

Clinical and Experimental Determination of Protection Afforded by BCG Vaccination against Infection with Non-Tuberculous Mycobacteria: A Role in Cystic Fibrosis?

 , ,
, , 

Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Animals
2.3. Mouse Immunisation and Infection
2.4. Cell Isolation and Flow Cytometry
2.5. Comparison of BCG Vaccinated and Unvaccinated CF Cohorts
2.6. Statistical Analysis
3. Results
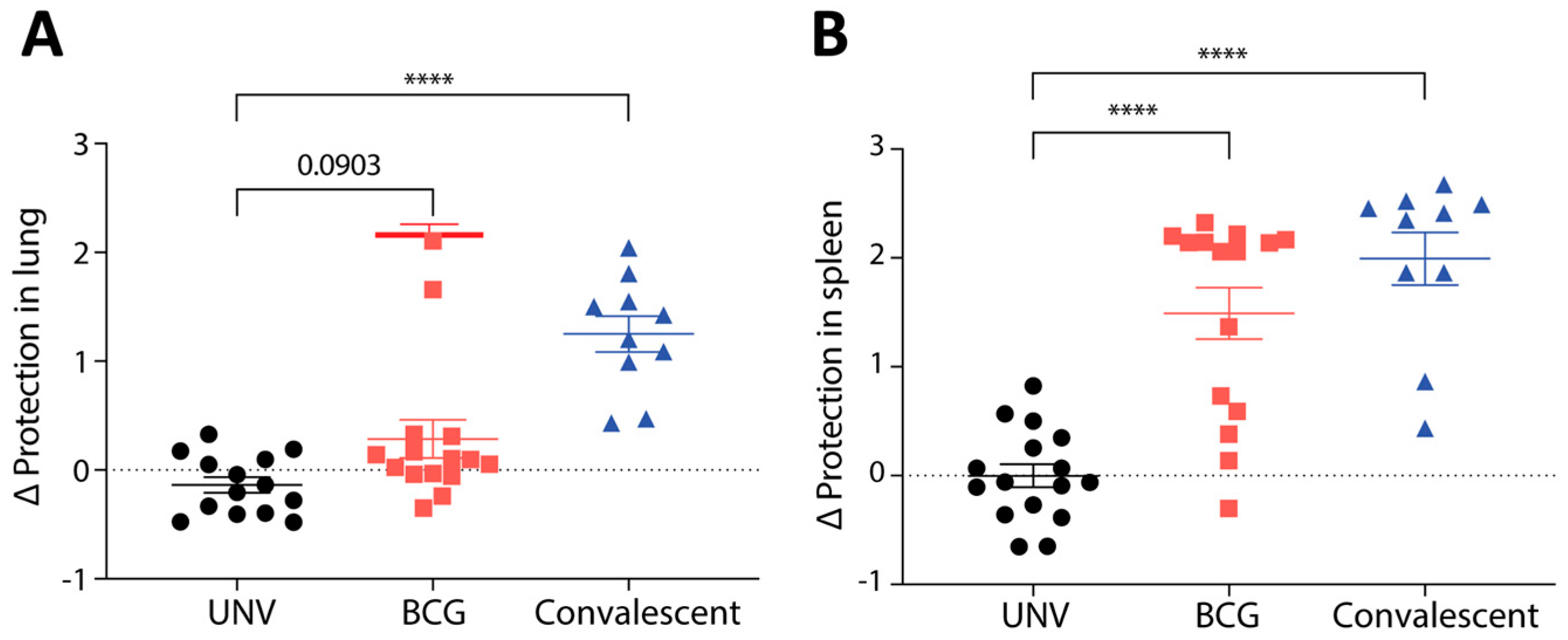
3.1. Protective Immunity Afforded by BCG Vaccination in Mice
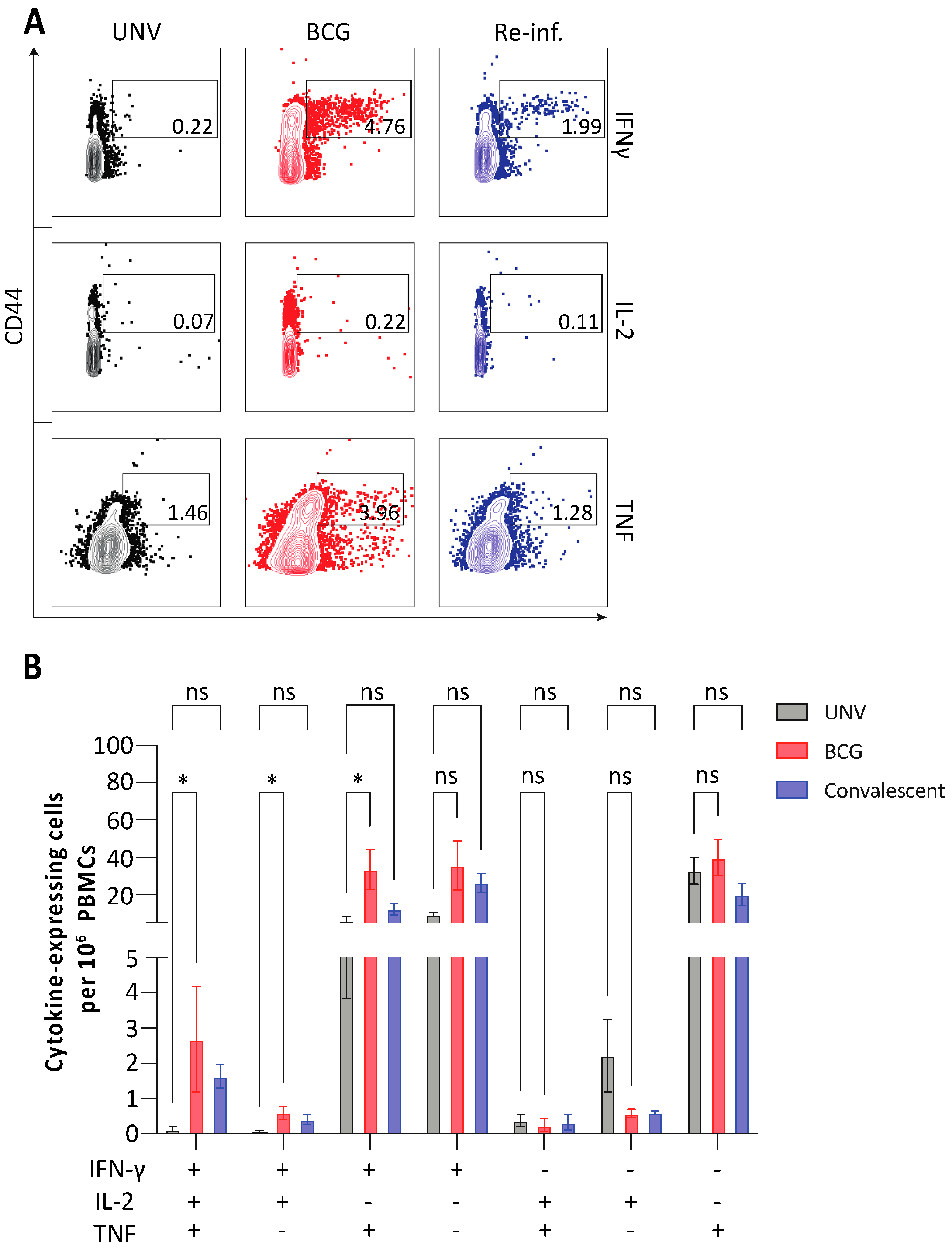
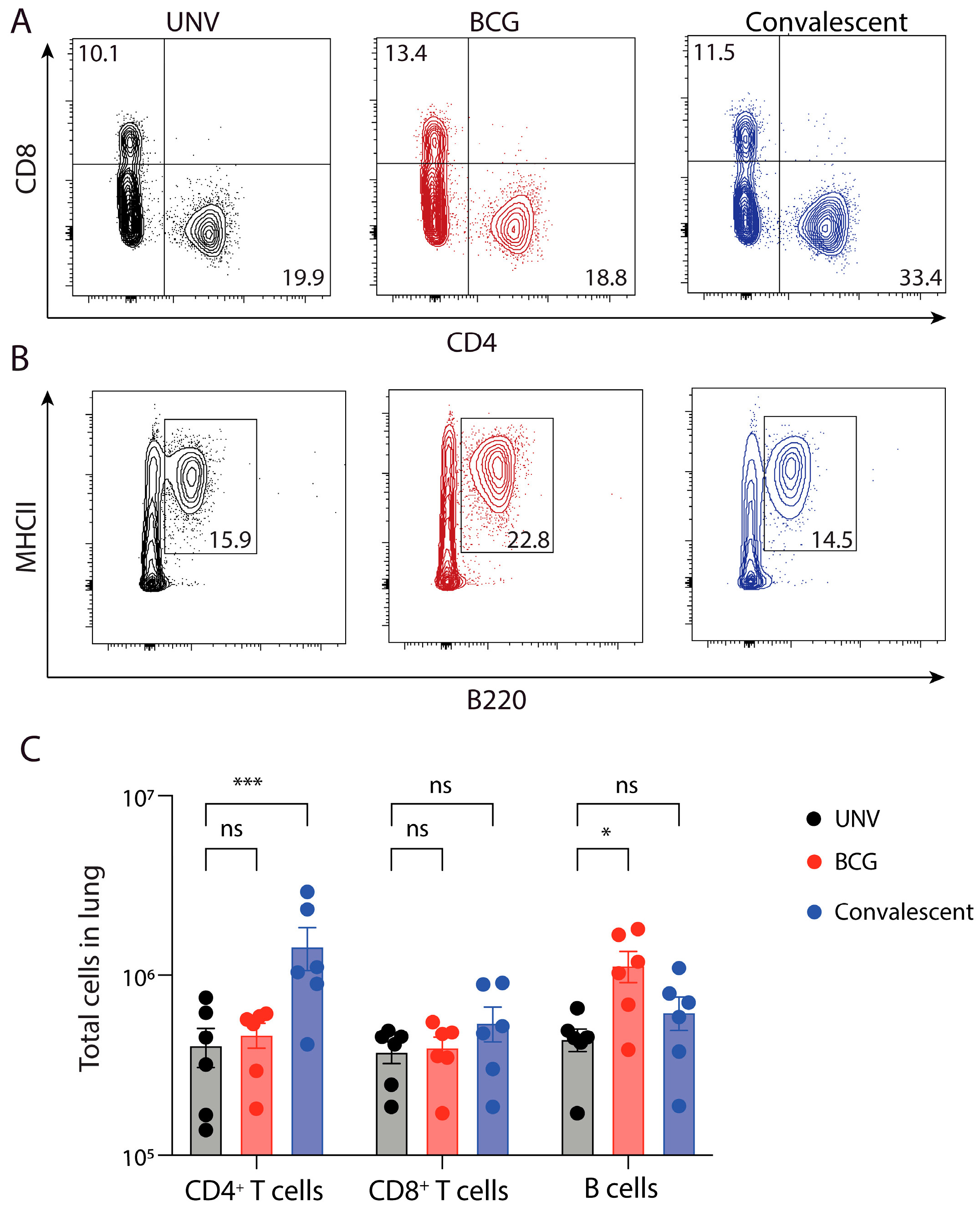
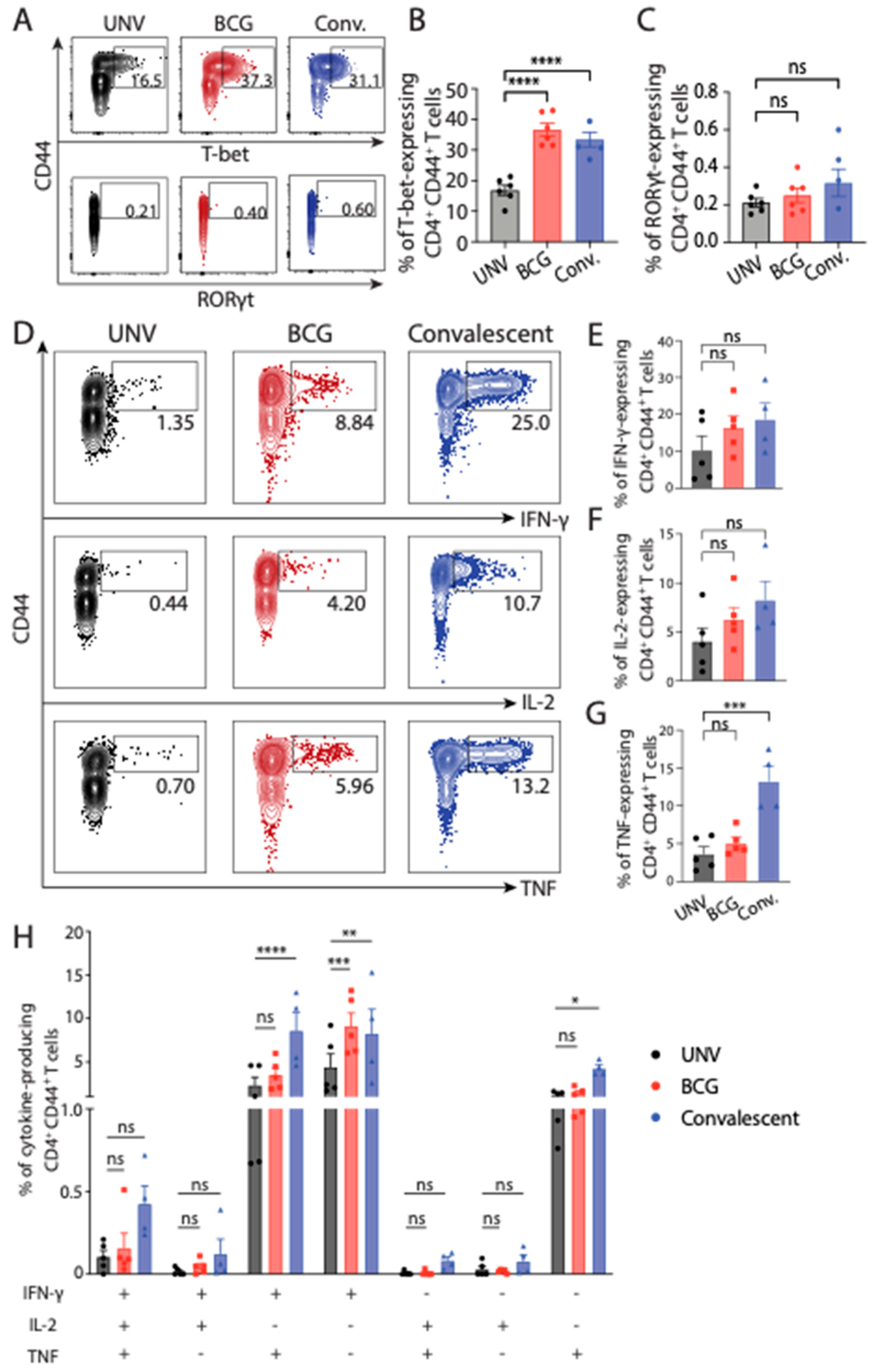
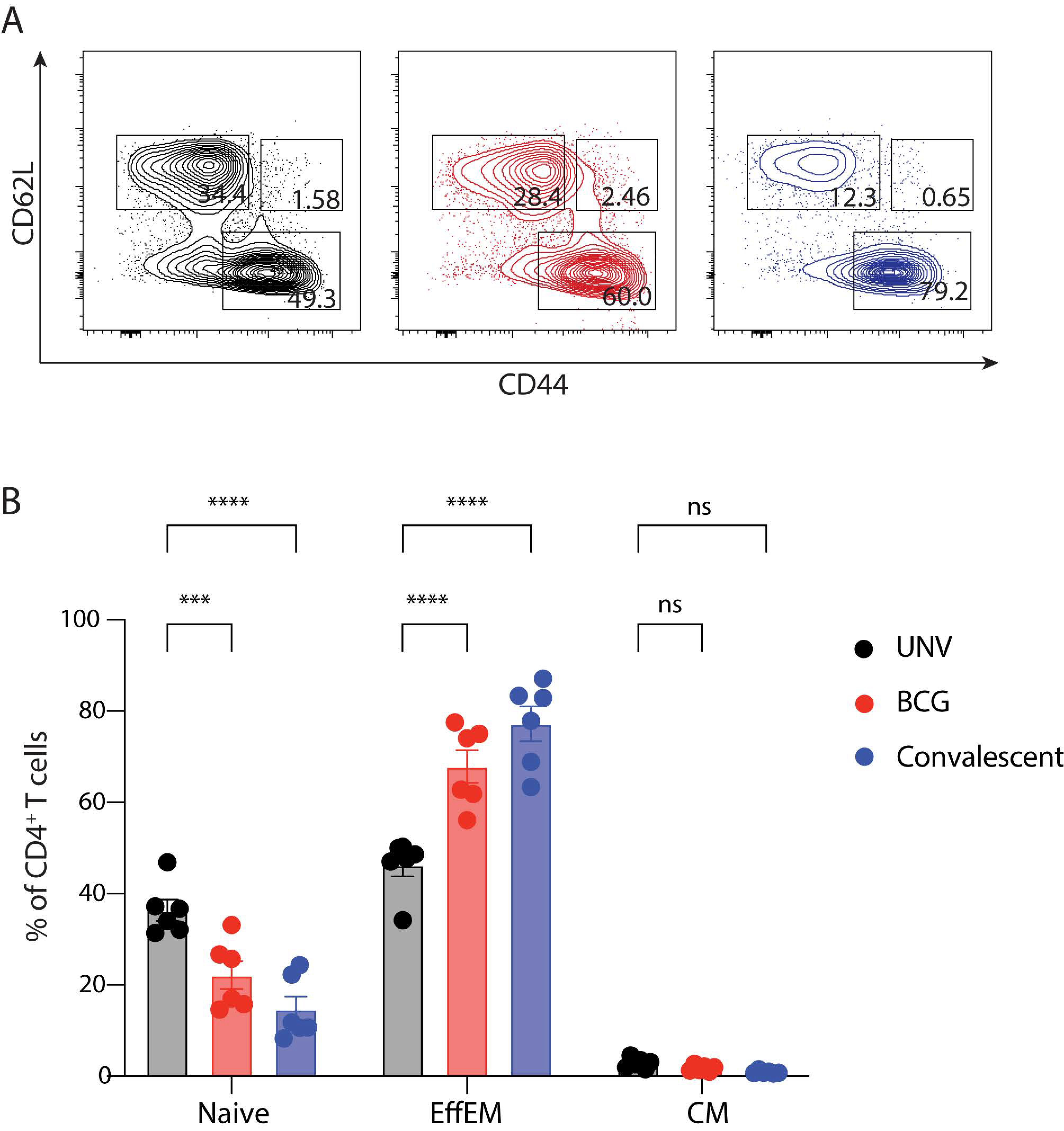
3.2. Adaptive Immune Cell Response after BCG Vaccination and Protection against M. abscessus Challenge in Mice
3.3. Comparison of NTM Infection in Relation to BCG Vaccination Status
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martiniano, S.L.; Nick, J.A.; Daley, C.L. Nontuberculous mycobacterial infections in cystic fibrosis. Thorac. Surg. Clin. 2019, 29, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Griffith, D.E.; Aksamit, T.; Brown-Elliott, B.A.; Catanzaro, A.; Daley, C.; Gordin, F.; Holland, S.M.; Horsburgh, R.; Huitt, G.; Iademarco, M.F.; et al. An official ATS/IDSA statement: Diagnosis, treatment, and prevention of nontuberculous mycobacterial diseases. Am. J. Respir. Crit. Care Med. 2007, 175, 367–416. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.R.; Sheng, W.H.; Hung, C.C.; Yu, C.J.; Lee, L.N.; Hsueh, P.R. Mycobacterium abscessus Complex infections in humans. Emerg. Infect. Dis. 2015, 21, 1638–1646. [Google Scholar] [CrossRef] [PubMed]
- Jhun, B.W.; Moon, S.M.; Jeon, K.; Kwon, O.J.; Yoo, H.; Carriere, K.C.; Huh, H.J.; Lee, N.Y.; Shin, S.J.; Daley, C.L.; et al. Prognostic factors associated with long-term mortality in 1445 patients with nontuberculous mycobacterial pulmonary disease: A 15-year follow-up study. Eur. Respir. J. 2020, 55, 1900798. [Google Scholar] [CrossRef]
- Marty, P.K.; Yetmar, Z.A.; Gerberi, D.J.; Escalante, P.; Pennington, K.M.; Mahmood, M. Risk factors and outcomes of non-tuberculous mycobacteria infection in lung transplant recipients: A systematic review and meta-analysis. J. Heart Lung Transplant. 2023, 42, 264–274. [Google Scholar] [CrossRef]
- Degiacomi, G.; Sammartino, J.C.; Chiarelli, L.R.; Riabova, O.; Makarov, V.; Pasca, M.R. Mycobacterium abscessus, an emerging and worrisome pathogen among cystic fibrosis patients. Int. J. Mol. Sci. 2019, 20, 5868. [Google Scholar] [CrossRef]
- Lu, M.; Saddi, V.; Britton, P.N.; Selvadurai, H.; Robinson, P.D.; Pandit, C.; Marais, B.J.; Fitzgerald, D.A. Disease caused by non-tuberculous mycobacteria in children with cystic fibrosis. Paediatr. Respir. Rev. 2019, 29, 42–52. [Google Scholar] [CrossRef]
- Choi, H.; Jhun, B.W.; Kim, S.-Y.; Kim, D.H.; Lee, H.; Jeon, K.; Kwon, O.J.; Huh, H.J.; Ki, C.-S.; Lee, N.Y.; et al. Treatment outcomes of macrolide-susceptible Mycobacterium abscessus lung disease. Diagn. Microbiol. Infect. Dis. 2018, 90, 293–295. [Google Scholar] [CrossRef]
- Mirsaeidi, M.; Allen, M.B.; Ebrahimi, G.; Schraufnagel, D. Hospital costs in the US for pulmonary mycobacterial diseases. Int. J. Mycobacteriol. 2015, 4, 217–221. [Google Scholar] [CrossRef]
- WHO. Impact of the COVID-19 Pandemic on TB Detection and Mortality; World Health Organisation: Geneva, Switzerland, 2021. [Google Scholar]
- Calmette, A.; Plotz, H. Protective inoculation against tuberculosis with BCG. Am. Rev. Tuberc. 1929, 19, 567–572. [Google Scholar]
- Kilpeläinen, A.; Maya-Hoyos, M.; Saubí, N.; Soto, C.Y.; Joseph Munne, J. Advances and challenges in recombinant Mycobacterium bovis BCG-based HIV vaccine development: Lessons learned. Expert. Rev. Vaccines 2018, 17, 1005–1020. [Google Scholar] [CrossRef] [PubMed]
- Moliva, J.I.; Turner, J.; Torrelles, J.B. Immune responses to Bacillus Calmette–Guérin vaccination: Why do they fail to protect against Mycobacterium tuberculosis? Front. Immunol. 2017, 8, 407. [Google Scholar] [CrossRef] [PubMed]
- Colditz, G.; Berkey, C.S.; Mosteller, F.; Brewer, T.F.; Wilson, M.E.; Burdick, E.; Fineberg, H.V. The efficacy of bacillus Calmette-Guerin vaccination of newborns and infants in the prevention of tuberculosis: Meta-analyses of the published literature. Pediatrics 1995, 96, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Schoenmakers, A.; Mieras, L.; Budiawan, T.; van Brakel, W.H. The state of affairs in post-exposure leprosy prevention: A descriptive meta-analysis on immuno- and chemo-prophylaxis. Res. Rep. Trop. Med. 2020, 11, 97–117. [Google Scholar] [CrossRef] [PubMed]
- Pittet, L.F.; Tebruegge, M.; Dutta, B.; Donath, S.; Messina, N.; Casalaz, D.; Hanekom, W.A.; Britton, W.J.; Robins-Browne, R.; Curtis, N.; et al. Mycobacterium ulcerans-specific immune response after immunisation with bacillus Calmette-Guérin (BCG) vaccine. Vaccine 2021, 39, 652–657. [Google Scholar] [CrossRef]
- Trnka, L.; Daňková, D.; Švandová, E. Six years’ experience with the discontinuation of BCG vaccination: 4. Protective effect of BCG vaccination against the Mycobacterium avium intracellulare complex. Int. J. Tuberc. Lung Dis. 1994, 75, 348–352. [Google Scholar] [CrossRef]
- Abate, G.; Hamzabegovic, F.; Eickhoff, C.S.; Hoft, D.F. BCG vaccination induces M. avium and M. abscessus cross-protective immunity. Front. Immunol. 2019, 10, 234. [Google Scholar] [CrossRef]
- Mousset, C.M.; Hobo, W.; Woestenenk, R.; Preijers, F.; Dolstra, H.; van der Waart, A.B. Comprehensive phenotyping of T cells using flow cytometry. Cytometry A 2019, 95, 647–654. [Google Scholar] [CrossRef]
- Dockrell, H.M.; Smith, S.G. What Have We Learnt about BCG Vaccination in the Last 20 Years? Front. Immunol. 2017, 8, 1134. [Google Scholar] [CrossRef]
- Darrah, P.A.; Zeppa, J.J.; Maiello, P.; Hackney, J.A.; Wadsworth, M.H.; Hughes, T.K.; Seder, R.A. Prevention of tuberculosis in macaques after intravenous BCG immunization. Nature 2020, 577, 95–102. [Google Scholar] [CrossRef]
- Shu, C.-C.; Wu, L.S.-H.; Wu, M.-F.; Lai, H.-C.; Wang, P.-H.; Cheng, S.-L.; Wang, J.-Y.; Yu, C.-J. Mono- and poly-functional T cells in nontuberculous mycobacteria lung disease patients: Implications in analyzing risk of disease progression. Cytokine 2019, 120, 176–185. [Google Scholar] [CrossRef]
- Day, C.L.; Abrahams, D.A.; Lerumo, L.; van Rensburg, E.J.; Stone, L.; O’rie, T.; Pienaar, B.; de Kock, M.; Kaplan, G.; Mahomed, H.; et al. Functional capacity of Mycobacterium tuberculosis-specific T cell responses in humans is associated with mycobacterial load. J. Immunol. 2011, 187, 2222–2232. [Google Scholar] [CrossRef]
- Tameris, M.D.; Hatherill, M.; Landry, B.S.; Scriba, T.J.; Snowden, M.A.; Lockhart, S.; Shea, J.E.; McClain, J.B.; Hussey, G.D.; Hanekom, W.A.; et al. Safety and efficacy of MVA85A, a new tuberculosis vaccine, in infants previously vaccinated with BCG: A randomised, placebo-controlled phase 2b trial. Lancet 2013, 381, 1021–1028. [Google Scholar] [CrossRef]
- Kaech, S.M.; Wherry, E.J.; Ahmed, R. Effector and memory T-cell differentiation: Implications for vaccine development. Nat. Rev. Immunol. 2002, 2, 251–262. [Google Scholar] [CrossRef]
- Kozakiewicz, L.; Chen, Y.; Xu, J.; Wang, Y.; Dunussi-Joannopoulos, K.; Ou, Q.; Flynn, J.L.; Porcelli, S.A.; Jacobsr, W.R., Jr.; Chan, J. B cells regulate neutrophilia during Mycobacterium tuberculosis infection and BCG vaccination by modulating the interleukin-17 response. PLoS Pathog. 2013, 9, e1003472. [Google Scholar] [CrossRef]
- Tanner, R.; Villarreal-Ramos, B.; Vordermeier, H.M.; McShane, H. The Humoral Immune Response to BCG Vaccination. Front. Immunol. 2019, 10, 1317. [Google Scholar] [CrossRef]
- de Vallière, S.; Abate, G.; Blazevic, A.; Heuertz, R.M.; Hoft, D.F. Enhancement of innate and cell-mediated immunity by antimycobacterial antibodies. Infect. Immun. 2005, 73, 6711–6720. [Google Scholar] [CrossRef]
- Chen, T.; Blanc, C.; Eder, A.Z.; Prados-Rosales, R.; Souza, A.C.O.; Kim, R.S.; Glatman-Freedman, A.; Joe, M.; Bai, Y.; Lowary, T.L.; et al. Association of human antibodies to arabinomannan With enhanced mycobacterial opsonophagocytosis and intracellular growth reduction. J. Infect. Dis. 2016, 214, 300–310. [Google Scholar] [CrossRef]
- Tural, D.A.; Emiralioglu, N.; Ozsezen, B.; Saribas, Z.; Ozcan, N.; Alp, A.; Sunman, B.; Hizal, M.; Polat, S.E.; Yalcin, E.; et al. The frequency and related factors of non-tuberculosis mycobacteria infections among patients with cystic fibrosis. Pediatr. Int. 2021, 63, 1369–1375. [Google Scholar] [CrossRef]
- Davenne, T.; McShane, H. Why don’t we have an effective tuberculosis vaccine yet? Expert. Rev. Vaccines. 2016, 15, 1009–1013. [Google Scholar] [CrossRef]
- Mangtani, P.; Abubakar, I.; Ariti, C.; Beynon, R.; Pimpin, L.; Fine, P.E.M.; Rodrigues, L.C.; Smith, P.G.; Lipman, M.; Whiting, P.F.; et al. Protection by BCG vaccine against tuberculosis: A systematic review of randomized controlled trials. Clin. Infect. Dis. 2014, 58, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Hoefsloot, W.; van Ingen, J.; Andrejak, C.; Ängeby, K.; Bauriaud, R.; Bemer, P.; Beylis, N.; Boeree, M.J.; Cacho, J.; Chihota, V.; et al. The geographic diversity of nontuberculous mycobacteria isolated from pulmonary samples: An NTM-NET collaborative study. Eur. Respir. J. 2013, 42, 1604–1613. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RCWMCH 1, Cape Town, South Africa | CHW 2, Sydney, Australia | |||||||
|---|---|---|---|---|---|---|---|---|
| Year 1 | Year 2 | Year 3 | Overall | Year 1 | Year 2 | Year 3 | Overall | |
| No. patients | 71 | 80 | 82 | 91 | 208 | 208 | 213 | 231 |
| % males | 51 | 49 | 50 | 49 | 51 | 51 | 52 | 52 |
| Age (years) | 8.8 | 8.9 | 9.7 | 9.4 | 9.1 | 9.4 | 9.7 | 9.8 |
| Best 3 FEV1 (% predicted) | 88.0 | 86.3 | 85.4 | 91.3 | 95.6 * | 96.5 * | 94.7 * | 98.7 * |
| Best Height (z score) | −0.91 | −0.89 | −0.86 | −0.57 | −0.08 * | −0.13 * | −0.09 * | 0.06 * |
| Best Weight (z score) | −0.78 | −0.74 | −0.72 | −0.45 | −0.01 * | −0.03 * | −0.03 * | 0.18 * |
| Best 4 BMI mean (z score) | −0.22 | −0.26 | −0.27 | 0.04 | 0.06 | 0.06 * | 0.11 * | 0.27 |
| RCWMCH 1, Cape Town, South Africa | CHW 2, Sydney, Australia | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Year 1 | Year 2 | Year 3 | Overall | Year 1 | Year 2 | Year 3 | Overall | ||
| Number tested for NTM (%) | 43 (61) | 41 (51) | 55 (67) | 77 (85) | 129 (62) | 139 (67) | 131 (62) | 193 (84) | |
| Any NTM | Intermittent (%) | 2.33 | 4.88 | 1.82 | 5.19 | 4.65 | 5.04 | 6.11 | 7.77 |
| colonised (%) | 0 | 0 | 0 | 0 | 3.88 | 5.76 | 6.11 | 6.74 * | |
| any isolation (%) | 2.33 | 4.88 | 1.82 | 5.19 | 8.53 | 10.79 | 12.21 | 11.92 | |
| M. abscessus | Intermittent (%) | 2.33 | 0 | 0 | 1.30 | 0.78 | 0 | 3.05 | 2.59 |
| colonised (%) | 0 | 0 | 0 | 0 | 3.10 | 5.04 | 6.11 | 5.70 * | |
| any isolation (%) | 2.33 | 0 | 0 | 1.30 | 3.88 | 5.04 | 9.16 * | 7.25 | |
| M. avium | Intermittent (%) | 2.33 | 2.44 | 1.82 | 3.90 | 3.88 | 3.60 | 3.05 | 4.66 |
| colonised (%) | 0 | 0 | 0 | 0 | 1.55 | 1.44 | 0 | 1.55 | |
| any isolation (%) | 2.33 | 2.44 | 1.82 | 3.90 | 5.43 | 5.04 | 3.05 | 5.70 | |
| M. intracellulare | Intermittent (%) | 0 | 4.88 | 1.82 | 3.90 | 0 | 1.44 | 0.76 | 1.55 |
| colonised (%) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| any isolation (%) | 0 | 4.88 | 1.82 | 3.90 | 0 | 1.44 | 0.76 | 1.55 | |
| Number tested for other bacteria (% sampled) | 71 (100) | 80 (100) | 82 (100) | 91 (100) | 208 (100) | 208 (100) | 213 (100) | 231 (100) | |
| Pseudomonas aeruginosa | Intermittent (%) | 28.17 | 18.75 | 29.27 | 45.05 | 19.71 | 14.90 | 15.49 * | 35.50 |
| colonised (%) | 16.90 | 18.75 | 14.63 | 28.57 | 13.94 | 12.02 | 9.39 | 19.05 | |
| Staphylococcus aureus | Intermittent (%) | 64.79 | 66.25 | 67.70 | 84.62 | 74.04 | 78.85 * | 80.75 * | 90.48 |
| Haemophilus spp. | Intermittent (%) | 12.68 | 15.00 | 12.20 | 27.47 | 16.83 | 10.58 | 11.74 | 28.14 |
| Aspergillus spp. | Intermittent (%) | 25.35 | 21.25 | 20.73 | 32.97 | 22.60 | 25.48 | 23.00 | 37.66 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Warner, S.; Blaxland, A.; Counoupas, C.; Verstraete, J.; Zampoli, M.; Marais, B.J.; Fitzgerald, D.A.; Robinson, P.D.; Triccas, J.A. Clinical and Experimental Determination of Protection Afforded by BCG Vaccination against Infection with Non-Tuberculous Mycobacteria: A Role in Cystic Fibrosis? Vaccines 2023, 11, 1313. https://doi.org/10.3390/vaccines11081313
Warner S, Blaxland A, Counoupas C, Verstraete J, Zampoli M, Marais BJ, Fitzgerald DA, Robinson PD, Triccas JA. Clinical and Experimental Determination of Protection Afforded by BCG Vaccination against Infection with Non-Tuberculous Mycobacteria: A Role in Cystic Fibrosis? Vaccines. 2023; 11(8):1313. https://doi.org/10.3390/vaccines11081313
Chicago/Turabian StyleWarner, Sherridan, Anneliese Blaxland, Claudio Counoupas, Janine Verstraete, Marco Zampoli, Ben J. Marais, Dominic A. Fitzgerald, Paul D. Robinson, and James A. Triccas. 2023. "Clinical and Experimental Determination of Protection Afforded by BCG Vaccination against Infection with Non-Tuberculous Mycobacteria: A Role in Cystic Fibrosis?" Vaccines 11, no. 8: 1313. https://doi.org/10.3390/vaccines11081313
APA StyleWarner, S., Blaxland, A., Counoupas, C., Verstraete, J., Zampoli, M., Marais, B. J., Fitzgerald, D. A., Robinson, P. D., & Triccas, J. A. (2023). Clinical and Experimental Determination of Protection Afforded by BCG Vaccination against Infection with Non-Tuberculous Mycobacteria: A Role in Cystic Fibrosis? Vaccines, 11(8), 1313. https://doi.org/10.3390/vaccines11081313