
Rational Design and Characterization of Trispecific Antibodies Targeting the HIV-1 Receptor and Envelope Glycoprotein

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Expression of Recombinant Human CD4 and CCR5 Proteins
2.2. Construction, Expression, and Purification of IgG and Trispecific Antibodies
2.3. ELISA
2.4. Kd Determination
2.5. Virus Neutralization Assay
2.6. Humanized Mouse Model
2.7. HIV-1 RNA Load Measurement
2.8. In Vivo Assay in Humanized Mice
2.9. Antibody-Dependent Cell-Mediated Cytotoxicity (ADCC) Assay
3. Results
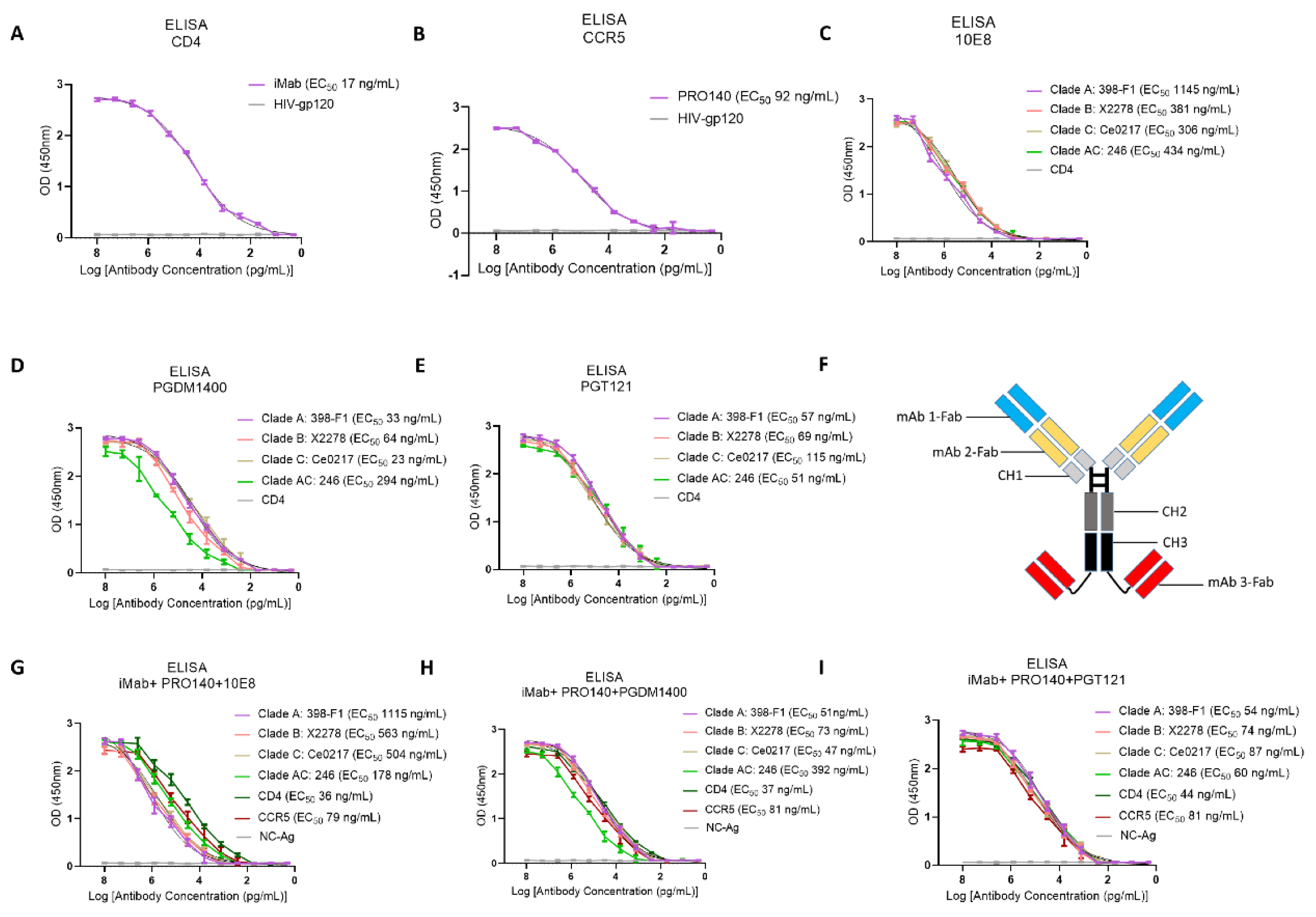
3.1. Determination of Antigen-Binding Activity of the Antibodies Generated in This Study
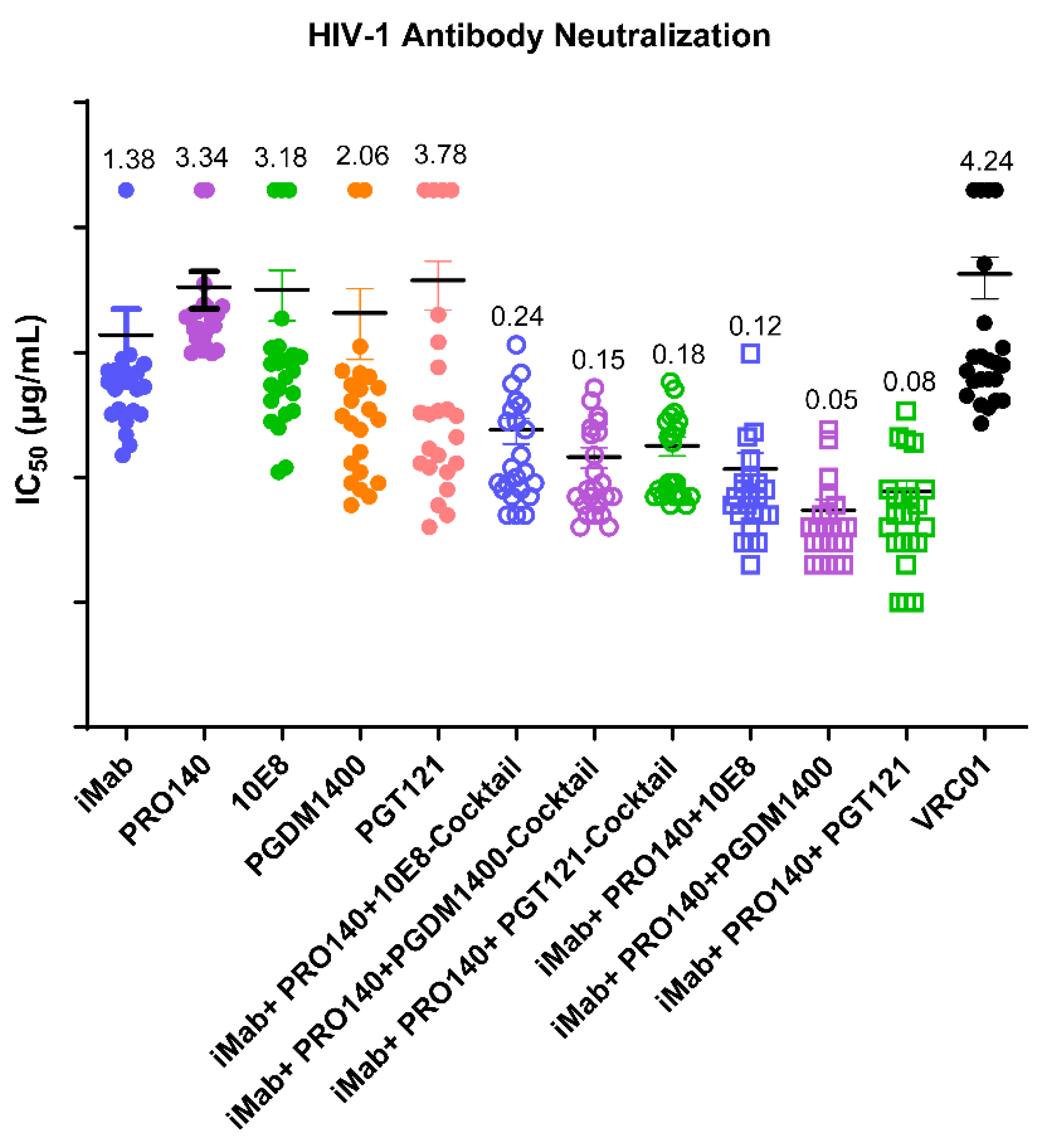
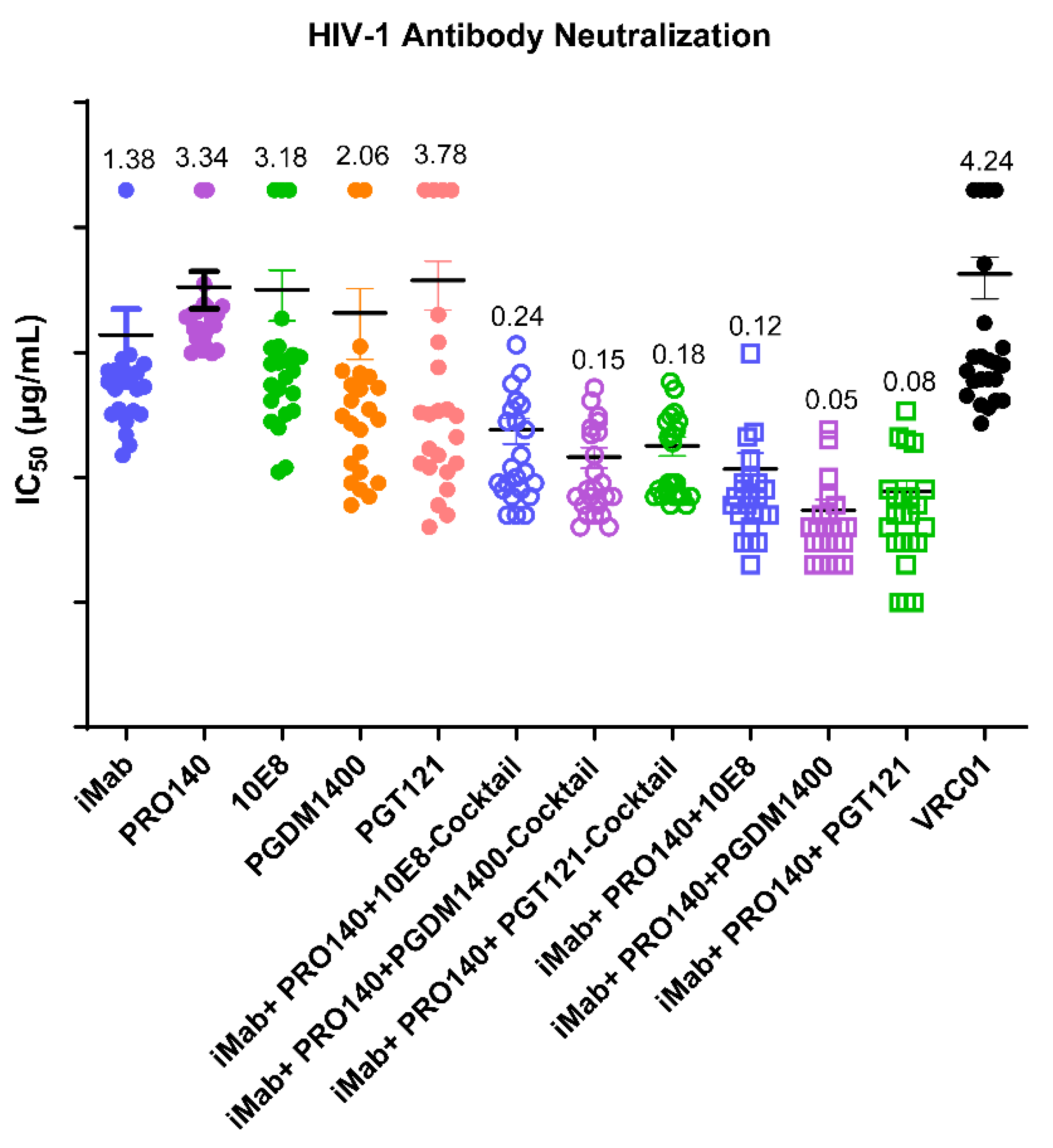
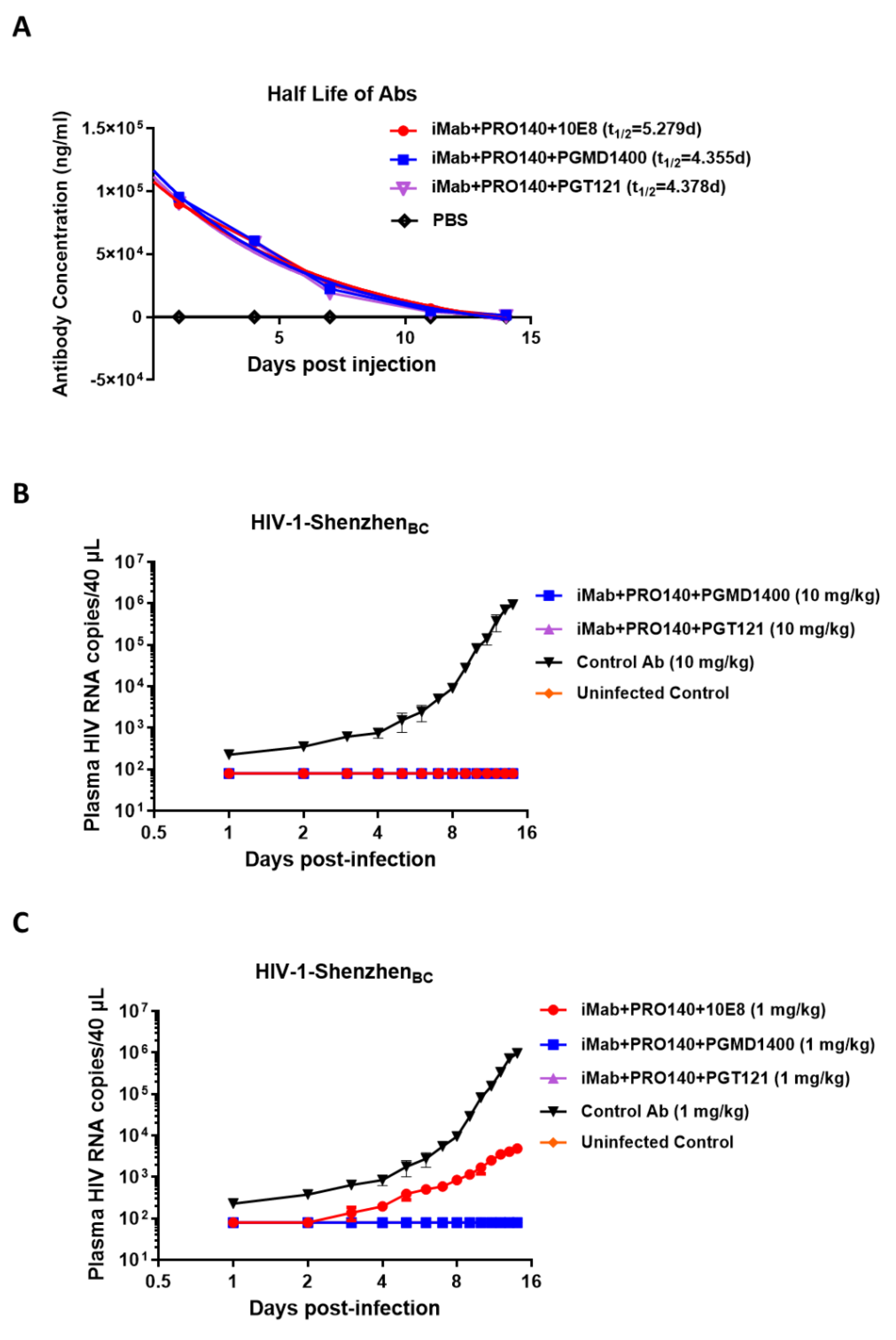
3.2. Determination of In Vitro Neutralization Activity against HIV-1 Pseudoviruses and In Vivo Antiviral Activity of the Antibodies Generated in This Study
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Segal-Maurer, S.; DeJesus, E.; Stellbrink, H.-J.; Castagna, A.; Richmond, G.J.; Sinclair, G.I.; Siripassorn, K.; Ruane, P.J.; Berhe, M.; Wang, H.; et al. Capsid Inhibition with Lenacapavir in Multidrug-Resistant HIV-1 Infection. N. Engl. J. Med. 2022, 386, 1793–1803. [Google Scholar] [CrossRef] [PubMed]
- Mayer, K.H.; Molina, J.-M.; Thompson, M.A.; Anderson, P.L.; Mounzer, K.C.; De Wet, J.J.; DeJesus, E.; Jessen, H.; Grant, R.M.; Ruane, P.J.; et al. Emtricitabine and tenofovir alafenamide vs emtricitabine and tenofovir disoproxil fumarate for HIV pre-exposure prophylaxis (DISCOVER): Primary results from a randomised, double-blind, multicentre, active-controlled, phase 3, non-inferiority trial. Lancet 2020, 396, 239–254. [Google Scholar] [CrossRef] [PubMed]
- Grant, R.M.; Lama, J.R.; Anderson, P.L.; McMahan, V.; Liu, A.Y.; Vargas, L.; Goicochea, P.; Casapía, M.; Guanira-Carranza, J.V.; Ramirez-Cardich, M.E.; et al. Preexposure Chemoprophylaxis for HIV Prevention in Men Who Have Sex with Men. N. Engl. J. Med. 2010, 363, 2587–2599. [Google Scholar] [CrossRef] [PubMed]
- Baeten, J.M.; Palanee-Phillips, T.; Brown, E.R.; Schwartz, K.; Soto-Torres, L.E.; Govender, V.; Mgodi, N.M.; Matovu Kiweewa, F.; Nair, G.; Mhlanga, F.; et al. Use of a Vaginal Ring Containing Dapivirine for HIV-1 Prevention in Women. N. Engl. J. Med. 2016, 375, 2121–2132. [Google Scholar] [CrossRef] [PubMed]
- Nel, A.; van Niekerk, N.; Kapiga, S.; Bekker, L.-G.; Gama, C.; Gill, K.; Kamali, A.; Kotze, P.; Louw, C.; Mabude, Z.; et al. Safety and Efficacy of a Dapivirine Vaginal Ring for HIV Prevention in Women. N. Engl. J. Med. 2016, 375, 2133–2143. [Google Scholar] [CrossRef] [PubMed]
- Markowitz, M.; Grossman, H.; Anderson, P.L.; Grant, R.; Gandhi, M.; Horng, H.; Mohri, H. Newly Acquired Infection with Multidrug-Resistant HIV-1 in a Patient Adherent to Preexposure Prophylaxis. JAIDS J. Acquir. Immune Defic. Syndr. 2017, 76, e104–e106. [Google Scholar] [CrossRef]
- Rivera, A.S.; Pak, K.J.; Mefford, M.T.; Hechter, R.C. Use of Tenofovir Alafenamide Fumarate for HIV Pre-Exposure Prophylaxis and Incidence of Hypertension and Initiation of Statins. JAMA Netw. Open 2023, 6, e2332968. [Google Scholar] [CrossRef]
- Casazza, J.P.; Cale, E.M.; Narpala, S.; Yamshchikov, G.V.; Coates, E.E.; Hendel, C.S.; Novik, L.; Holman, L.A.; Widge, A.T.; Apte, P.; et al. Safety and tolerability of AAV8 delivery of a broadly neutralizing antibody in adults living with HIV: A phase 1, dose-escalation trial. Nat. Med. 2022, 28, 1022–1030. [Google Scholar] [CrossRef]
- Corey, L.; Gilbert, P.B.; Juraska, M.; Montefiori, D.C.; Morris, L.; Karuna, S.T.; Edupuganti, S.; Mgodi, N.M.; Decamp, A.C.; Rudnicki, E.; et al. Two Randomized Trials of Neutralizing Antibodies to Prevent HIV-1 Acquisition. N. Engl. J. Med. 2021, 384, 1003–1014. [Google Scholar] [CrossRef]
- Pejchal, R.; Doores, K.J.; Walker, L.M.; Khayat, R.; Huang, P.-S.; Wang, S.-K.; Stanfield, R.L.; Julien, J.-P.; Ramos, A.; Crispin, M.; et al. A Potent and Broad Neutralizing Antibody Recognizes and Penetrates the HIV Glycan Shield. Science 2011, 334, 1097–1103. [Google Scholar] [CrossRef]
- McCoy, L.E.; Burton, D.R. Identification and specificity of broadly neutralizing antibodies against HIV. Immunol. Rev. 2017, 275, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Margolis, D.M.; Koup, R.A.; Ferrari, G. HIV antibodies for treatment of HIV infection. Immunol. Rev. 2017, 275, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Kang, B.H.; Ishida, E.; Zhou, T.; Griesman, T.; Sheng, Z.; Wu, F.; Doria-Rose, N.A.; Zhang, B.; McKee, K.; et al. Identification of a CD4-Binding-Site Antibody to HIV that Evolved Near-Pan Neutralization Breadth. Immunity 2016, 45, 1108–1121. [Google Scholar] [CrossRef] [PubMed]
- Julien, J.-P.; Sok, D.; Khayat, R.; Lee, J.H.; Doores, K.J.; Walker, L.M.; Ramos, A.; Diwanji, D.C.; Pejchal, R.; Cupo, A.; et al. Broadly Neutralizing Antibody PGT121 Allosterically Modulates CD4 Binding via Recognition of the HIV-1 gp120 V3 Base and Multiple Surrounding Glycans. PLoS Pathog. 2013, 9, e1003342. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Ofek, G.; Laub, L.; Louder, M.K.; Doria-Rose, N.A.; Longo, N.S.; Imamichi, H.; Bailer, R.T.; Chakrabarti, B.; Sharma, S.K.; et al. Broad and potent neutralization of HIV-1 by a gp41-specific human antibody. Nature 2012, 491, 406–412. [Google Scholar] [CrossRef] [PubMed]
- Sok, D.; van Gils, M.J.; Pauthner, M.; Julien, J.-P.; Saye-Francisco, K.L.; Hsueh, J.; Briney, B.; Lee, J.H.; Le, K.M.; Lee, P.S.; et al. Recombinant HIV envelope trimer selects for quaternary-dependent antibodies targeting the trimer apex. Proc. Natl. Acad. Sci. USA 2014, 111, 17624–17629. [Google Scholar] [CrossRef]
- Xu, L.; Pegu, A.; Rao, E.; Doria-Rose, N.; Beninga, J.; McKee, K.; Lord, D.M.; Wei, R.R.; Deng, G.; Louder, M.; et al. Trispecific broadly neutralizing HIV antibodies mediate potent SHIV protection in macaques. Science 2017, 358, 85–90. [Google Scholar] [CrossRef]
- Song, R.; Franco, D.; Kao, C.-Y.; Yu, F.; Huang, Y.; Ho David, D. Epitope Mapping of Ibalizumab, a Humanized Anti-CD4 Monoclonal Antibody with Anti-HIV-1 Activity in Infected Patients. J. Virol. 2010, 84, 6935–6942. [Google Scholar] [CrossRef]
- Trkola, A.; Ketas Thomas, J.; Nagashima Kirsten, A.; Zhao, L.; Cilliers, T.; Morris, L.; Moore, J.P.; Maddon, P.J.; Olson, W.C. Potent, Broad-Spectrum Inhibition of Human Immunodeficiency Virus Type 1 by the CCR5 Monoclonal Antibody PRO 140. J. Virol. 2001, 75, 579–588. [Google Scholar] [CrossRef]
- Bar, K.J.; Sneller, M.C.; Harrison, L.J.; Justement, J.S.; Overton, E.T.; Petrone, M.E.; Salantes, D.B.; Seamon, C.A.; Scheinfeld, B.; Kwan, R.W.; et al. Effect of HIV Antibody VRC01 on Viral Rebound after Treatment Interruption. N. Engl. J. Med. 2016, 375, 2037–2050. [Google Scholar] [CrossRef]
- Khan Salar, N.; Sok, D.; Tran, K.; Movsesyan, A.; Dubrovskaya, V.; Burton Dennis, R.; Wyatt, R.T. Targeting the HIV-1 Spike and Coreceptor with Bi- and Trispecific Antibodies for Single-Component Broad Inhibition of Entry. J. Virol. 2018, 92, e00384-18. [Google Scholar] [PubMed]
- Steinhardt, J.J.; Guenaga, J.; Turner, H.L.; McKee, K.; Louder, M.K.; O’Dell, S.; Chiang, C.-I.; Lei, L.; Galkin, A.; Andrianov, A.K.; et al. Rational design of a trispecific antibody targeting the HIV-1 Env with elevated anti-viral activity. Nat. Commun. 2018, 9, 877. [Google Scholar] [CrossRef] [PubMed]
- Pegu, A.; Xu, L.; DeMouth, M.E.; Fabozzi, G.; March, K.; Almasri, C.G.; Cully, M.D.; Wang, K.; Yang, E.S.; Dias, J.; et al. Potent anti-viral activity of a trispecific HIV neutralizing antibody in SHIV-infected monkeys. Cell Rep. 2022, 38, 110199. [Google Scholar] [CrossRef] [PubMed]
- Zhai, L.; Zhang, L.; Jiang, Y.; Li, B.; Yang, M.; Victorovich, K.V.; Aleksandrovna, K.T.; Li, M.; Wang, Y.; Huang, D.; et al. Broadly neutralizing antibodies recognizing different antigenic epitopes act synergistically against the influenza B virus. J. Med. Virol. 2023, 95, e28106. [Google Scholar] [CrossRef] [PubMed]
- Perdomo-Celis, F.; Medina-Moreno, S.; Davis, H.; Bryant, J.; Zapata, J.C. HIV Replication in Humanized IL-3/GM-CSF-Transgenic NOG Mice. Pathogens 2019, 8, 33. [Google Scholar] [CrossRef] [PubMed]
- Rosás-Umbert, M.; Gunst, J.D.; Pahus, M.H.; Olesen, R.; Schleimann, M.; Denton, P.W.; Ramos, V.; Ward, A.; Kinloch, N.N.; Copertino, D.C.; et al. Administration of broadly neutralizing anti-HIV-1 antibodies at ART initiation maintains long-term CD8+ T cell immunity. Nat. Commun. 2022, 13, 6473. [Google Scholar] [CrossRef] [PubMed]
- Waters, L.; de Miguel-Buckley, R.; Poulin Sb Arribas, J.R. Broadly Neutralizing Antibodies for Human Immunodeficiency Virus Treatment: Broad in Theory, Narrow in Reality. Clin. Infect. Dis. 2023, 76, 1136–1141. [Google Scholar] [CrossRef]
- Sneller, M.C.; Blazkova, J.; Justement, J.S.; Shi, V.; Kennedy, B.D.; Gittens, K.; Tolstenko, J.; McCormack, G.; Whitehead, E.J.; Schneck, R.F.; et al. Combination anti-HIV antibodies provide sustained virological suppression. Nature 2022, 606, 375–381. [Google Scholar] [CrossRef]
- Julg, B.; Stephenson, K.E.; Wagh, K.; Tan, S.C.; Zash, R.; Walsh, S.; Ansel, J.; Kanjilal, D.; Nkolola, J.; Walker-Sperling, V.E.K.; et al. Safety and antiviral activity of triple combination broadly neutralizing monoclonal antibody therapy against HIV-1: A phase 1 clinical trial. Nat. Med. 2022, 28, 1288–1296. [Google Scholar] [CrossRef]
- Zhang, B.; Gorman, J.; Kwon, Y.D.; Pegu, A.; Chao, C.W.; Liu, T.; Asokan, M.; Bender, M.F.; Bylund, T.; Damron, L.; et al. Bispecific antibody CAP256.J3LS targets V2-apex and CD4-binding sites with high breadth and potency. mAbs 2023, 15, 2165390. [Google Scholar] [CrossRef]
- Montefiori, D.C. Bispecific Antibodies Against HIV. Cell 2016, 165, 1563–1564. [Google Scholar] [CrossRef] [PubMed]
- Paneerselvam, N.; Khan, A.; Lawson, B.R. Broadly neutralizing antibodies targeting HIV: Progress and challenges. Clin. Immunol. 2023, 257, 109809. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.; Fang, C.; Liu, J.; Yu, H.; Qi, J.; Zhang, Z.; Yuan, R.; Xiong, D.; Gao, S.; Yuan, Y.A.; et al. Molecular insights into the inhibition of HIV-1 infection using a CD4 domain-1-specific monoclonal antibody. Antivir. Res. 2015, 122, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Lai, Y. Monoclonal CCR5 Antibody: A Promising Therapy for HIV. Curr. HIV Res. 2023, 21, 1–8. [Google Scholar]
- Dimitrov, A. Ibalizumab, a CD4-specific mAb to inhibit HIV-1 infection. Curr. Opin Investig. Drugs. 2007, 8, 653–661. [Google Scholar] [PubMed]
- Poli, G. PRO-140 (Progenics). IDrugs Investig. Drugs J. 2001, 4, 1068–1071. [Google Scholar]
- Goo, L.; Jalalian-Lechak, Z.; Richardson Barbra, A.; Overbaugh, J. A Combination of Broadly Neutralizing HIV-1 Monoclonal Antibodies Targeting Distinct Epitopes Effectively Neutralizes Variants Found in Early Infection. J. Virol. 2012, 86, 10857–10861. [Google Scholar] [CrossRef]
- Huang, Y.; Yu, J.; Lanzi, A.; Yao, X.; Andrews, C.D.; Tsai, L.; Gajjar, M.R.; Sun, M.; Seaman, M.S.; Padte, N.N.; et al. Engineered Bispecific Antibodies with Exquisite HIV-1-Neutralizing Activity. Cell 2016, 165, 1621–1631. [Google Scholar] [CrossRef]
- Wu, X.; Guo, J.; Niu, M.; An, M.; Liu, L.; Wang, H.; Jin, X.; Zhang, Q.; Lam, K.S.; Wu, T.; et al. Tandem bispecific neutralizing antibody eliminates HIV-1 infection in humanized mice. J. Clin. Investig. 2018, 128, 2239–2251. [Google Scholar] [CrossRef]
- Promsote, W.; Xu, L.; Hataye, J.; Fabozzi, G.; March, K.; Almasri, C.G.; DeMouth, M.E.; Lovelace, S.E.; Talana, C.A.; Doria-Rose, N.A.; et al. Trispecific antibody targeting HIV-1 and T cells activates and eliminates latently-infected cells in HIV/SHIV infections. Nat. Commun. 2023, 14, 3719. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, J.; Zhai, L.; Liang, Z.; Chen, X.; Jiang, Y.; Lin, Y.; Feng, S.; Liu, Y.; Zhao, W.; Wang, F. Rational Design and Characterization of Trispecific Antibodies Targeting the HIV-1 Receptor and Envelope Glycoprotein. Vaccines 2024, 12, 19. https://doi.org/10.3390/vaccines12010019
Liang J, Zhai L, Liang Z, Chen X, Jiang Y, Lin Y, Feng S, Liu Y, Zhao W, Wang F. Rational Design and Characterization of Trispecific Antibodies Targeting the HIV-1 Receptor and Envelope Glycoprotein. Vaccines. 2024; 12(1):19. https://doi.org/10.3390/vaccines12010019
Chicago/Turabian StyleLiang, Jinhu, Linlin Zhai, Zuxin Liang, Xiaoling Chen, Yushan Jiang, Yuanlong Lin, Shiyan Feng, Yingxia Liu, Wei Zhao, and Fuxiang Wang. 2024. "Rational Design and Characterization of Trispecific Antibodies Targeting the HIV-1 Receptor and Envelope Glycoprotein" Vaccines 12, no. 1: 19. https://doi.org/10.3390/vaccines12010019
APA StyleLiang, J., Zhai, L., Liang, Z., Chen, X., Jiang, Y., Lin, Y., Feng, S., Liu, Y., Zhao, W., & Wang, F. (2024). Rational Design and Characterization of Trispecific Antibodies Targeting the HIV-1 Receptor and Envelope Glycoprotein. Vaccines, 12(1), 19. https://doi.org/10.3390/vaccines12010019