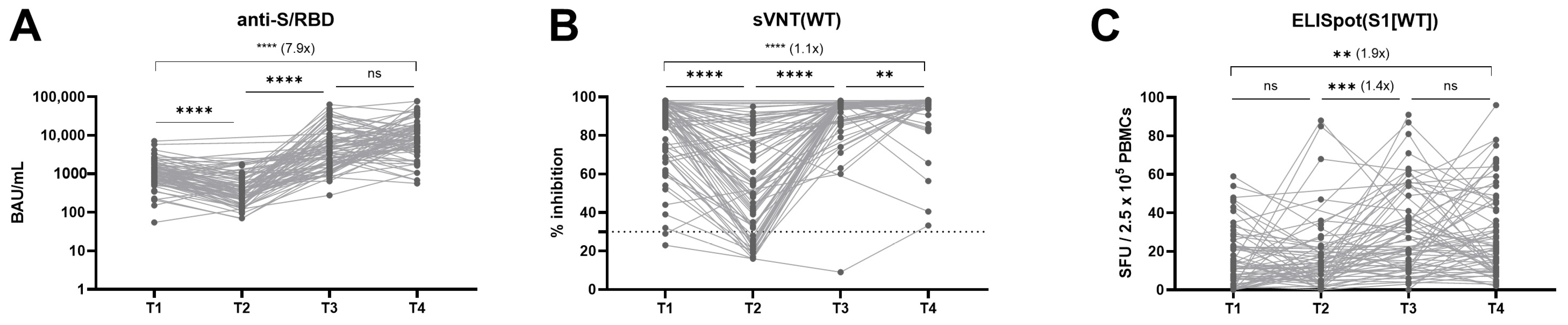
3.2. Longitudinal Kinetics of Humoral and Cellular Immune Response
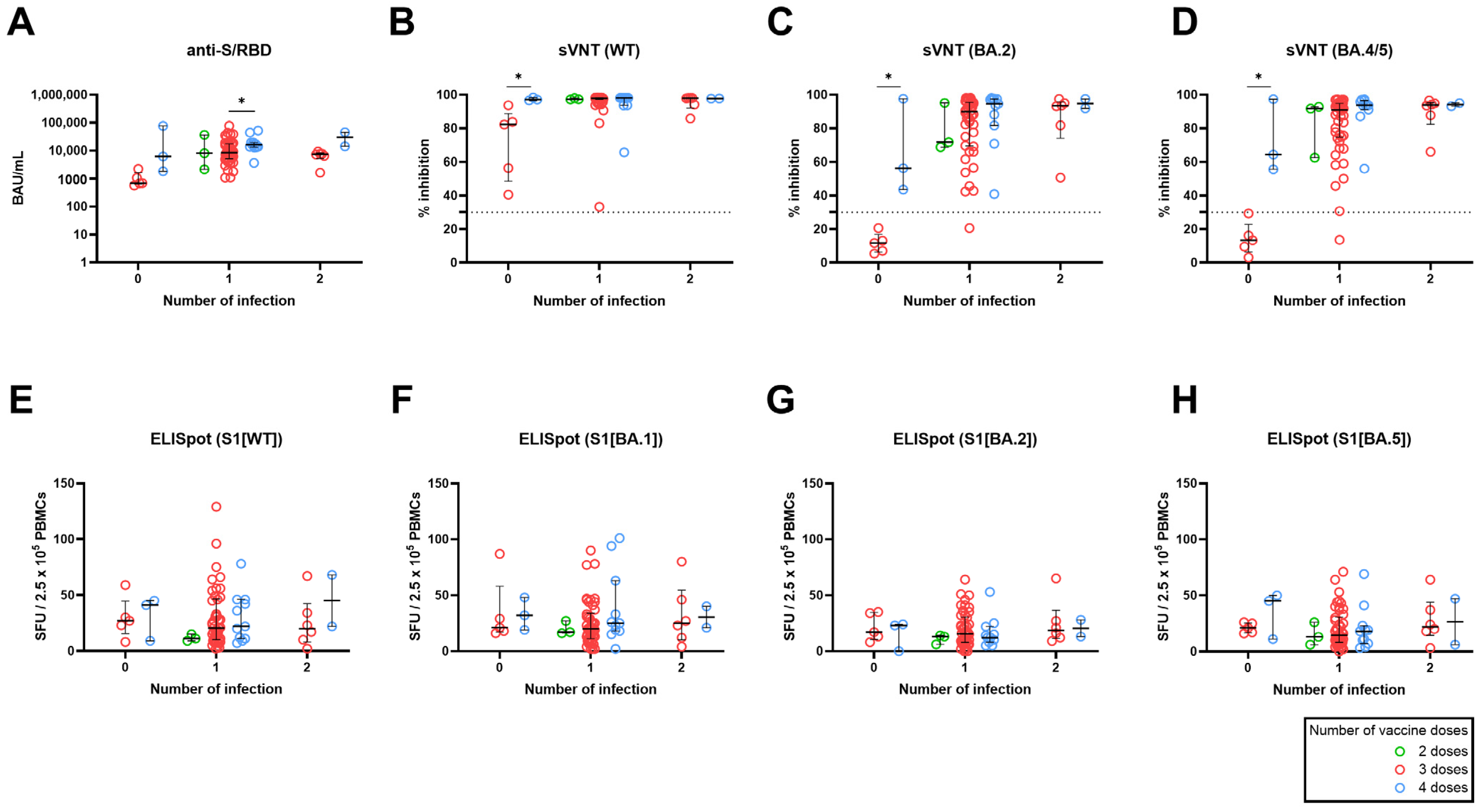
3.3. Effect of the Number of Vaccine Doses Stratified by Number of Breakthrough Infection on SARS-CoV-2–Specific Adaptive Immune Response
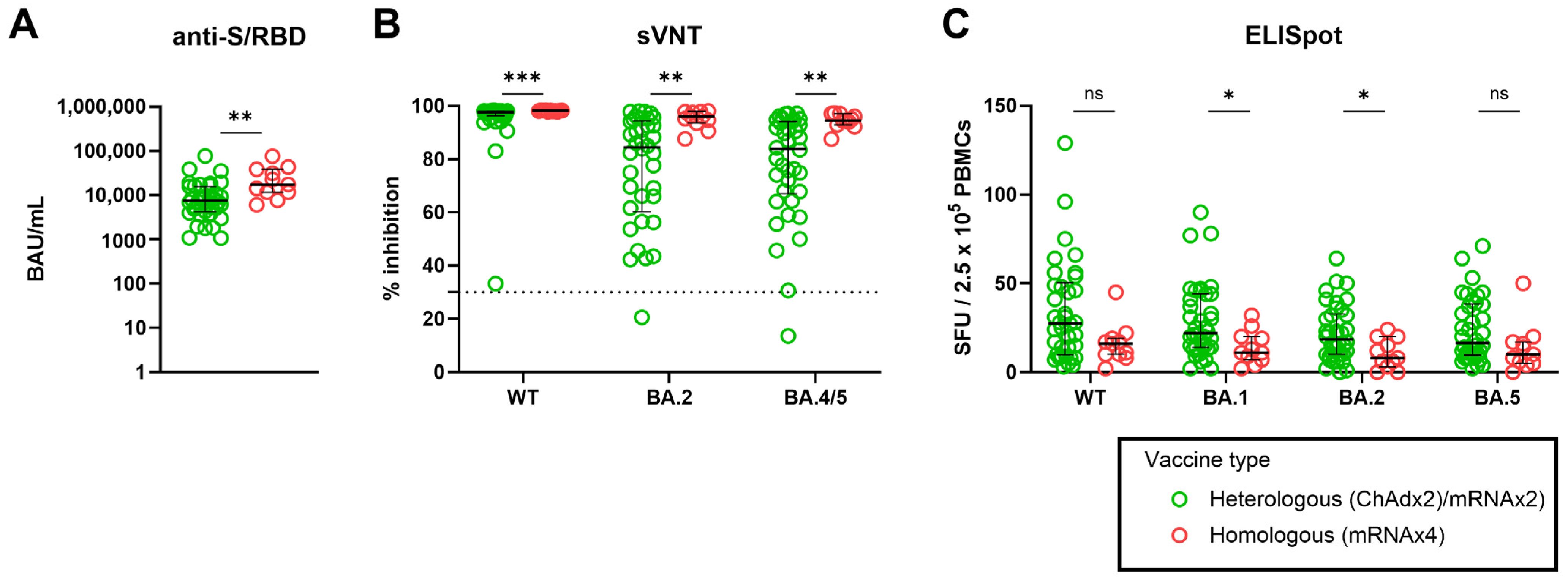
3.4. Effect of Heterologous versus Homologous Vaccination on SARS-CoV-2–Specific Adaptive Immune Response
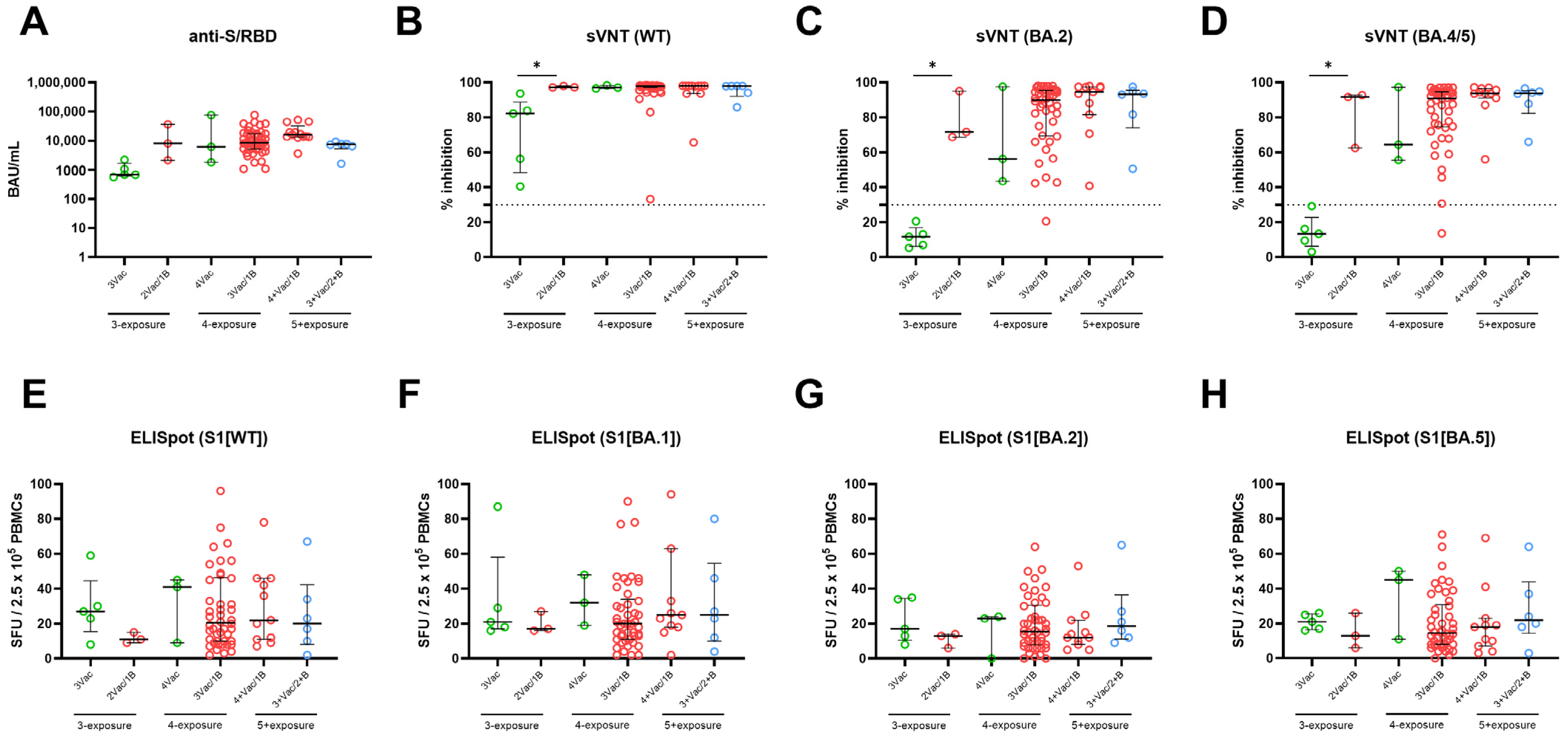
3.5. Effect of Breakthrough Infection and Bivalent Vaccination Stratified by Number of Antigen-Exposure on SARS-CoV-2–Specific Adaptive Immune Response
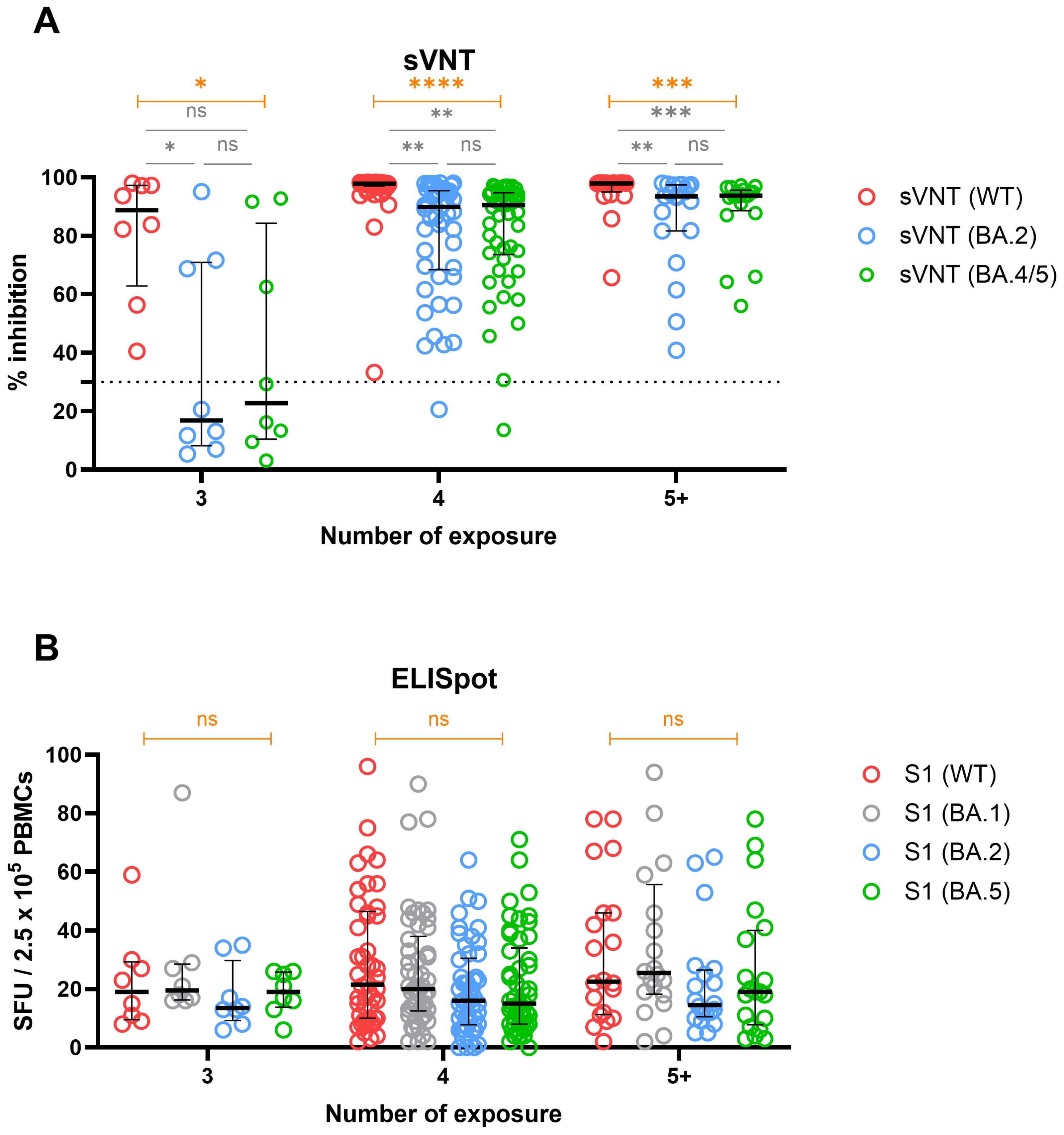
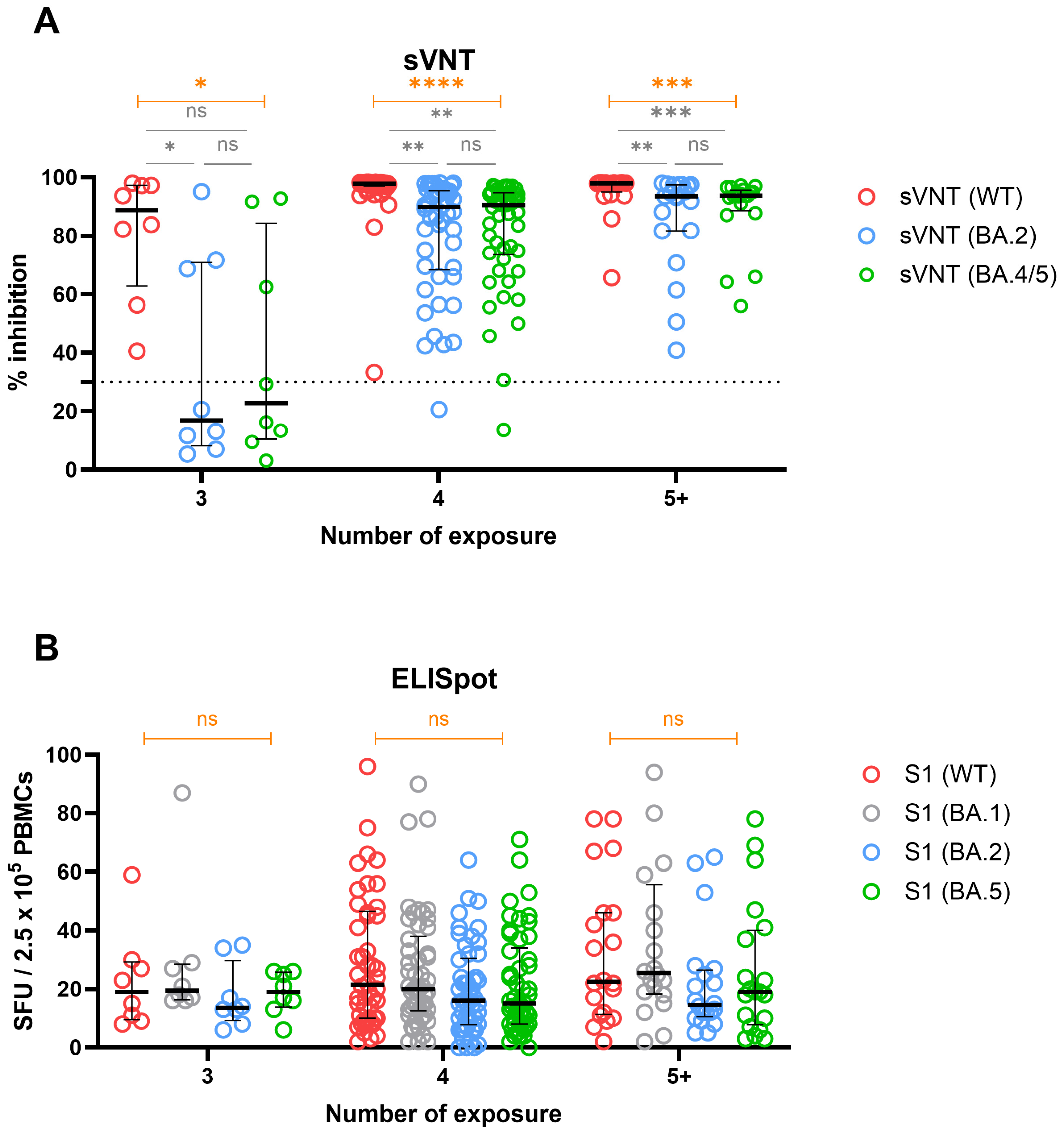
3.6. Cross-Immunity against WT and Omicron Variants in Groups Stratified by Number of Exposures
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ssentongo, P.; Ssentongo, A.E.; Voleti, N.; Groff, D.; Sun, A.; Ba, D.M.; Nunez, J.; Parent, L.J.; Chinchilli, V.M.; Paules, C.I. SARS-CoV-2 vaccine effectiveness against infection, symptomatic and severe COVID-19: A systematic review and meta-analysis. BMC Infect. Dis. 2022, 22, 439. [Google Scholar] [CrossRef]
- The, L. The COVID-19 pandemic in 2023: Far from over. Lancet 2023, 401, 79. [Google Scholar] [CrossRef]
- Bergwerk, M.; Gonen, T.; Lustig, Y.; Amit, S.; Lipsitch, M.; Cohen, C.; Mandelboim, M.; Levin, E.G.; Rubin, C.; Indenbaum, V.; et al. COVID-19 Breakthrough Infections in Vaccinated Health Care Workers. N. Engl. J. Med. 2021, 385, 1474–1484. [Google Scholar] [CrossRef]
- Wang, S.Y.; Juthani, P.V.; Borges, K.A.; Shallow, M.K.; Gupta, A.; Price, C.; Won, C.H.; Chun, H.J. Severe breakthrough COVID-19 cases in the SARS-CoV-2 delta (B.1.617.2) variant era. Lancet Microbe 2022, 3, e4–e5. [Google Scholar] [CrossRef]
- Pusnik, J.; Monzon-Posadas, W.O.; Zorn, J.; Peters, K.; Baum, M.; Proksch, H.; Schluter, C.B.; Alter, G.; Menting, T.; Streeck, H. SARS-CoV-2 humoral and cellular immunity following different combinations of vaccination and breakthrough infection. Nat. Commun. 2023, 14, 572. [Google Scholar] [CrossRef]
- Lyke, K.E.; Atmar, R.L.; Islas, C.D.; Posavad, C.M.; Szydlo, D.; Paul Chourdhury, R.; Deming, M.E.; Eaton, A.; Jackson, L.A.; Branche, A.R.; et al. Rapid decline in vaccine-boosted neutralizing antibodies against SARS-CoV-2 Omicron variant. Cell Rep. Med. 2022, 3, 100679. [Google Scholar] [CrossRef]
- Kotaki, R.; Moriyama, S.; Takahashi, Y. Humoral immunity for durable control of SARS-CoV-2 and its variants. Inflamm. Regen. 2023, 43, 4. [Google Scholar] [CrossRef]
- Luczkowiak, J.; Rivas, G.; Labiod, N.; Lasala, F.; Rolo, M.; Lora-Tamayo, J.; Mancheno-Losa, M.; Rial-Crestelo, D.; Perez-Rivilla, A.; Folgueira, M.D.; et al. Cross neutralization of SARS-CoV-2 omicron subvariants after repeated doses of COVID-19 mRNA vaccines. J. Med. Virol. 2023, 95, e28268. [Google Scholar] [CrossRef]
- Chen, Y.; Tong, P.; Whiteman, N.; Sanjari Moghaddam, A.; Zarghami, M.; Zuiani, A.; Habibi, S.; Gautam, A.; Keerti; Bi, C.; et al. Immune recall improves antibody durability and breadth to SARS-CoV-2 variants. Sci. Immunol. 2022, 7, eabp8328. [Google Scholar] [CrossRef]
- Fong, C.H.; Zhang, X.; Chen, L.L.; Poon, R.W.; Chan, B.P.; Zhao, Y.; Wong, C.K.; Chan, K.H.; Yuen, K.Y.; Hung, I.F.; et al. Effect of vaccine booster, vaccine type, and hybrid immunity on humoral and cellular immunity against SARS-CoV-2 ancestral strain and Omicron variant sublineages BA.2 and BA.5 among older adults with comorbidities: A cross sectional study. EBioMedicine 2023, 88, 104446. [Google Scholar] [CrossRef]
- Guo, L.; Wang, G.; Wang, Y.; Zhang, Q.; Ren, L.; Gu, X.; Huang, T.; Zhong, J.; Wang, Y.; Wang, X.; et al. SARS-CoV-2-specific antibody and T-cell responses 1 year after infection in people recovered from COVID-19: A longitudinal cohort study. Lancet Microbe 2022, 3, e348–e356. [Google Scholar] [CrossRef]
- Zuo, J.; Dowell, A.C.; Pearce, H.; Verma, K.; Long, H.M.; Begum, J.; Aiano, F.; Amin-Chowdhury, Z.; Hoschler, K.; Brooks, T.; et al. Robust SARS-CoV-2-specific T cell immunity is maintained at 6 months following primary infection. Nat. Immunol. 2021, 22, 620–626. [Google Scholar] [CrossRef]
- Chivu-Economescu, M.; Bleotu, C.; Grancea, C.; Chiriac, D.; Botezatu, A.; Iancu, I.V.; Pitica, I.; Necula, L.G.; Neagu, A.; Matei, L.; et al. Kinetics and persistence of cellular and humoral immune responses to SARS-CoV-2 vaccine in healthcare workers with or without prior COVID-19. J. Cell Mol. Med. 2022, 26, 1293–1305. [Google Scholar] [CrossRef]
- Herzberg, J.; Fischer, B.; Lindenkamp, C.; Becher, H.; Becker, A.K.; Honarpisheh, H.; Guraya, S.Y.; Strate, T.; Knabbe, C. Persistence of Immune Response in Health Care Workers After Two Doses BNT162b2 in a Longitudinal Observational Study. Front. Immunol. 2022, 13, 839922. [Google Scholar] [CrossRef]
- Tao, Y.; Ma, M.; Hu, F.; Li, M.; Geng, Y.; Wan, Y.; Mao, M.; Chen, L.; Shen, Y.; Zhu, L.; et al. A longitudinal study of humoral immune responses induced by a 3-dose inactivated COVID-19 vaccine in an observational, prospective cohort. BMC Immunol. 2022, 23, 57. [Google Scholar] [CrossRef]
- Monzon-Posadas, W.O.; Zorn, J.; Peters, K.; Baum, M.; Proksch, H.; Schluter, C.B.; Menting, T.; Pusnik, J.; Streeck, H. Longitudinal monitoring of mRNA-vaccine-induced immunity against SARS-CoV-2. Front. Immunol. 2023, 14, 1066123. [Google Scholar] [CrossRef]
- Hvidt, A.K.; Guo, H.; Andersen, R.; Lende, S.S.F.; Vibholm, L.K.; Sogaard, O.S.; Schleimann, M.H.; Russell, V.; Cheung, A.M.; Paramithiotis, E.; et al. Long-term humoral and cellular immunity after primary SARS-CoV-2 infection: A 20-month longitudinal study. BMC Immunol. 2023, 24, 45. [Google Scholar] [CrossRef]
- Ko, G.Y.; Lee, J.; Bae, H.; Ryu, J.H.; Park, H.S.; Kang, H.; Jung, J.; Choi, A.R.; Lee, R.; Lee, D.G.; et al. Longitudinal Analysis of SARS-CoV-2-Specific Cellular and Humoral Immune Responses and Breakthrough Infection following BNT162b2/BNT162b2/BNT162b2 and ChAdOx1/ChAdOx1/BNT162b2 Vaccination: A Prospective Cohort in Naive Healthcare Workers. Vaccines 2023, 11, 1613. [Google Scholar] [CrossRef]
- Lee, D.W.; Kim, J.M.; Park, A.K.; Kim, D.W.; Kim, J.Y.; Lim, N.; Lee, H.; Kim, I.H.; Kim, J.A.; Lee, C.Y.; et al. Genomic epidemiology of SARS-CoV-2 Omicron variants in the Republic of Korea. Sci. Rep. 2022, 12, 22414. [Google Scholar] [CrossRef]
- Jeong, S.; Kim, J.S.; Lee, S.K.; Cho, E.J.; Hyun, J.; Song, W.; Kim, H.S. Tracking the Genomic Evolution of SARS-CoV-2 for 29 Months in South Korea. Viruses 2023, 15, 873. [Google Scholar] [CrossRef]
- Park, A.K.; Kim, I.H.; Lee, C.Y.; Kim, J.A.; Lee, H.; Kim, H.M.; Lee, N.J.; Woo, S.; Lee, J.; Rhee, J.; et al. Rapid Emergence of the Omicron Variant of Severe Acute Respiratory Syndrome Coronavirus 2 in Korea. Ann. Lab. Med. 2023, 43, 211–213. [Google Scholar] [CrossRef]
- Hwang, S.D.; Ha, J.; Yun, Y.N.; Jung, S.; Chun, J.-H. The Status of Genomic Surveillance of the Coronavirus Disease 2019 Virus Variants in Chungcheong Region in 2021–2022. Public Health Wkly. Rep. 2023, 16, 1257–1271. [Google Scholar]
- Qu, P.; Evans, J.P.; Faraone, J.N.; Zheng, Y.M.; Carlin, C.; Anghelina, M.; Stevens, P.; Fernandez, S.; Jones, D.; Lozanski, G.; et al. Enhanced neutralization resistance of SARS-CoV-2 Omicron subvariants BQ.1, BQ.1.1, BA.4.6, BF.7, and BA.2.75.2. Cell Host Microbe 2023, 31, 9–17.e3. [Google Scholar] [CrossRef]
- Lustig, Y.; Gonen, T.; Meltzer, L.; Gilboa, M.; Indenbaum, V.; Cohen, C.; Amit, S.; Jaber, H.; Doolman, R.; Asraf, K.; et al. Superior immunogenicity and effectiveness of the third compared to the second BNT162b2 vaccine dose. Nat. Immunol. 2022, 23, 940–946. [Google Scholar] [CrossRef]
- Regev-Yochay, G.; Gonen, T.; Gilboa, M.; Mandelboim, M.; Indenbaum, V.; Amit, S.; Meltzer, L.; Asraf, K.; Cohen, C.; Fluss, R.; et al. Efficacy of a Fourth Dose of COVID-19 mRNA Vaccine against Omicron. N. Engl. J. Med. 2022, 386, 1377–1380. [Google Scholar] [CrossRef]
- Koutsakos, M.; Ellebedy, A.H. Immunological imprinting: Understanding COVID-19. Immunity 2023, 56, 909–913. [Google Scholar] [CrossRef]
- Chemaitelly, H.; Ayoub, H.H.; Tang, P.; Hasan, M.R.; Coyle, P.; Yassine, H.M.; Al-Khatib, H.A.; Smatti, M.K.; Al-Kanaani, Z.; Al-Kuwari, E.; et al. Immune Imprinting and Protection against Repeat Reinfection with SARS-CoV-2. N. Engl. J. Med. 2022, 387, 1716–1718. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Behrens, G.M.N.; Arora, P.; Kempf, A.; Nehlmeier, I.; Cossmann, A.; Manthey, L.; Dopfer-Jablonka, A.; Pohlmann, S. Effect of hybrid immunity and bivalent booster vaccination on omicron sublineage neutralisation. Lancet Infect. Dis. 2023, 23, 25–28. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Kurhade, C.; Patel, S.; Kitchin, N.; Tompkins, K.; Cutler, M.; Cooper, D.; Yang, Q.; Cai, H.; Muik, A.; et al. Neutralization of BA.4-BA.5, BA.4.6, BA.2.75.2, BQ.1.1, and XBB.1 with Bivalent Vaccine. N. Engl. J. Med. 2023, 388, 854–857. [Google Scholar] [CrossRef] [PubMed]
- Chalkias, S.; Harper, C.; Vrbicky, K.; Walsh, S.R.; Essink, B.; Brosz, A.; McGhee, N.; Tomassini, J.E.; Chen, X.; Chang, Y.; et al. A Bivalent Omicron-Containing Booster Vaccine against COVID-19. N. Engl. J. Med. 2022, 387, 1279–1291. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chandrashekar, A.; Sellers, D.; Barrett, J.; Jacob-Dolan, C.; Lifton, M.; McMahan, K.; Sciacca, M.; VanWyk, H.; Wu, C.; et al. Vaccines elicit highly conserved cellular immunity to SARS-CoV-2 Omicron. Nature 2022, 603, 493–496. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, H.; Jung, J.; Ko, G.Y.; Lee, J.; Oh, E.-J. Evaluation of Long-Term Adaptive Immune Responses Specific to SARS-CoV-2: Effect of Various Vaccination and Omicron Exposure. Vaccines 2024, 12, 301. https://doi.org/10.3390/vaccines12030301
Kang H, Jung J, Ko GY, Lee J, Oh E-J. Evaluation of Long-Term Adaptive Immune Responses Specific to SARS-CoV-2: Effect of Various Vaccination and Omicron Exposure. Vaccines. 2024; 12(3):301. https://doi.org/10.3390/vaccines12030301
Chicago/Turabian StyleKang, Hyunhye, Jin Jung, Geon Young Ko, Jihyun Lee, and Eun-Jee Oh. 2024. "Evaluation of Long-Term Adaptive Immune Responses Specific to SARS-CoV-2: Effect of Various Vaccination and Omicron Exposure" Vaccines 12, no. 3: 301. https://doi.org/10.3390/vaccines12030301
APA StyleKang, H., Jung, J., Ko, G. Y., Lee, J., & Oh, E.-J. (2024). Evaluation of Long-Term Adaptive Immune Responses Specific to SARS-CoV-2: Effect of Various Vaccination and Omicron Exposure. Vaccines, 12(3), 301. https://doi.org/10.3390/vaccines12030301