
Degron Pathways and Leishmaniasis: Debating Potential Roles of Leishmania spp. Proteases Activity on Guiding Hosts Immune Response and Their Relevance to the Development of Vaccines

 ,
,  , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Querying for Protease Sequence Motifs in Databases
2.2. Searches for Degrons
2.3. Interaction Network Prediction
2.4. Three-Dimensional Models
3. Results
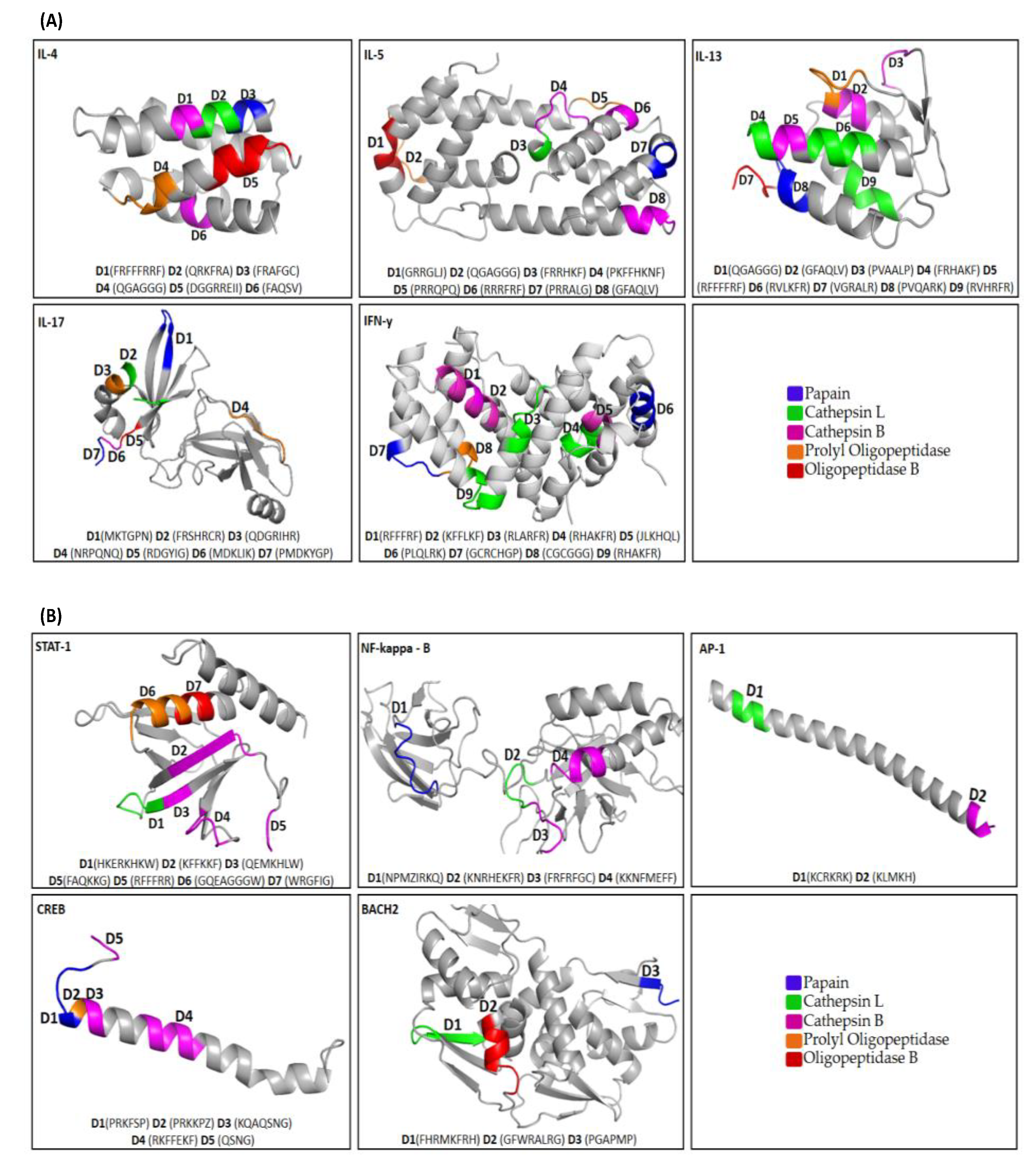
3.1. Degrons Mapping
3.2. Degrons Location
3.3. Proteins with Conserved Structures
3.4. Degrons Composition-Motifs
3.5. Network of Protease-Induced Interactions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reithinger, R.; Dujardin, J.-C.; Louzir, H.; Pirmez, C.; Alexander, B.; Brooker, S. Cutaneous leishmaniasis. Lancet Infect. Dis. 2007, 7, 581–596. [Google Scholar] [CrossRef] [PubMed]
- Kedzierski, L. Leishmaniasis vaccine: Where are we today? J. Glob. Infect. Dis. 2010, 2, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Matlashewski, G. Leishmania infection and virulence. Med. Microbiol. Immunol. 2001, 190, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Barrett, A.J. Classification of peptidases. Methods Enzymol. 1994, 244, 1–15. [Google Scholar] [CrossRef]
- McKerrow, J.H.; Caffrey, C.; Kelly, B.; Loke, P.; Sajid, M. Proteases in Parasitic Diseases. Annu. Rev. Pathol. Mech. Dis. 2006, 1, 497–536. [Google Scholar] [CrossRef]
- Silva-Lopez, R.E.; Morgado-Díaz, J.A.; Alves, C.R.; Côrte-Real, S.; Giovanni-De-Simone, S. Subcellular localization of an extracellular serine protease in Leishmania (Leishmania) amazonensis. Parasitol. Res. 2004, 93, 328–331. [Google Scholar] [CrossRef]
- Silva-Almeida, M.; Pereira, B.A.; Ribeiro-Guimarães, M.L.; Alves, C.R. Proteinases as virulence factors in Leishmania spp. infection in mammals. Parasites Vectors 2012, 5, 160. [Google Scholar] [CrossRef]
- Varshavsky, A. N-degron and C-degron pathways of protein degradation. Proc. Natl. Acad. Sci. USA 2019, 116, 358–366. [Google Scholar] [CrossRef]
- Wu, T.; Yoon, H.; Xiong, Y. Targeted protein degradation as a powerful research tool in basic biology and drug target discovery. Nat. Struct. Mol. Biol. 2020, 27, 605–614. [Google Scholar] [CrossRef]
- Ruan, H.; Yu, C.; Niu, X.; Zhang, W.; Liu, H.; Chen, L.; Xiong, R.; Sun, Q.; Jin, C.; Liu, Y.; et al. Computational strategy for intrinsically disordered protein ligand design leads to the discovery of p53 transactivation domain I binding compounds that activate the p53 pathway. Chem. Sci. 2020, 12, 3004–3016. [Google Scholar] [CrossRef]
- Kwon, Y.T.; Ciechanover, A. The Ubiquitin Code in the Ubiquitin-Proteasome System and Autophagy. Trends Biochem. Sci. 2017, 42, 873–886. [Google Scholar] [CrossRef]
- Pickart, C.M.; Eddins, M.J. Ubiquitin: Structures, functions, mechanisms. Biochim. Biophys. Acta 2004, 1695, 55–72. [Google Scholar] [CrossRef]
- Dye, B.T.; Schulman, B.A. Structural Mechanisms Underlying Posttranslational Modification by Ubiquitin-Like Proteins. Annu. Rev. Biophys. Biomol. Struct. 2007, 36, 131–150. [Google Scholar] [CrossRef] [PubMed]
- Zheng, N.; Shabek, N. Ubiquitin Ligases: Structure, Function, and Regulation. Annu. Rev. Biochem. 2017, 86, 129–157. [Google Scholar] [CrossRef] [PubMed]
- Cameron, P.; McGachy, A.; Anderson, M.; Paul, A.; Coombs, G.H.; Mottram, J.C.; Alexander, J.; Plevin, R. Inhibition of Lipopolysaccharide-Induced Macrophage IL-12 Production by Leishmania mexicana Amastigotes: The Role of Cysteine Peptidases and the NF-kappaB Signaling Pathway. J. Immunol. 2004, 173, 3297–3304. [Google Scholar] [CrossRef]
- Contreras, I.; Gómez, M.A.; Nguyen, O.; Shio, M.T.; McMaster, R.W.; Olivier, M. Leishmania-Induced Inactivation of the Macrophage Transcription Factor AP-1 Is Mediated by the Parasite Metalloprotease GP63. PLoS Pathog. 2010, 6, e1001148. [Google Scholar] [CrossRef]
- Antoine, J.C.; Jouanne, C.; Lang, T.; Prina, E.; de Chastellier, C.; Frehel, C. Localization of major histocompatibility complex class II molecules in phagolysosomes of murine macrophages infected with Leishmania amazonensis. Infect. Immun. 1991, 59, 764–775. [Google Scholar] [CrossRef]
- Pereira, B.A.; Alves, C.R. Immunological characteristics of experimental murine infection with Leishmania (Leishmania) amazonensis. Veter. Parasitol. 2008, 158, 239–255. [Google Scholar] [CrossRef] [PubMed]
- Morozov, A.V.; Spasskaya, D.S.S.; Karpov, V.D.S.; Karpov, L. The central domain of yeast transcription factor Rpn4 facilitates degradation of reporter protein in human cells. FEBS Lett. 2014, 588, 3713–3719. [Google Scholar] [CrossRef] [PubMed]
- Fortmann, K.T.; Lewis, R.D.; Ngo, K.A.; Fagerlund, R.; Hoffmann, A. A Regulated, Ubiquitin-Independent Degron in IκBα. J. Mol. Biol. 2015, 427, 2748–2756. [Google Scholar] [CrossRef] [PubMed]
- Mashahreh, B.; Armony, S.; Johansson, K.E.; Chappleboim, A.; Friedman, N.; Gardner, R.G.; Hartmann-Petersen, R.; Lindorff-Larsen, K.; Ravid, T. Conserved degronome features governing quality control associated proteolysis. Nat. Commun. 2022, 13, 7588. [Google Scholar] [CrossRef]
- Ahn, D.; Prince, A. Participation of the IL-10RB Related Cytokines, IL-22 and IFN-λ in Defense of the Airway Mucosal Barrier. Front. Cell. Infect. Microbiol. 2020, 10, 300. [Google Scholar] [CrossRef] [PubMed]
- Alves, C.; Corte-Real, S.; Bourguignon, S.C.; Chaves, C.; Saraiva, E. Leishmania amazonensis: Early proteinase activities during promastigote–amastigote differentiation in vitro. Exp. Parasitol. 2005, 109, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Gomes, C.B.; Souza-Silva, F.; dos Santos Charret, K.; Pereira, B.A.S.; Finkelstein, L.C.; Santos-de-Souza, R.; de Castro Côrtes, L.M.; Pereira, M.C.S.; de Oliveira, F.O.R., Jr.; Alves, C.R. Increasing in cysteine proteinase B expression and enzymatic activity during in vitro differentiation of Leishmania (Viannia) braziliensis: First evidence of modulation during morphological transition. Biochimie 2017, 133, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Alves, C.R.; de Souza, R.S.; Charret, K.S.; Côrtes, L.M.C.; de Sá-Silva, M.P.; Barral-Veloso, L.; Oliveira, L.F.G.; da Silva, F.S. Understanding serine proteases implications on Leishmania spp lifecycle. Exp. Parasitol. 2018, 184, 67–81. [Google Scholar] [CrossRef]
- Silva-Almeida, M.; Souza-Silva, F.; Pereira, B.A.S.; Ribeiro-Guimarães, M.L.; Alves, C.R. Overview of the organization of protease genes in the genome of Leishmania spp. Parasit. Vectors 2014, 7, 387. [Google Scholar] [CrossRef]
- Besteiro, S.; Williams, R.A.; Coombs, G.H.; Mottram, J.C. Protein turnover and differentiation in Leishmania. Int. J. Parasitol. 2007, 37, 1063–1075. [Google Scholar] [CrossRef]
- Silverman, J.M.; Clos, J.; Horakova, E.; Wang, A.Y.; Wiesgigl, M.; Kelly, I.; Lynn, M.A.; McMaster, W.R.; Foster, L.J.; Levings, M.K.; et al. Leishmania Exosomes Modulate Innate and Adaptive Immune Responses through Effects on Monocytes and Dendritic Cells. J. Immunol. 2010, 185, 5011–5022. [Google Scholar] [CrossRef]
- Hassani, K.; Shio, M.T.; Martel, C.; Faubert, D.; Olivier, M. Absence of Metalloprotease GP63 Alters the Protein Content of Leishmania Exosomes. PLoS ONE 2014, 9, e95007. [Google Scholar] [CrossRef]
- Clausen, L.; Abildgaard, A.B.; Gersing, S.K.; Stein, A.; Lindorff-Larsen, K.; Hartmann-Petersen, R. Protein stability and degradation in health and disease. Adv. Protein Chem. Struct. Biol. 2019, 114, 61–83. [Google Scholar] [CrossRef]
- Guharoy, M.; Bhowmick, P.; Sallam, M.; Tompa, P. Tripartite degrons confer diversity and specificity on regulated protein degradation in the ubiquitin-proteasome system. Nat. Commun. 2016, 7, 10239. [Google Scholar] [CrossRef]
- Kanarek, N.; London, N.; Schueler-Furman, O.; Ben-Neriah, Y. Ubiquitination and Degradation of the Inhibitors of NF- B. Cold Spring Harb. Perspect. Biol. 2009, 2, a000166. [Google Scholar] [CrossRef] [PubMed]
- Scott, P.; Novais, P.S.F.O. Cutaneous leishmaniasis: Immune responses in protection and pathogenesis. Nat. Rev. Immunol. 2016, 16, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Huang, L.; Mendez, S. A live Leishmania major vaccine containing CpG motifs induces the de novo generation of Th17 cells in C57BL/6 mice. Eur. J. Immunol. 2010, 40, 2517–2527. [Google Scholar] [CrossRef] [PubMed]
- Quirino, G.F.S.; Nascimento, M.S.L.; Davoli-Ferreira, M.; Sacramento, L.A.; Lima, M.H.F.; Almeida, R.P.; Carregaro, V.; Silva, J.S. Interleukin-27 (IL-27) Mediates Susceptibility to Visceral Leishmaniasis by Suppressing the IL-17–Neutrophil Response. Infect. Immun. 2016, 84, 2289–2298. [Google Scholar] [CrossRef] [PubMed]
- Matthews, D.J.; Emson, C.L.; McKenzie, G.J.; Jolin, H.E.; Blackwell, J.M.; McKenzie, A.N. IL-13 Is a Susceptibility Factor for Leishmania major Infection. J. Immunol. 2000, 164, 1458–1462. [Google Scholar] [CrossRef]
- Paul, K.M. Extracellular Vesicles - Role in Diseases, Pathogenesis and Therapy. 2022. Available online: https://www.intechopen.com/books/10796 (accessed on 22 February 2023).
- Dong, G.; Wagner, V.; Minguez-Menendez, A.; Fernandez-Prada, C.; Olivier, M. Extracellular vesicles and leishmaniasis: Current knowledge and promising avenues for future development. Mol. Immunol. 2021, 135, 73–83. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. Signaling to NF-kappaB by Toll-like receptors. Trends Mol. Med. 2007, 13, 460–469. [Google Scholar] [CrossRef]
- Bhardwaj, N.; Rosas, L.E.; Lafuse, W.; Satoskar, A.R. Leishmania inhibits STAT1-mediated IFN-γ signaling in macrophages: Increased tyrosine phosphorylation of dominant negative STAT1β by Leishmania mexicana. Int. J. Parasitol. 2005, 35, 75–82. [Google Scholar] [CrossRef]
- Edwards, C.L.; de Oca, M.M.; Rivera, F.D.L.; Kumar, R.; Ng, S.; Wang, Y.; Amante, F.H.; Kometani, K.; Kurosaki, T.; Sidwell, T.; et al. The Role of BACH2 in T Cells in Experimental Malaria Caused by Plasmodium chabaudi chabaudi AS. Front. Immunol. 2018, 9, 2578. [Google Scholar] [CrossRef]
- Cheeseman, K.; Weitzman, J.B. Host–parasite interactions: An intimate epigenetic relationship. Cell. Microbiol. 2015, 17, 1121–1132. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Organisms | Proteases | Degrons | ||||||||||||||||||||||||||
| Protease Amino Acid Residues Positions Described in Merops | Corresponding Amino Acid Residue Positions (Mus musculus) | |||||||||||||||||||||||||||
| IFN-y | IL-4 | |||||||||||||||||||||||||||
| P4 | P3 | P2 | P1 | P1’ | P2’ | P3’ | P4’ | P4 | P3 | P2 | P1 | P1’ | P2’ | P3’ | P4’ | P4 | P3 | P2 | P1 | P1’ | P2’ | P3’ | P4’ | |||||
| Carica papaya | Papain | P | P | V | R | A | S | G | P | - | G | C | R | C | H | G | P | - | - | F | R | A | F | G | C | |||
| P | P | V | Q | S | R | K | Q | - | P | L | Q | L | R | K | - | - | - | - | - | - | - | - | - | |||||
| P | P | V | A | T | - | P | N | - | P | L | A | L | P | Q | - | - | - | - | - | - | - | - | - | |||||
| P | P | V | A | T | R | P | N | - | P | L | A | L | R | P | - | - | - | - | - | - | - | - | - | |||||
| Homo sapiens | Cathepsin L | - | K | F | R | H | S | - | - | - | R | H | A | K | F | R | - | - | Q | R | K | F | R | A | - | |||
| - | R | F | R | S | H | - | - | - | - | R | L | A | R | F | R | - | F | R | A | F | R | C | - | |||||
| H | K | F | R | H | - | - | - | - | - | F | R | A | H | K | F | - | - | Q | R | K | F | R | A | |||||
| Homo sapiens | Cathepsin B | - | - | F | R | F | F | - | - | - | R | F | F | F | R | F | - | F | R | F | F | F | R | R | F | |||
| - | - | Y | K | F | F | - | - | - | K | F | F | L | K | F | - | Q | K | F | F | R | K | F | - | |||||
| H | L | M | K | - | - | - | - | - | J | L | K | H | Q | L | - | - | - | - | - | - | - | - | - | |||||
| Q | S | V | G | F | A | - | - | - | C | G | F | A | Q | Q | L | - | F | A | Q | S | V | - | - | |||||
| Sus scrofa | Prolyl oligopeptidase | P | P | R | P | Q | P | Q | P | - | P | R | P | Q | P | Q | - | - | P | G | P | N | P | Q | - | |||
| G | Q | G | A | G | G | G | L | G | C | G | C | G | G | G | - | - | Q | G | A | G | G | G | - | |||||
| E. coli | Oligopeptidase B | D | G | R | R | G | Y | I | G | - | G | R | R | G | Q | J | - | D | G | G | R | R | E | I | I | |||
| G | V | G | R | S | S | R | G | - | I | G | R | L | Q | R | - | - | - | - | - | - | - | - | - | |||||
| Organisms | Proteases | Corresponding amino acid residue positions (Mus musculus) | ||||||||||||||||||||||||||
| IL-5 | IL-13 | IL-17 | ||||||||||||||||||||||||||
| P4 | P3 | P2 | P1 | P1’ | P2’ | P3’ | P4’ | P4 | P3 | P2 | P1 | P1’ | P2’ | P3’ | P4 | P3 | P2 | P1 | P1’ | P2’ | P3’ | P4’ | ||||||
| Carica papaya | Papain | - | P | R | R | A | L | G | - | - | P | V | R | A | L | G | - | - | P | M | D | K | Y | G | P | |||
| - | P | R | R | M | R | K | - | - | P | V | Q | A | R | K | - | - | P | M | D | K | R | K | Q | |||||
| - | P | V | P | T | H | K | N | - | P | V | A | A | L | P | - | - | - | M | K | T | G | P | N | |||||
| - | P | V | P | T | R | K | N | - | P | V | A | A | R | P | - | - | P | M | D | K | R | P | N | |||||
| Homo sapiens | Cathepsin L | - | R | H | S | K | F | R | - | - | R | V | L | K | F | R | - | - | H | D | K | F | R | H | - | |||
| - | M | R | R | F | R | L | - | - | R | V | H | R | F | R | - | - | F | R | S | H | R | C | R | |||||
| - | F | R | R | H | K | F | - | - | F | R | H | A | K | F | - | - | F | R | V | H | W | F | - | |||||
| Homo sapiens | Cathepsin B | - | R | R | R | F | R | F | - | - | R | F | F | F | R | F | - | - | R | F | F | F | W | F | - | |||
| P | K | F | F | H | K | N | F | Q | K | F | R | H | G | F | F | P | K | D | F | Y | K | F | - | |||||
| - | L | R | K | H | L | L | - | - | A | M | K | A | L | A | - | - | - | M | D | K | L | I | K | |||||
| - | G | F | A | Q | L | V | - | - | G | F | A | Q | L | V | - | G | R | A | Q | G | V | G | F | |||||
| Sus scrofa | Prolyl oligopeptidase | - | P | R | R | Q | P | Q | - | - | P | R | P | Q | P | Q | - | - | N | R | P | Q | N | Q | - | |||
| - | Q | G | A | G | G | G | - | - | Q | G | A | G | G | G | - | - | Q | G | V | G | W | Q | - | |||||
| E. coli | Oligopeptidase B | - | G | R | R | G | L | J | - | - | G | R | R | G | L | I | - | - | - | R | D | G | Y | I | G | |||
| - | M | G | R | M | L | R | - | - | V | G | R | A | L | R | - | G | D | G | R | I | H | R | - | |||||
| Organisms | Proteases | Degrons | ||||||||||||||||||||||||||
| Protease Amino Acid Residues Positions Described in Merops | Corresponding Amino Acid Residue Positions (Mus musculus) | |||||||||||||||||||||||||||
| Nf-kappa-B | STAT-1 | |||||||||||||||||||||||||||
| P4 | P3 | P2 | P1 | P1’ | P2’ | P3’ | P4’ | P4 | P3 | P2 | P1 | P1’ | P2’ | P3’ | P4’ | P4 | P3 | P2 | P1 | P1’ | P2’ | P3’ | P4’ | |||||
| Carica papaya | Papain | P | P | V | R | A | S | G | P | - | P | G | R | D | G | G | - | - | P | K | R | A | P | G | - | |||
| P | P | V | Q | S | R | K | Q | N | P | M | Z | I | R | K | Q | - | P | V | Q | D | R | K | D | |||||
| P | P | V | A | T | - | P | N | - | P | G | A | D | G | P | - | - | P | M | A | K | K | P | - | |||||
| P | P | V | A | T | R | P | N | P | G | A | D | G | P | - | - | P | M | A | K | K | P | - | ||||||
| Homo sapiens | Cathepsin L | - | K | F | R | H | S | - | - | K | N | R | H | E | K | F | R | - | F | R | H | S | K | D | - | |||
| - | R | F | R | S | H | - | - | F | R | G | F | R | F | R | Y | - | - | H | R | E | R | K | H | |||||
| H | K | F | R | H | - | - | - | - | K | N | R | H | E | K | F | H | K | E | R | K | H | K | W | |||||
| Homo sapiens | Cathepsin B | - | - | F | R | F | F | - | - | - | F | R | F | R | F | G | C | - | R | F | F | F | R | R | - | |||
| - | - | Y | K | F | F | - | - | K | K | N | F | M | E | F | F | - | K | F | F | K | K | F | - | |||||
| H | L | M | K | - | - | - | - | H | L | M | K | K | N | M | K | Q | E | M | K | H | L | W | - | |||||
| Q | S | V | G | F | A | - | - | Q | M | E | G | F | I | Q | - | - | - | F | A | Q | K | K | G | |||||
| Sus scrofa | Prolyl oligopeptidase | P | P | R | P | Q | P | Q | P | P | P | E | P | Q | P | Q | - | - | P | K | P | Q | P | R | - | |||
| G | Q | G | A | G | G | G | L | - | P | G | A | G | G | G | - | G | Q | E | A | G | G | G | W | |||||
| E. coli | Oligopeptidase B | D | G | R | R | G | Y | I | G | - | G | G | R | G | G | I | - | - | - | W | R | G | F | I | G | |||
| G | V | G | R | S | S | R | G | - | P | G | R | D | G | R | - | P | G | R | A | P | R | - | - | |||||
| Organisms | Proteases | Corresponding amino acid residue positions (Mus musculus) | ||||||||||||||||||||||||||
| AP-1 | CREB | BACH2 | ||||||||||||||||||||||||||
| P4 | P3 | P2 | P1 | P1’ | P2’ | P3’ | P4’ | P4 | P3 | P2 | P1 | P1’ | P2’ | P3’ | P4’ | P4 | P3 | P2 | P1 | P1’ | P2’ | P3’ | P4’ | |||||
| Carica papaya | Papain | - | - | - | - | - | - | - | - | - | P | V | R | A | N | G | - | - | P | G | R | P | M | G | - | |||
| - | - | - | - | - | - | - | - | - | P | V | Q | S | R | N | - | - | - | V | Q | D | R | G | Q | |||||
| - | - | - | - | - | - | - | - | - | P | R | K | F | S | P | - | - | P | G | A | P | M | P | - | |||||
| - | P | V | A | S | R | N | - | - | P | G | A | P | R | P | - | |||||||||||||
| Homo sapiens | Cathepsin L | K | C | R | K | R | K | - | - | F | R | H | M | E | N | R | I | F | H | R | M | K | F | R | H | |||
| - | - | - | - | - | - | - | - | - | D | R | V | H | R | E | R | - | F | R | A | H | R | F | - | |||||
| - | K | C | R | K | R | N | - | - | - | F | R | H | H | K | N | F | R | H | H | K | F | R | H | |||||
| Homo sapiens | Cathepsin B | F | E | F | F | F | R | D | D | F | R | F | F | E | R | F | F | F | R | F | F | F | K | F | F | |||
| E | K | F | F | Y | D | D | - | R | K | F | F | E | K | F | - | F | K | F | F | I | K | F | F | |||||
| K | L | M | K | H | L | N | - | K | Q | A | Q | S | N | G | - | F | Q | M | K | I | K | M | K | |||||
| V | G | N | A | Q | N | V | G | - | F | A | Q | S | N | G | V | G | F | A | C | S | E | G | ||||||
| Sus scrofa | Prolyl oligopeptidase | - | - | - | - | - | - | - | - | - | P | R | K | K | P | Z | - | - | P | G | P | P | P | Q | - | |||
| - | - | G | G | G | G | G | L | - | Q | G | A | G | G | G | - | - | P | G | A | G | G | G | - | |||||
| E. coli | Oligopeptidase B | D | E | R | R | G | Y | D | G | - | G | R | R | G | N | N | - | D | G | D | G | G | Y | N | - | |||
| - | - | - | - | - | - | - | - | - | M | G | R | S | N | R | - | G | F | W | R | A | L | R | G | |||||
| Similarity Analysis | ||||||||
|---|---|---|---|---|---|---|---|---|
| Mus musculus Target Proteins | Homo sapiens | Primate (Not Human) | ||||||
| Coverage (%) | E-Value | Identity (%) | Access | Coverage (%) | E-Value | Identity (%) | Access | |
| Nfkappa-B | 100 | 0.0 | 96.2 | NP_001070962.1 | 100 | 0.0 | 91.9 | XP_004050061.1 |
| SAT-1 | 100 | 9 × 10−170 | 83.1 | NP_001304698.1 | 100 | 6 × 10−176 | 85.6 | XP_045388373.1 |
| AP-1 | 100 | 2 × 10−178 | 95.5 | NP_002219.1 | 100 | 0.0 | 98.2 | XP_003921548.1 |
| CREB | 100 | 0.0 | 86.3 | NP_056136.2 | 100 | 0.0 | 86.0 | XP_003796627.1 |
| BACH2 | 99 | 0.0 | 90.1 | NP_001164265.1 | 100 | 0.0 | 89.6 | XP_037858189.1 |
| IFN-Y | 99 | 6 × 10−36 | 41.0 | NP_000610.2 | 99 | 2 × 10−45 | 47.4 | XP_012497489.1 |
| IL-4 | 100 | 4 × 10−23 | 41.3 | CAP72493.1 | 100 | 1 × 10−26 | 42.2 | DP000644.1 |
| IL-5 | 87 | 7 × 10−58 | 71.7 | NP_000870.1 | 94 | 1 × 10−62 | 73.4 | XP_012513070.1 |
| IL-13 | 75 | 1 × 10−30 | 59.8 | NP_002179.2 | 75 | 9 × 10−34 | 59.0 | XP_012506775.1 |
| IL-17 | 96 | 0.0 | 69.5 | NP_703191.2 | 96 | 0.0 | 71.0 | XP_012616045.1 |
| Target | Protease | Protease Motifs | Residues | Alignment of Motifs in the Targets | p-Value Alignment |
|---|---|---|---|---|---|
| Nf-kappaB | Cathepsin B | FRFRFGC | 7 |  | 1.5 × 10−9 |
| STAT-1 | OPB | WRGFIG | 6 | 1.5 × 10−7 | |
| AP-1 | Cathepsin L | KCRKRK | 6 |  | 1.1 × 10−5 |
| CREB | Cathepsin L | DRVHRER | 7 |  | 1.3 × 10−8 |
| BACH2 | Cathepsin L | FRAHRF | 6 |  | 2.7 × 10−8 |
| IFN-y | Cathepsin B | GFAQQL | 6 |  | 2.4 × 10−7 |
| IL-4 | OPB | DGRREIIG | 8 |  | 2.8 × 10−7 |
| IL-5 | OPB | MGRMLR | 6 |  | 1.3 × 10−7 |
| IL-13 | Cathepsin B | AMKALA | 6 |  | 1.0 × 10−8 |
| IL-17 | Cathepsin L | FRSHRCR | 7 |  | 1.1 × 10−9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, A.S.; Aredes-Riguetti, L.M.; Pereira, B.A.S.; Alves, C.R.; Souza-Silva, F. Degron Pathways and Leishmaniasis: Debating Potential Roles of Leishmania spp. Proteases Activity on Guiding Hosts Immune Response and Their Relevance to the Development of Vaccines. Vaccines 2023, 11, 1015. https://doi.org/10.3390/vaccines11061015
Oliveira AS, Aredes-Riguetti LM, Pereira BAS, Alves CR, Souza-Silva F. Degron Pathways and Leishmaniasis: Debating Potential Roles of Leishmania spp. Proteases Activity on Guiding Hosts Immune Response and Their Relevance to the Development of Vaccines. Vaccines. 2023; 11(6):1015. https://doi.org/10.3390/vaccines11061015
Chicago/Turabian StyleOliveira, Adriane Silva, Lara Mata Aredes-Riguetti, Bernardo Acácio Santini Pereira, Carlos Roberto Alves, and Franklin Souza-Silva. 2023. "Degron Pathways and Leishmaniasis: Debating Potential Roles of Leishmania spp. Proteases Activity on Guiding Hosts Immune Response and Their Relevance to the Development of Vaccines" Vaccines 11, no. 6: 1015. https://doi.org/10.3390/vaccines11061015
APA StyleOliveira, A. S., Aredes-Riguetti, L. M., Pereira, B. A. S., Alves, C. R., & Souza-Silva, F. (2023). Degron Pathways and Leishmaniasis: Debating Potential Roles of Leishmania spp. Proteases Activity on Guiding Hosts Immune Response and Their Relevance to the Development of Vaccines. Vaccines, 11(6), 1015. https://doi.org/10.3390/vaccines11061015