
Optimizing the Production of gp145, an HIV-1 Envelope Glycoprotein Vaccine Candidate and Its Encapsulation in Guanosine Microparticles

 , , , and
, , , and
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
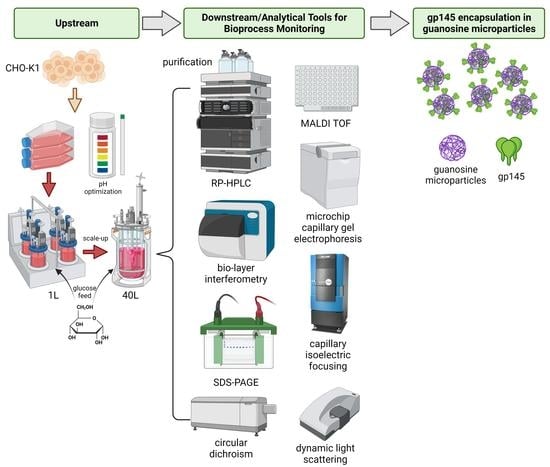
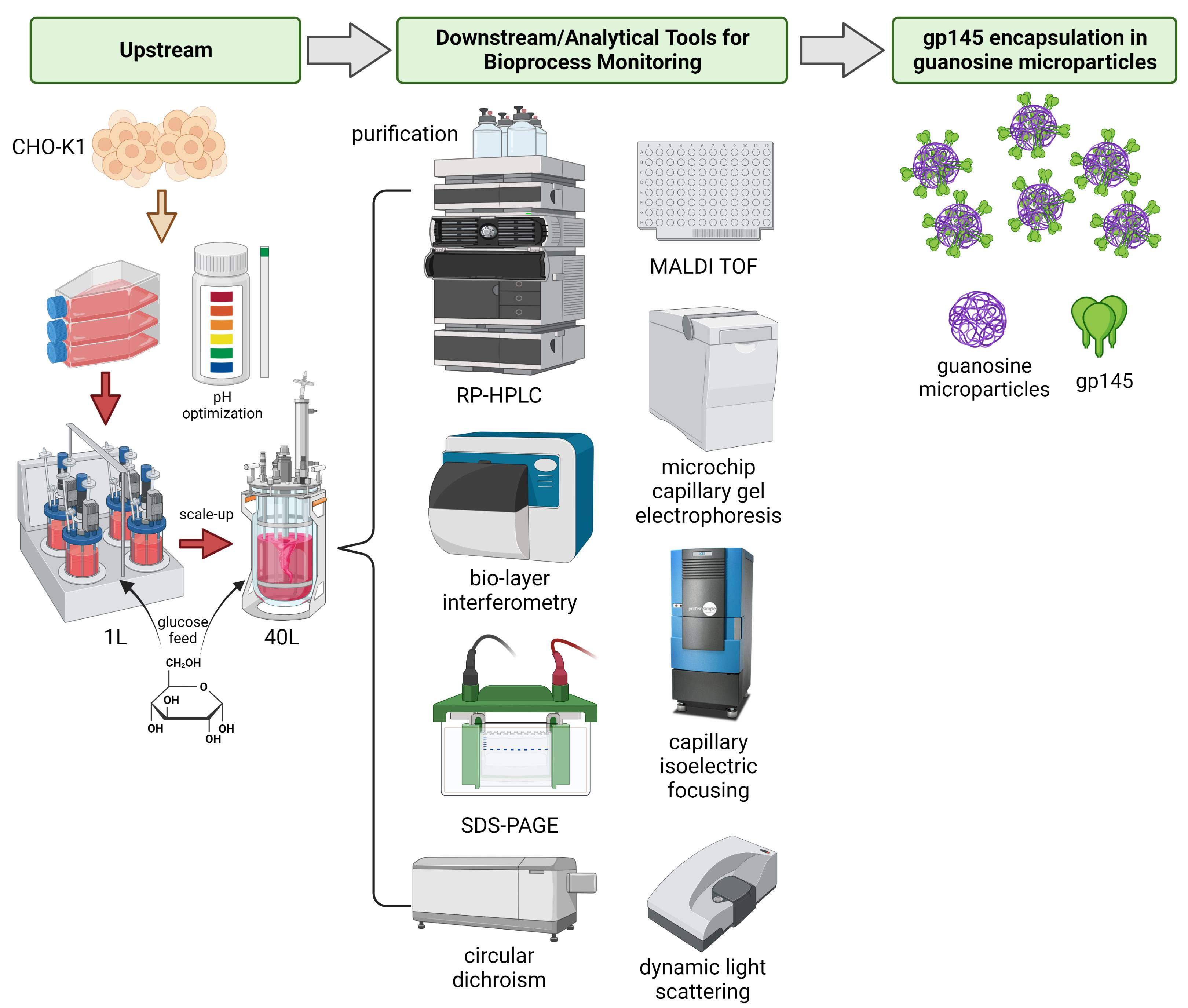
1. Introduction
2. Materials and Methods
2.1. CHO-K1 Cell Line Expressing gp145
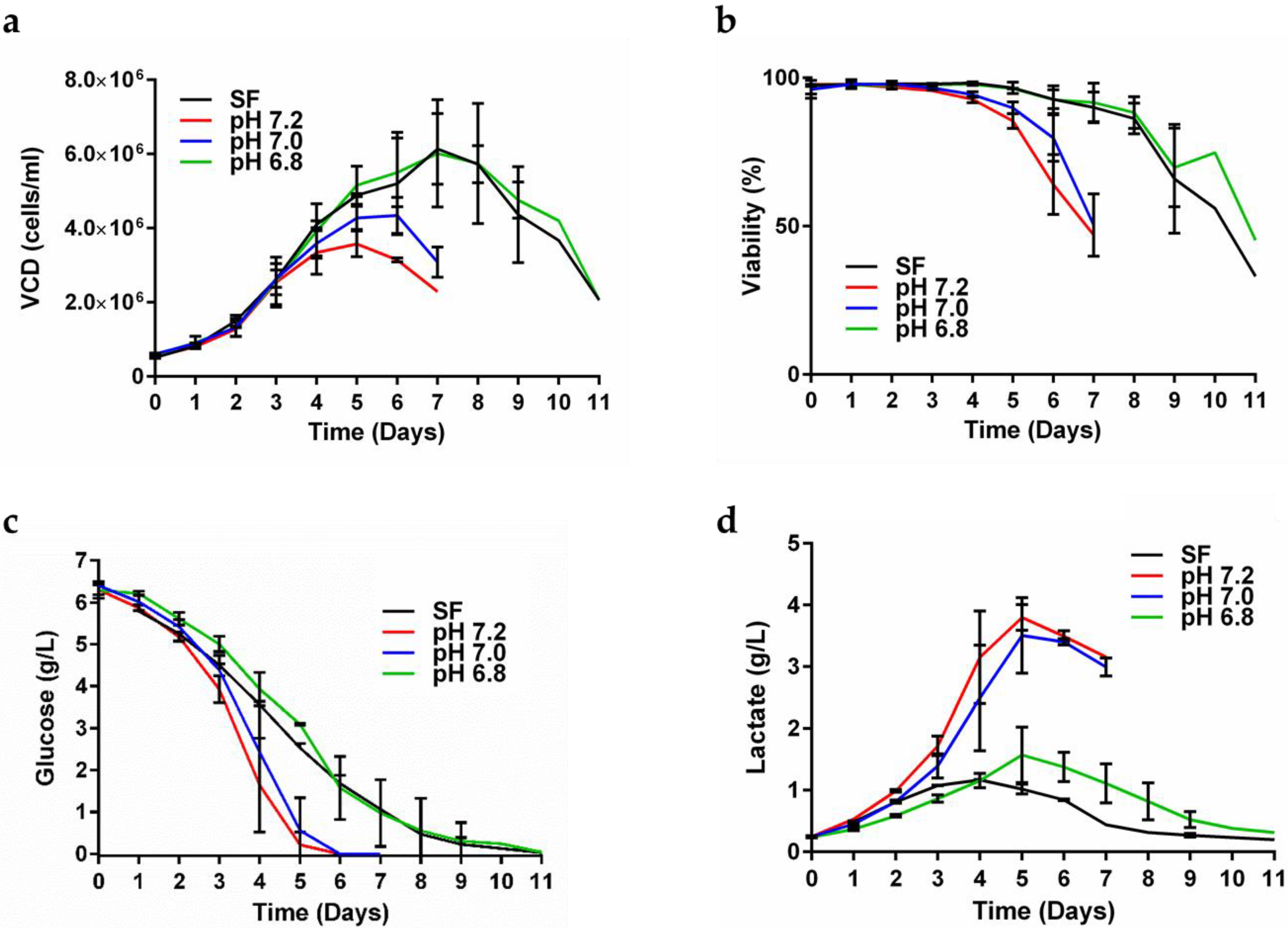
2.2. Upstream and Downstream Strategies to Increase gp145 Titers
2.3. Titer Determination and Hydrophobic Footprint Analysis Using Reversed-Phase High-Performance Liquid Chromatography (RP-HPLC)
2.4. Host Cell Protein (HCP) Determination and Binding Affinity Analysis Using Bio-Layer Interferometry
2.5. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.6. MALDI-TOF Mass Spectrometry
2.7. Mass Analysis
2.8. Glycoform Profiling by Imaged Capillary Isoelectric Focusing (cIEF)
2.9. Size Distribution Analysis by Dynamic Light Scattering (DLS)
2.10. Secondary Structure Analysis by Circular Dichroism
2.11. Synthesis of Supramolecular Hacky Sacks (SHS)
2.12. Fluorescent Labeling of gp145
2.13. Preparation of the gp145-FITC-SHS Complex and VRC01-Alexa Fluor 647 Complex
2.14. Flow Cytometry
2.15. Confocal Imaging
3. Results
3.1. Optimizing the Expression of gp145
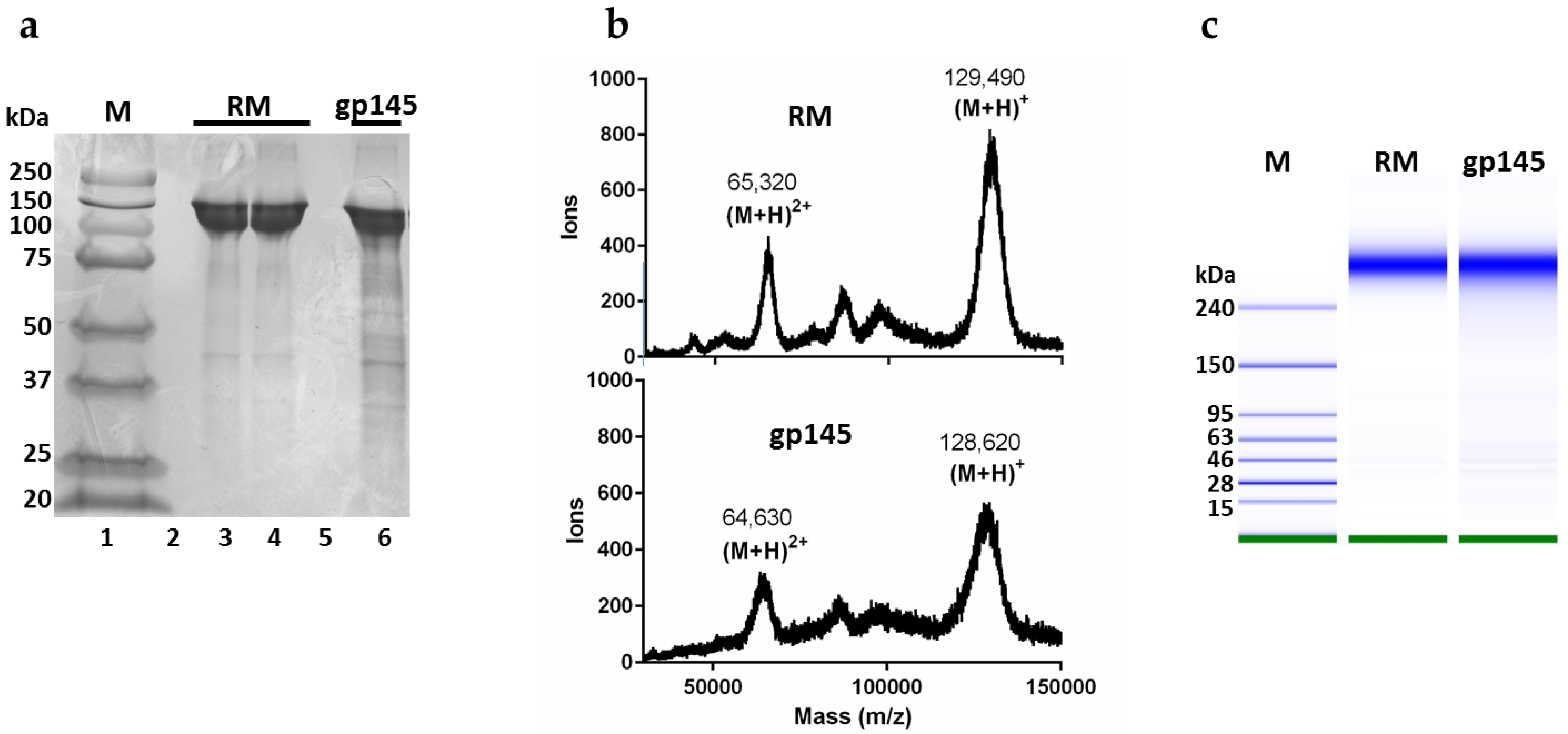
3.2. Analysis of Whole gp145 by SDS PAGE and MALDI-TOF Mass Spectrometry
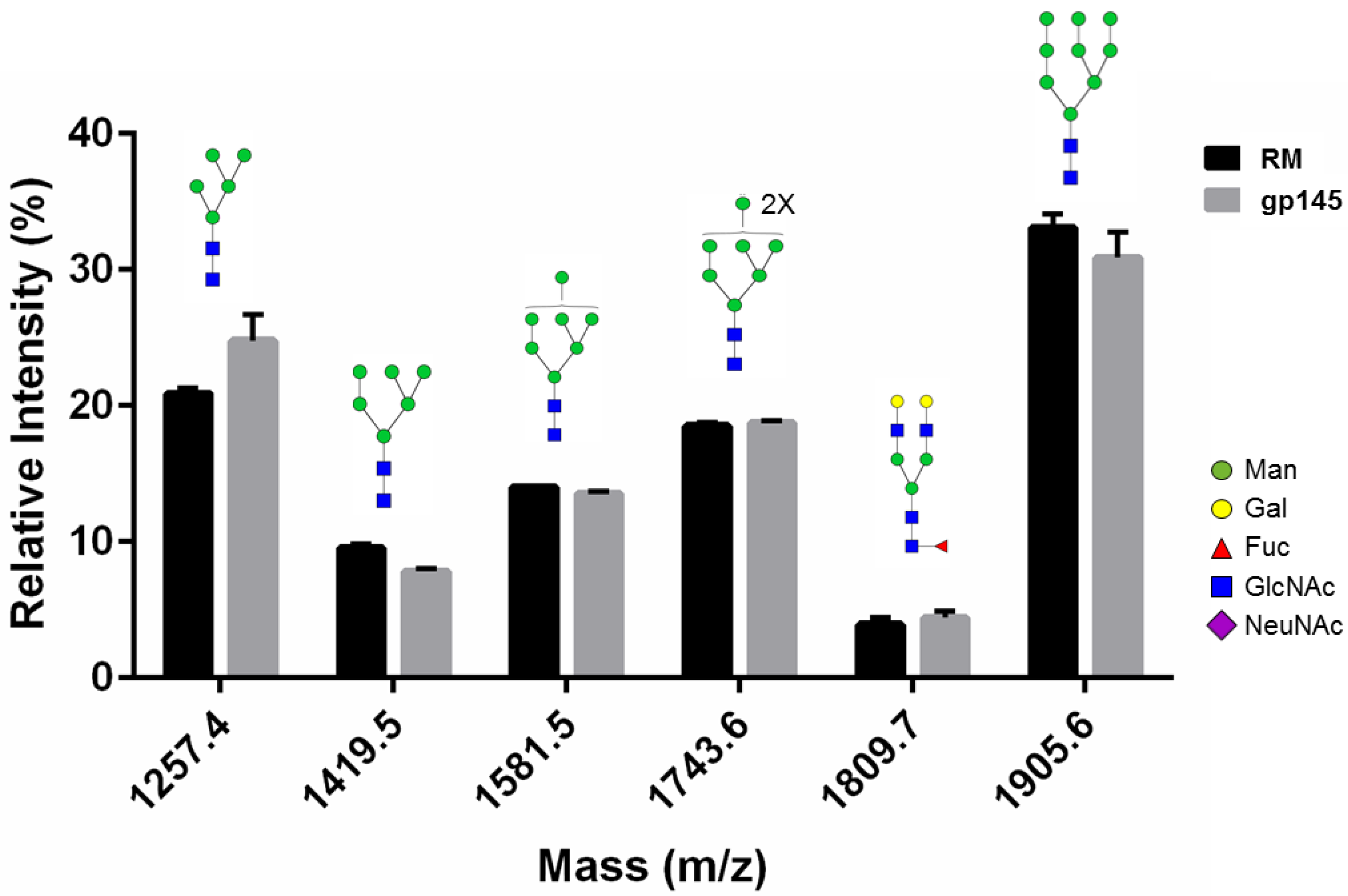
3.3. Glycan Analysis
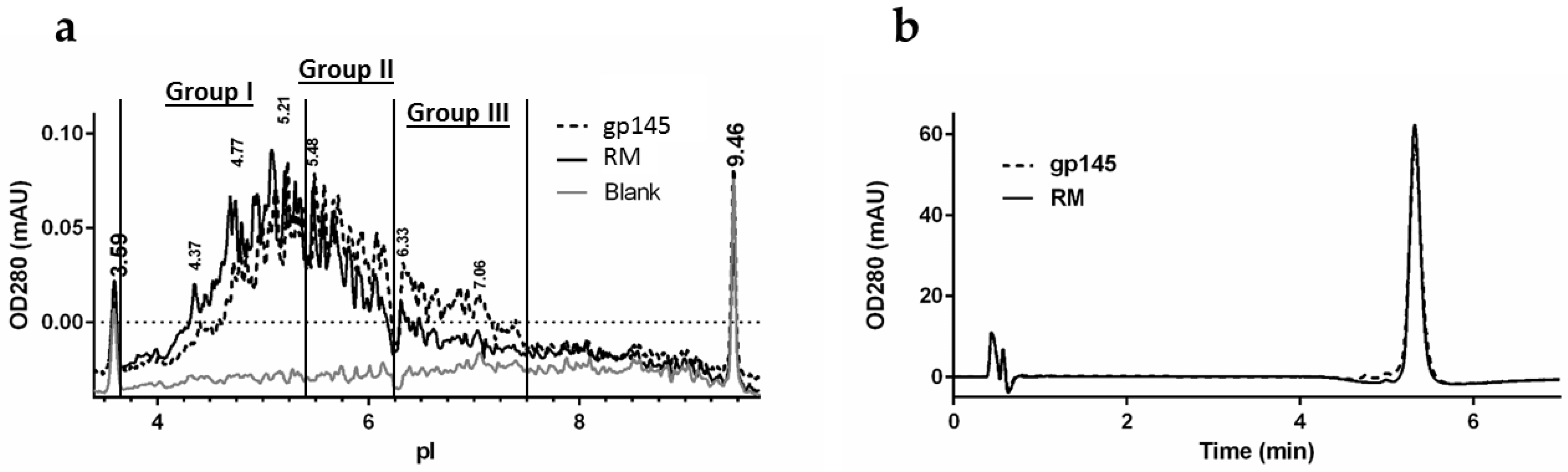
3.4. Biochemical Analysis of gp145 by Capillary Isoelectric Focusing (cIEF) and RP-HPLC
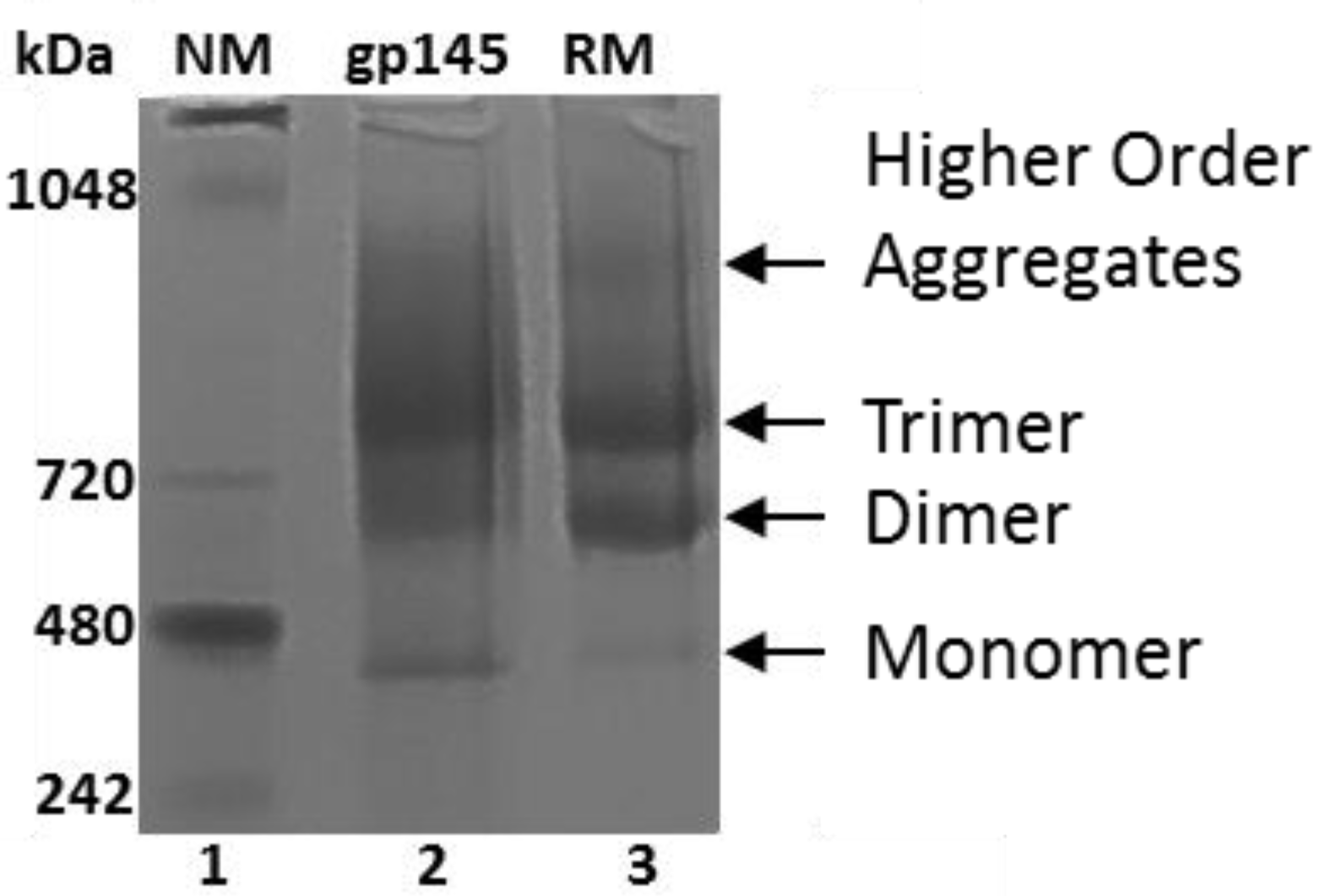
3.5. Oligomeric State of gp145
3.6. Native-Like Properties of gp145
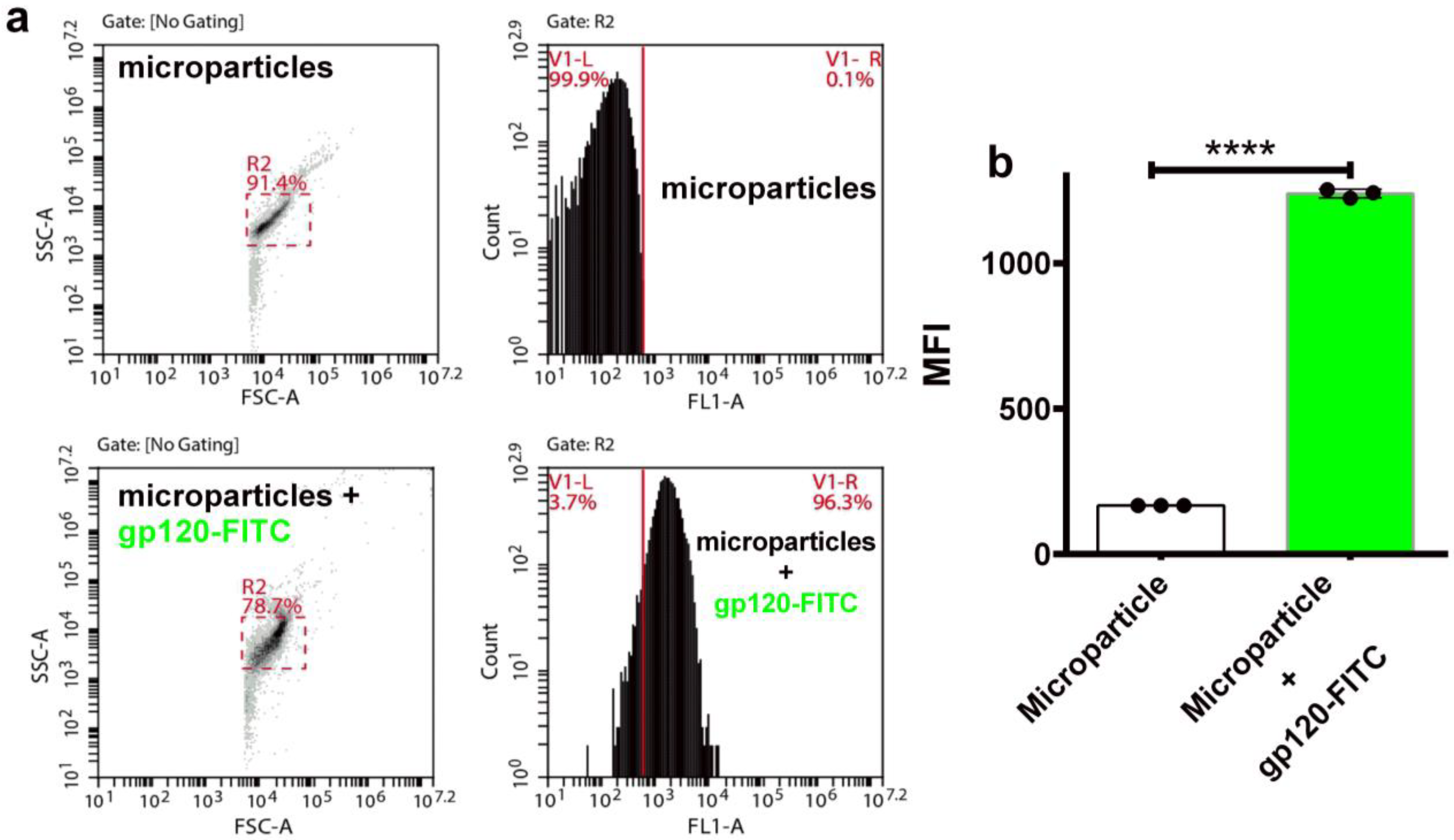
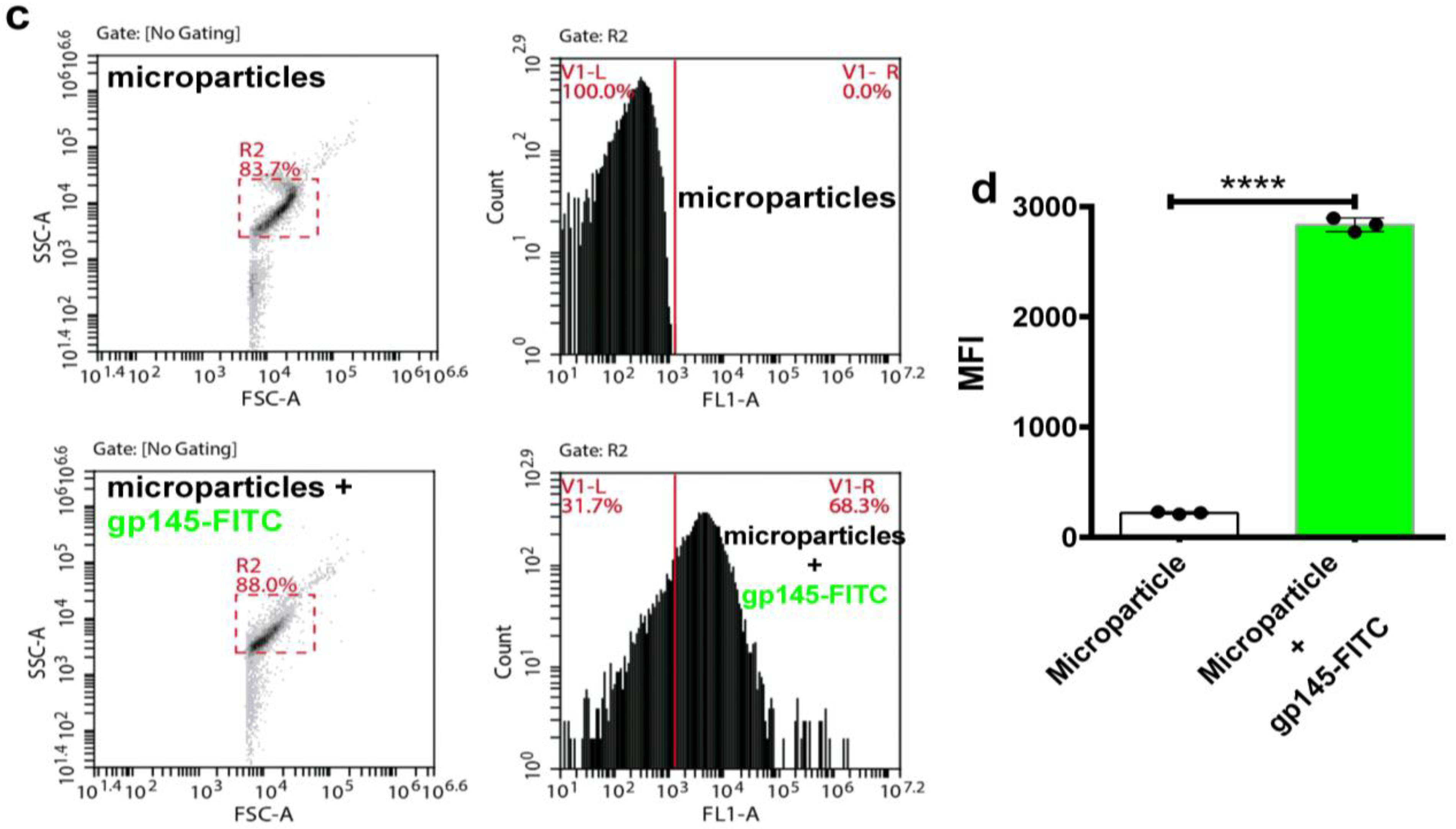
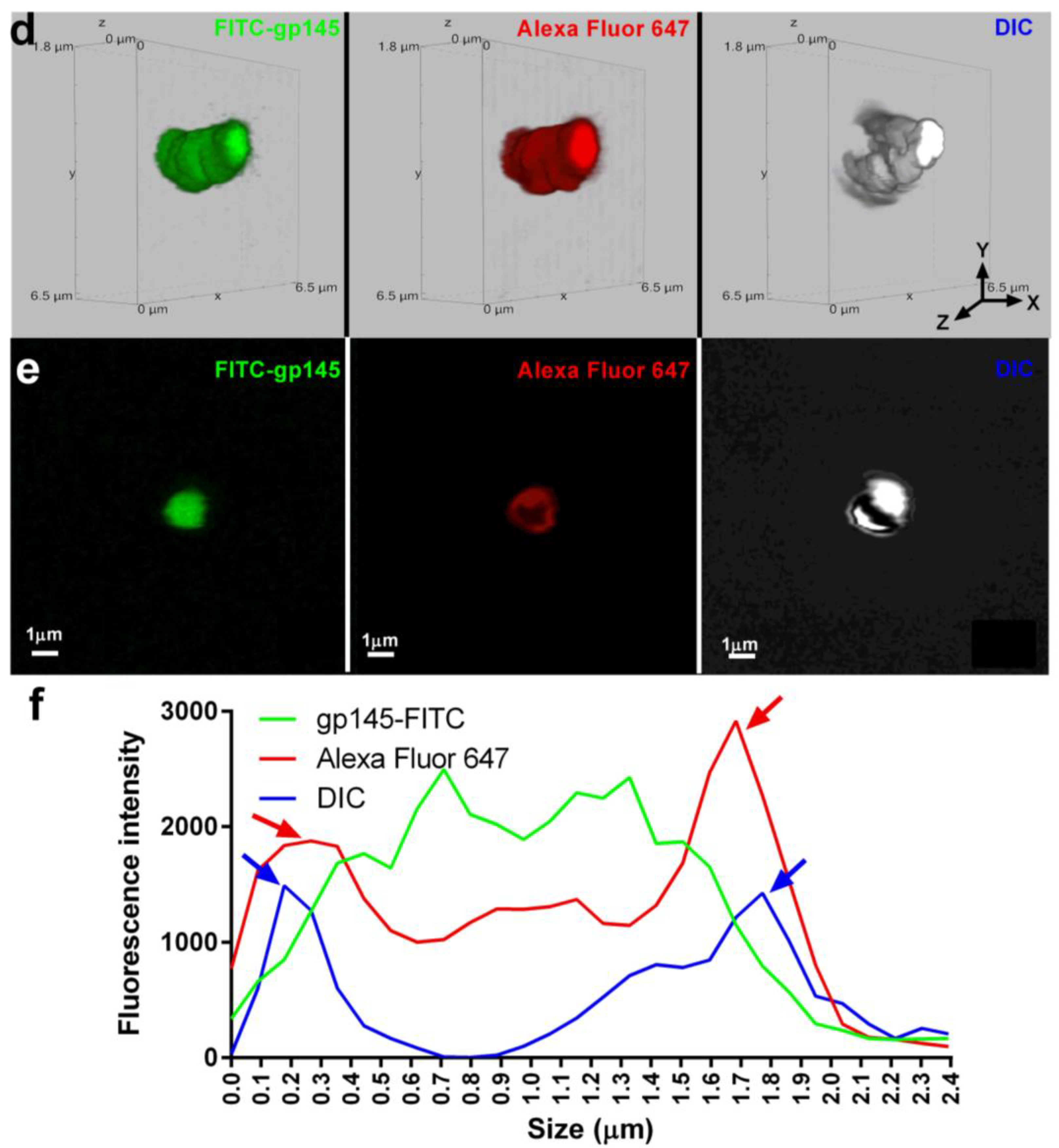
3.7. Encapsulation of gp145 for Delivery
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Code Availability
Acknowledgments
Conflicts of Interest
References
- Kim, J.; Vasan, S.; Kim, J.H.; Ake, J.A. Current approaches to HIV vaccine development: A narrative review. J. Int. AIDS Soc. 2021, 24, e25793. [Google Scholar] [CrossRef] [PubMed]
- Fauci, A.S. An HIV Vaccine Is Essential for Ending the HIV/AIDS Pandemic. JAMA 2017, 318, 1535–1536. [Google Scholar] [CrossRef] [PubMed]
- Rerks-Ngarm, S.; Pitisuttithum, P.; Nitayaphan, S.; Kaewkungwal, J.; Chiu, J.; Paris, R.; Premsri, N.; Namwat, C.; De Souza, M.; Adams, E.; et al. Vaccination with ALVAC and AIDSVAX to Prevent HIV-1 Infection in Thailand. N. Engl. J. Med. 2009, 361, 2209–2220. [Google Scholar] [CrossRef]
- Sanders, R.W.; Moore, J.P. Native-like Env trimers as a platform for HIV-1 vaccine design. Immunol. Rev. 2017, 275, 161–182. [Google Scholar] [CrossRef] [PubMed]
- Kong, R.; Xu, K.; Zhou, T.; Acharya, P.; Lemmin, T.; Liu, K.; Ozorowski, G.; Soto, C.; Taft, J.D.; Bailer, R.T.; et al. Fusion peptide of HIV-1 as a site of vulnerability to neutralizing antibody. Science 2016, 352, 828–833. [Google Scholar] [CrossRef]
- Duan, H.; Chen, X.; Boyington, J.C.; Cheng, C.; Zhang, Y.; Jafari, A.J.; Stephens, T.; Tsybovsky, Y.; Kalyuzhniy, O.; Zhao, P.; et al. Glycan Masking Focuses Immune Responses to the HIV-1 CD4-Binding Site and Enhances Elicitation of VRC01-Class Precursor Antibodies. Immunity 2018, 49, 301–311.e5. [Google Scholar] [CrossRef]
- Prabakaran, P.; Dimitrov, A.S.; Fouts, T.R.; Dimitrov, D.S. Structure and Function of the HIV Envelope Glycoprotein as Entry Mediator, Vaccine Immunogen, and Target for Inhibitors. Adv. Pharmacol. 2007, 55, 33–97. [Google Scholar] [CrossRef]
- McMichael, A.J.; Haynes, B.F. Lessons learned from HIV-1 vaccine trials: New priorities and directions. Nat. Immunol. 2012, 13, 423–427. [Google Scholar] [CrossRef]
- Walker, L.M.; Phogat, S.K.; Chan-Hui, P.-Y.; Wagner, D.; Phung, P.; Goss, J.L.; Wrin, T.; Simek, M.D.; Fling, S.; Mitcham, J.L.; et al. Broad and Potent Neutralizing Antibodies from an African Donor Reveal a New HIV-1 Vaccine Target. Science 2009, 326, 285–289. [Google Scholar] [CrossRef]
- Kovacs, J.M.; Nkolola, J.P.; Peng, H.; Cheung, A.; Perry, J.; Miller, C.A.; Seaman, M.S.; Barouch, D.H.; Chen, B. HIV-1 envelope trimer elicits more potent neutralizing antibody responses than monomeric gp120. Proc. Natl. Acad. Sci. USA 2012, 109, 12111–12116. [Google Scholar] [CrossRef]
- Wieczorek, L.; Krebs, S.J.; Kalyanaraman, V.; Whitney, S.; Tovanabutra, S.; Moscoso, C.G.; Sanders-Buell, E.; Williams, C.; Slike, B.; Molnar, S.; et al. Comparable Antigenicity and Immunogenicity of Oligomeric Forms of a Novel, Acute HIV-1 Subtype C gp145 Envelope for Use in Preclinical and Clinical Vaccine Research. J. Virol. 2015, 89, 7478–7493. [Google Scholar] [CrossRef]
- Sharma, S.K.; de Val, N.; Bale, S.; Guenaga, J.; Tran, K.; Feng, Y.; Dubrovskaya, V.; Ward, A.B.; Wyatt, R.T. Cleavage-Independent HIV-1 Env Trimers Engineered as Soluble Native Spike Mimetics for Vaccine Design. Cell Rep. 2015, 11, 539–550. [Google Scholar] [CrossRef]
- Aldon, Y.; McKay, P.F.; Allen, J.; Ozorowski, G.; Lévai, R.F.; Tolazzi, M.; Rogers, P.; He, L.; de Val, N.; Fábián, K.; et al. Rational Design of DNA-Expressed Stabilized Native-Like HIV-1 Envelope Trimers. Cell Rep. 2018, 24, 3324–3338.e5. [Google Scholar] [CrossRef]
- Binley, J.M.; Sanders, R.W.; Clas, B.; Schuelke, N.; Master, A.; Guo, Y.; Kajumo, F.; Anselma, D.J.; Maddon, P.J.; Olson, W.C.; et al. A Recombinant Human Immunodeficiency Virus Type 1 Envelope Glycoprotein Complex Stabilized by an Intermolecular Disulfide Bond between the gp120 and gp41 Subunits Is an Antigenic Mimic of the Trimeric Virion-Associated Structure. J. Virol. 2000, 74, 627–643. [Google Scholar] [CrossRef]
- Sanders, R.W.; Derking, R.; Cupo, A.; Julien, J.-P.; Yasmeen, A.; de Val, N.; Kim, H.J.; Blattner, C.; de la Peña, A.T.; Korzun, J.; et al. A Next-Generation Cleaved, Soluble HIV-1 Env Trimer, BG505 SOSIP.664 gp140, Expresses Multiple Epitopes for Broadly Neutralizing but Not Non-Neutralizing Antibodies. PLoS Pathog. 2013, 9, e1003618. [Google Scholar] [CrossRef]
- Sanders, R.W.; Venturi, M.; Schiffner, L.; Kalyanaraman, R.; Katinger, H.; Lloyd, K.O.; Kwong, P.D.; Moore, J.P. The Mannose-Dependent Epitope for Neutralizing Antibody 2G12 on Human Immunodeficiency Virus Type 1 Glycoprotein gp120. J. Virol. 2002, 76, 7293–7305. [Google Scholar] [CrossRef]
- O’rourke, S.M.; Byrne, G.; Tatsuno, G.; Wright, M.; Yu, B.; Mesa, K.A.; Doran, R.C.; Alexander, D.; Berman, P.W. Robotic selection for the rapid development of stable CHO cell lines for HIV vaccine production. PLoS ONE 2018, 13, e0197656. [Google Scholar] [CrossRef]
- Hossler, P.; Khattak, S.F.; Li, Z.J. Optimal and consistent protein glycosylation in mammalian cell culture. Glycobiology 2009, 19, 936–949. [Google Scholar] [CrossRef]
- Li, Y.; Luo, L.; Thomas, D.Y.; Kang, O. Control of Expression, Glycosylation, and Secretion of HIV-1 gp120 by Homologous and Heterologous Signal Sequences. Virology 1994, 204, 266–278. [Google Scholar] [CrossRef]
- Bale, S.; Martiné, A.; Wilson, R.; Behrens, A.-J.; Le Fourn, V.; De Val, N.; Sharma, S.K.; Tran, K.; Torres, J.L.; Girod, P.-A.; et al. Cleavage-Independent HIV-1 Trimers from CHO Cell Lines Elicit Robust Autologous Tier 2 Neutralizing Antibodies. Front. Immunol. 2018, 9, 1116. [Google Scholar] [CrossRef]
- Pati, R.; Shevtsov, M.; Sonawane, A. Nanoparticle Vaccines Against Infectious Diseases. Front. Immunol. 2018, 9, 2224. [Google Scholar] [CrossRef] [PubMed]
- Luthra, A.; Cheema, S.; Whitney, S.; Bakker, W.A.; Sandalon, Z.; Richardson, J.; Yallop, C.; Havenga, M. Stable, high yield expression of gp145 Env glycoprotein from HIV-1 in mammalian cells. Biologicals 2021, 73, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Dey, A.K.; Cupo, A.; Ozorowski, G.; Sharma, V.K.; Behrens, A.; Go, E.P.; Ketas, T.J.; Yasmeen, A.; Klasse, P.J.; Sayeed, E.; et al. cGMP production and analysis of BG505 SOSIP.664, an extensively glycosylated, trimeric HIV-1 envelope glycoprotein vaccine candidate. Biotechnol. Bioeng. 2017, 115, 885–899. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, L.S.; Smedley, J.G.; Bubna, N.; Hussain, A.; Harper, R.; Mostafa, S. Development of a platform-based approach for the clinical production of HIV gp120 envelope glycoprotein vaccine candidates. Vaccine 2021, 39, 3852–3861. [Google Scholar] [CrossRef] [PubMed]
- Lasalde-Dominicci, J.A.; Baerga-Ortiz, A.; Akamine, P.; González-Feliciano, J.A.; Capó-Vélez, C.; Delgado-Vélez, M. A Center for the Rapid Analysis of Clinical-Grade Biologics: The Biophysical Characterization of HIV-1 Env Protein. Proceedings 2020, 50, 130. [Google Scholar] [CrossRef]
- González-Feliciano, J.A.; Capó-Vélez, C.M.; Akamine, P.; Delgado-Vélez, M.; Almodóvar, R.; Rivera, J.; Pino, I.; Morell, G.; Eichinger, D.; Rivera, J.H.; et al. A reversed phase HPLC method for the quantification of HIV gp145 glycoprotein levels from cell culture supernatants. J. Chromatogr. B 2021, 1167, 122562. [Google Scholar] [CrossRef]
- González-Feliciano, J.A.; Akamine, P.; Capó-Vélez, C.M.; Delgado-Vélez, M.; Dussupt, V.; Krebs, S.J.; Wojna, V.; Polonis, V.R.; Baerga-Ortiz, A.; Lasalde-Dominicci, J.A. A recombinant gp145 Env glycoprotein from HIV-1 expressed in two different cell lines: Effects on glycosylation and antigenicity. PLoS ONE 2020, 15, e0231679. [Google Scholar] [CrossRef]
- Strohalm, M.; Hassman, M.; Košata, B.; Kodíček, M. mMass data miner: An open source alternative for mass spectrometric data analysis. Rapid Commun. Mass Spectrom. 2008, 22, 905–908. [Google Scholar] [CrossRef]
- Küster, B.; Hunter, A.P.; Wheeler, S.F.; Dwek, R.A.; Harvey, D.J. Structural determination ofN-linked carbohydrates by matrix-assisted laser desorption/ionization-mass spectrometry following enzymatic release within sodium dodecyl sulphate-polyacrylamide electrophoresis gels: Application to species-specific glycosylation of α1-acid glycoprotein. Electrophoresis 1998, 19, 1950–1959. [Google Scholar] [CrossRef]
- Negrón, L.M.; Meléndez-Contés, Y.; Rivera, J.M. Patchy Supramolecules as Versatile Tools to Probe Hydrophobicity in Nanoglobular Systems. J. Am. Chem. Soc. 2013, 135, 3815–3817. [Google Scholar] [CrossRef]
- Rivera, J.M.; Negron, L.M. Supramolecular Hacky Sacks (SHS), Method of Synthesis and Applications Thereof. U.S. Patent No. 10,106,572, 23 October 2018. [Google Scholar]
- Jeyarajah, S.; Parker, C.E.; Sumner, M.T.; Tomer, K.B. Matrix-assisted laser desorption ionization/mass spectrometry mapping of human immunodeficiency virus-gp120 epitopes recognized by a limited polyclonal antibody. J. Am. Soc. Mass Spectrom. 1998, 9, 157–165. [Google Scholar] [CrossRef]
- Engel, N.; Weiss, V.U.; Wenz, C.; Rüfer, A.; Kratzmeier, M.; Glück, S.; Marchetti-Deschmann, M.; Allmaier, G. Challenges of glycoprotein analysis by microchip capillary gel electrophoresis. Electrophoresis 2015, 36, 1754–1758. [Google Scholar] [CrossRef]
- Doores, K.J.; Bonomelli, C.; Harvey, D.J.; Vasiljevic, S.; Dwek, R.A.; Burton, D.R.; Crispin, M.; Scanlan, C.N. Envelope glycans of immunodeficiency virions are almost entirely oligomannose antigens. Proc. Natl. Acad. Sci. USA 2010, 107, 13800–13805. [Google Scholar] [CrossRef]
- Go, E.P.; Ding, H.; Zhang, S.; Ringe, R.P.; Nicely, N.; Hua, D.; Steinbock, R.T.; Golabek, M.; Alin, J.; Alam, S.M.; et al. Glycosylation Benchmark Profile for HIV-1 Envelope Glycoprotein Production Based on Eleven Env Trimers. J. Virol. 2017, 91, e02428-16. [Google Scholar] [CrossRef]
- Yu, B.; Morales, J.F.; O’Rourke, S.M.; Tatsuno, G.P.; Berman, P.W. Glycoform and Net Charge Heterogeneity in gp120 Immunogens Used in HIV Vaccine Trials. PLoS ONE 2012, 7, e43903. [Google Scholar] [CrossRef]
- Nobbmann, U. Polydispersity—What Does It Mean for DLS and Chromatography?—Materials Talks. 2017. Available online: https://www.materials-talks.com/polydispersity-what-does-it-mean-for-dls-and-chromatography/ (accessed on 25 July 2022).
- Priya, R.S.; Veena, M.; Kalisz, I.; Whitney, S.; Priyanka, D.; LaBranche, C.C.; Teja, M.S.; Montefiori, D.C.; Pal, R.; Mahalingam, S.; et al. Antigenicity and Immunogenicity of a Trimeric Envelope Protein from an Indian Clade C HIV-1 Isolate. J. Biol. Chem. 2015, 290, 9195–9208. [Google Scholar] [CrossRef]
- Santos, S.; Ramírez, M.; Miranda, E.; Reyes, N.; Martinez, O.; Acosta-Santiago, M.; Rivera, J.M.; Otero, M. Enhancement of Immune Responses by Guanosine-Based Particles in DNA Plasmid Formulations against Infectious Diseases. J. Immunol. Res. 2019, 2019, 3409371. [Google Scholar] [CrossRef]
- Wu, X.; Yang, Z.-Y.; Li, Y.; Hogerkorp, C.-M.; Schief, W.R.; Seaman, M.S.; Zhou, T.; Schmidt, S.D.; Wu, L.; Xu, L.; et al. Rational Design of Envelope Identifies Broadly Neutralizing Human Monoclonal Antibodies to HIV-1. Science 2010, 329, 856–861. [Google Scholar] [CrossRef]
- Zhou, T.; Georgiev, I.; Wu, X.; Yang, Z.-Y.; Dai, K.; Finzi, A.; Kwon, Y.D.; Scheid, J.F.; Shi, W.; Xu, L.; et al. Structural Basis for Broad and Potent Neutralization of HIV-1 by Antibody VRC01. Science 2010, 329, 811–817. [Google Scholar] [CrossRef]
- Trummer, E.; Fauland, K.; Seidinger, S.; Schriebl, K.; Lattenmayer, C.; Kunert, R.; Vorauer-Uhl, K.; Weik, R.; Borth, N.; Katinger, H.; et al. Process parameter shifting: Part I. Effect of DOT, pH, and temperature on the performance of Epo-Fc expressing CHO cells cultivated in controlled batch bioreactors. Biotechnol. Bioeng. 2006, 94, 1033–1044. [Google Scholar] [CrossRef]
- Xu, S.; Chen, H. High-density mammalian cell cultures in stirred-tank bioreactor without external pH control. J. Biotechnol. 2016, 231, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Pabst, M.; Chang, M.; Stadlmann, J.; Altmann, F. Glycan profiles of the 27 N-glycosylation sites of the HIV envelope protein CN54gp140. Biol. Chem. 2012, 393, 719–730. [Google Scholar] [CrossRef] [PubMed]
- Go, E.P.; Irungu, J.; Zhang, Y.; Dalpathado, D.S.; Liao, H.-X.; Sutherland, L.L.; Alam, S.M.; Haynes, B.F.; Desaire, H. Glycosylation Site-Specific Analysis of HIV Envelope Proteins (JR-FL and CON-S) Reveals Major Differences in Glycosylation Site Occupancy, Glycoform Profiles, and Antigenic Epitopes’ Accessibility. J. Proteome Res. 2008, 7, 1660–1674. [Google Scholar] [CrossRef] [PubMed]
- Rogers, R.S.; Nightlinger, N.S.; Livingston, B.; Campbell, P.; Bailey, R.; Balland, A. Development of a quantitative mass spectrometry multi-attribute method for characterization, quality control testing and disposition of biologics. Mabs 2015, 7, 881–890. [Google Scholar] [CrossRef]
- Rogstad, S.; Yan, H.; Wang, X.; Powers, D.; Brorson, K.; Damdinsuren, B.; Lee, S.L. Multi-Attribute Method for Quality Control of Therapeutic Proteins. Anal. Chem. 2019, 91, 14170–14177. [Google Scholar] [CrossRef]
- Brinkkemper, M.; Sliepen, K. Nanoparticle Vaccines for Inducing HIV-1 Neutralizing Antibodies. Vaccines 2019, 7, 76. [Google Scholar] [CrossRef]
- Nguyen, B.; Tolia, N.H. Protein-based antigen presentation platforms for nanoparticle vaccines. NPJ Vaccines 2021, 6, 70. [Google Scholar] [CrossRef]
- Moodie, Z.; Dintwe, O.; Sawant, S.; Grove, D.; Huang, Y.; Janes, H.; Heptinstall, J.; Omar, F.L.; Cohen, K.; De Rosa, S.C.; et al. Analysis of the HIV Vaccine Trials Network 702 Phase 2b–3 HIV-1 Vaccine Trial in South Africa Assessing RV144 Antibody and T-Cell Correlates of HIV-1 Acquisition Risk. J. Infect. Dis. 2022, 226, 246–257. [Google Scholar] [CrossRef]
- National Institutes of Health. HIV Vaccine Candidate Does Not Sufficiently Protect Women Against HIV Infection|NIH: National Institute of Allergy and Infectious Diseases n.d. Available online: https://www.niaid.nih.gov/news-events/hiv-vaccine-candidate-does-not-sufficiently-protect-women-against-hiv-infection (accessed on 19 January 2023).
- National Institutes of Health. Experimental HIV Vaccine Regimen Safe but Ineffective, Study Finds. National Institutes of Health (NIH). 2023. Available online: https://www.nih.gov/news-events/news-releases/experimental-hiv-vaccine-regimen-safe-ineffective-study-finds (accessed on 19 January 2023).












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akamine, P.; González-Feliciano, J.A.; Almodóvar, R.; Morell, G.; Rivera, J.; Capó-Vélez, C.M.; Delgado-Vélez, M.; Prieto-Costas, L.; Madera, B.; Eichinger, D.; et al. Optimizing the Production of gp145, an HIV-1 Envelope Glycoprotein Vaccine Candidate and Its Encapsulation in Guanosine Microparticles. Vaccines 2023, 11, 975. https://doi.org/10.3390/vaccines11050975
Akamine P, González-Feliciano JA, Almodóvar R, Morell G, Rivera J, Capó-Vélez CM, Delgado-Vélez M, Prieto-Costas L, Madera B, Eichinger D, et al. Optimizing the Production of gp145, an HIV-1 Envelope Glycoprotein Vaccine Candidate and Its Encapsulation in Guanosine Microparticles. Vaccines. 2023; 11(5):975. https://doi.org/10.3390/vaccines11050975
Chicago/Turabian StyleAkamine, Pearl, José A. González-Feliciano, Ruth Almodóvar, Gloriner Morell, Javier Rivera, Coral M. Capó-Vélez, Manuel Delgado-Vélez, Luis Prieto-Costas, Bismark Madera, Daniel Eichinger, and et al. 2023. "Optimizing the Production of gp145, an HIV-1 Envelope Glycoprotein Vaccine Candidate and Its Encapsulation in Guanosine Microparticles" Vaccines 11, no. 5: 975. https://doi.org/10.3390/vaccines11050975
APA StyleAkamine, P., González-Feliciano, J. A., Almodóvar, R., Morell, G., Rivera, J., Capó-Vélez, C. M., Delgado-Vélez, M., Prieto-Costas, L., Madera, B., Eichinger, D., Pino, I., Rivera, J. H., Ortiz-Ubarri, J., Rivera, J. M., Baerga-Ortiz, A., & Lasalde-Dominicci, J. A. (2023). Optimizing the Production of gp145, an HIV-1 Envelope Glycoprotein Vaccine Candidate and Its Encapsulation in Guanosine Microparticles. Vaccines, 11(5), 975. https://doi.org/10.3390/vaccines11050975