
Targeting Membrane Trafficking as a Strategy for Cancer Treatment




Abstract
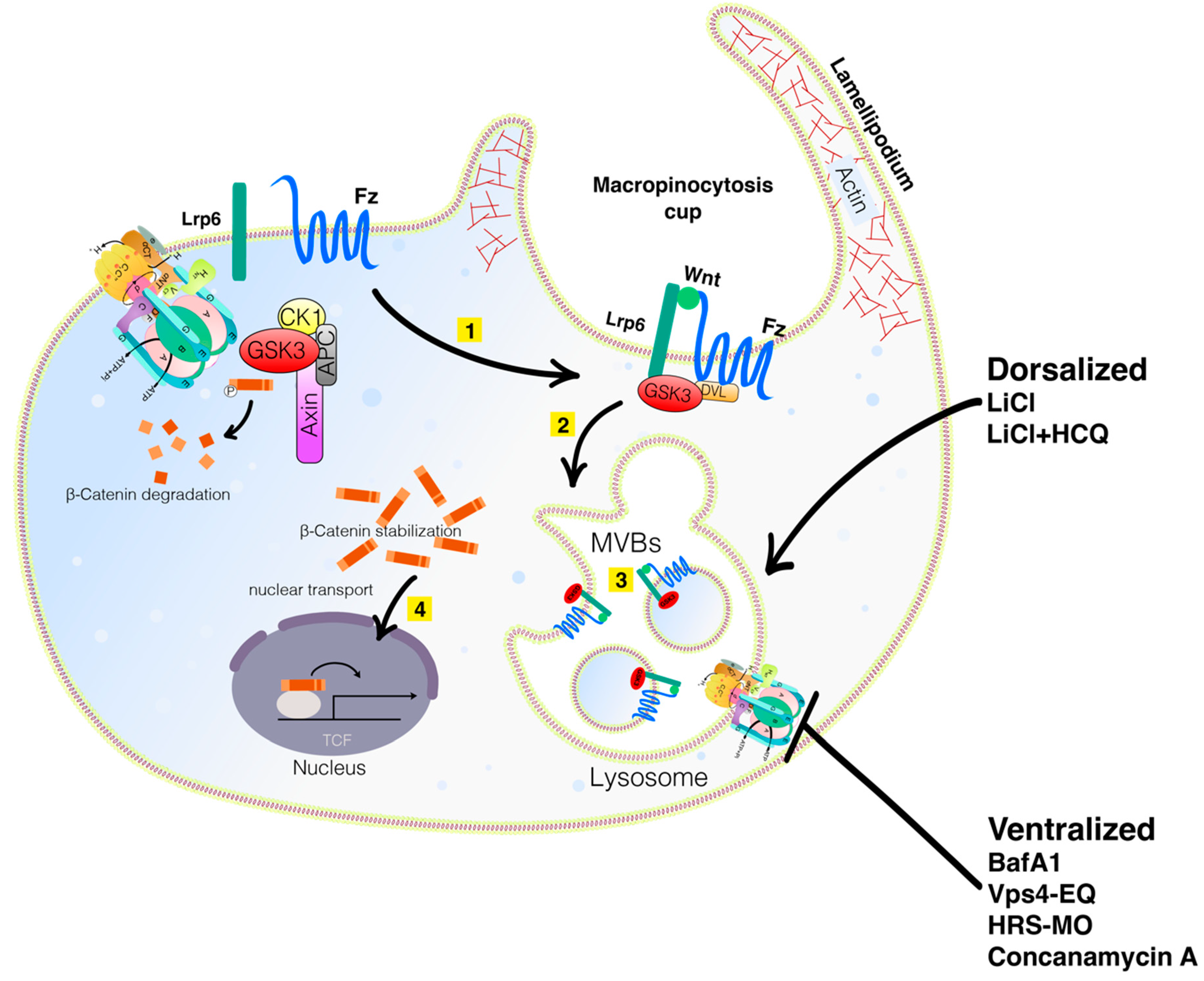
:1. Membrane Trafficking, Lysosomes, V-ATPase, and Macropinocytosis in the Wnt Pathway
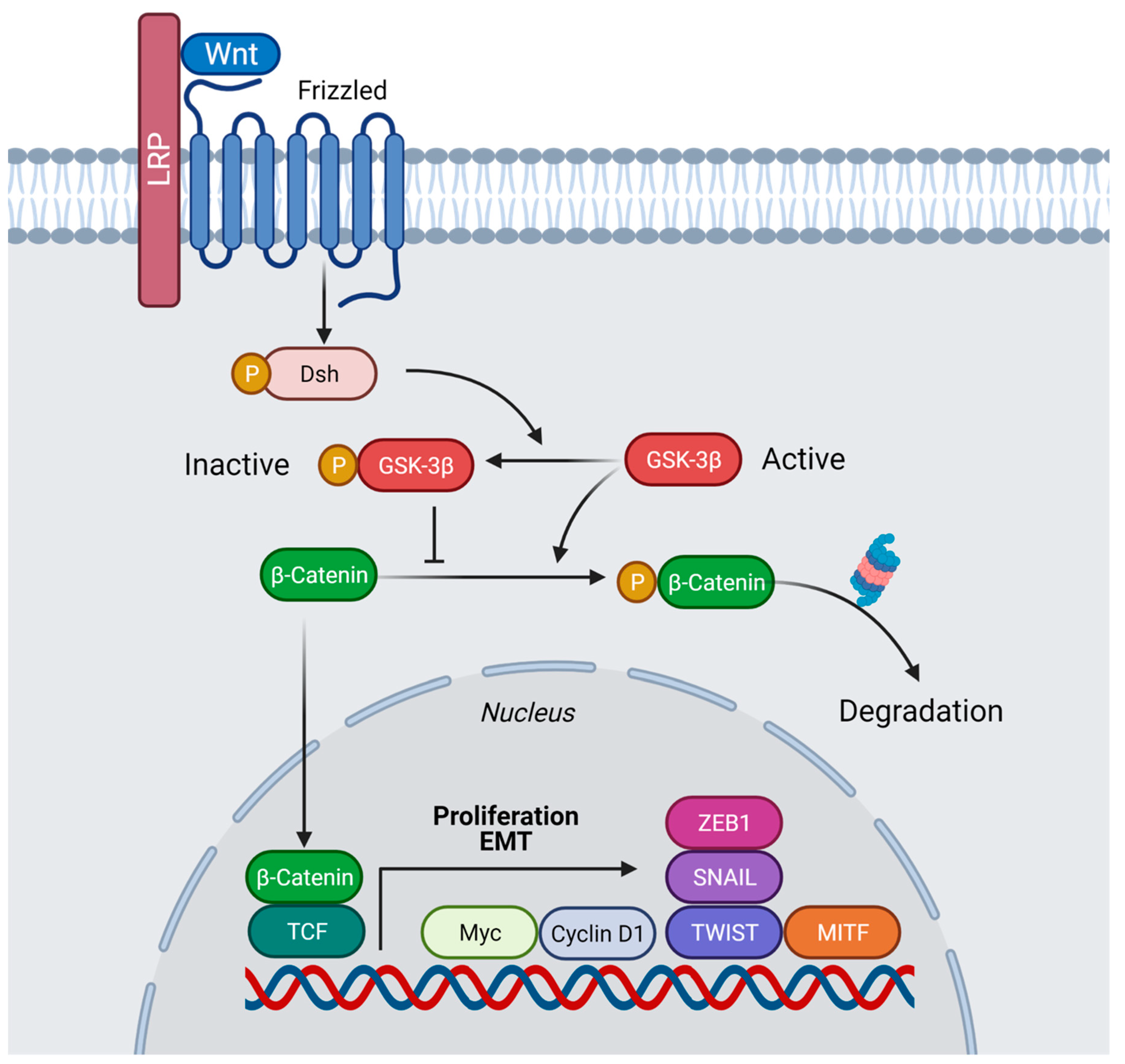
2. Wnt Signaling Triggers Macropinocytosis
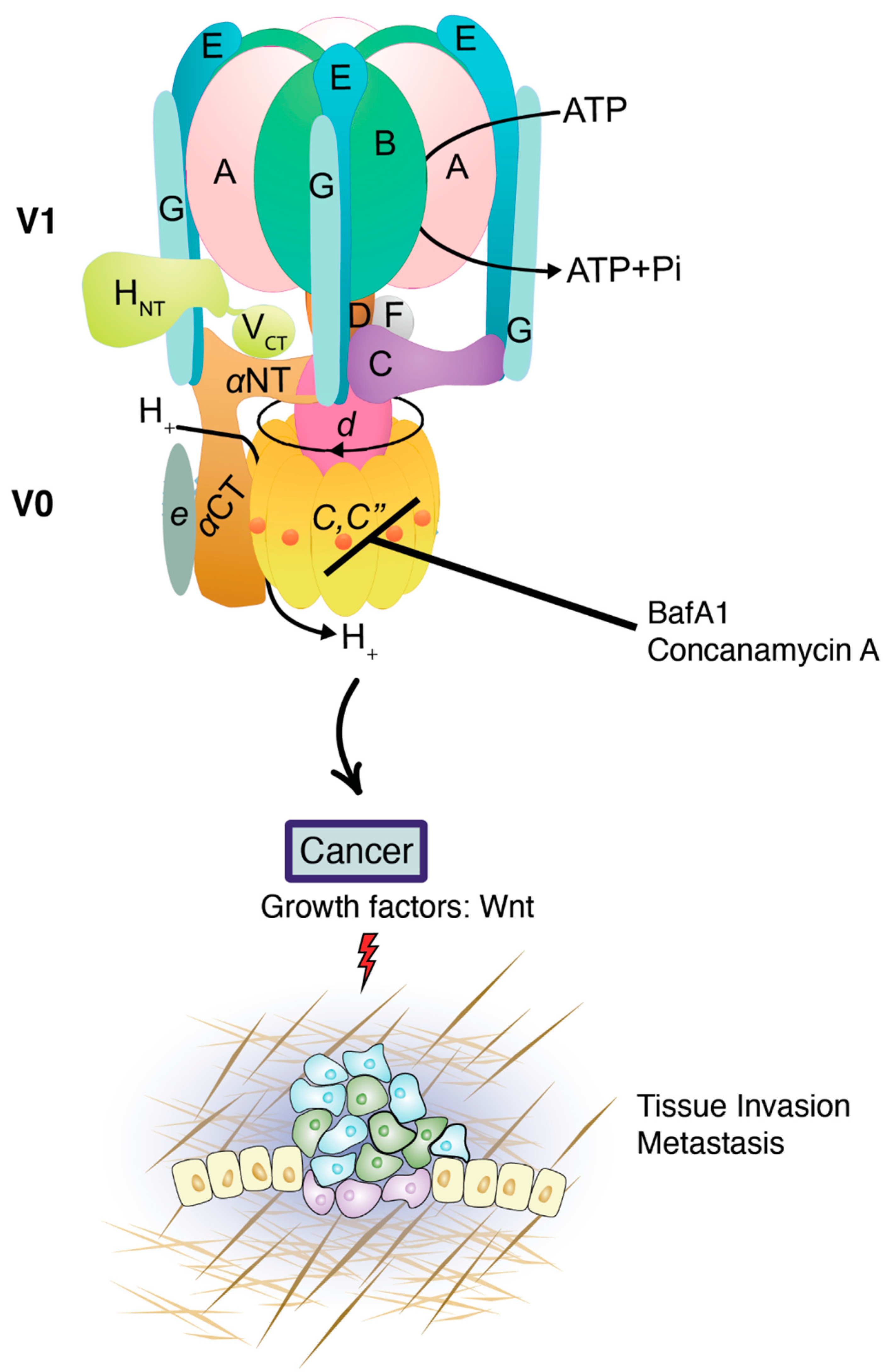
3. V-ATPase in Cancer
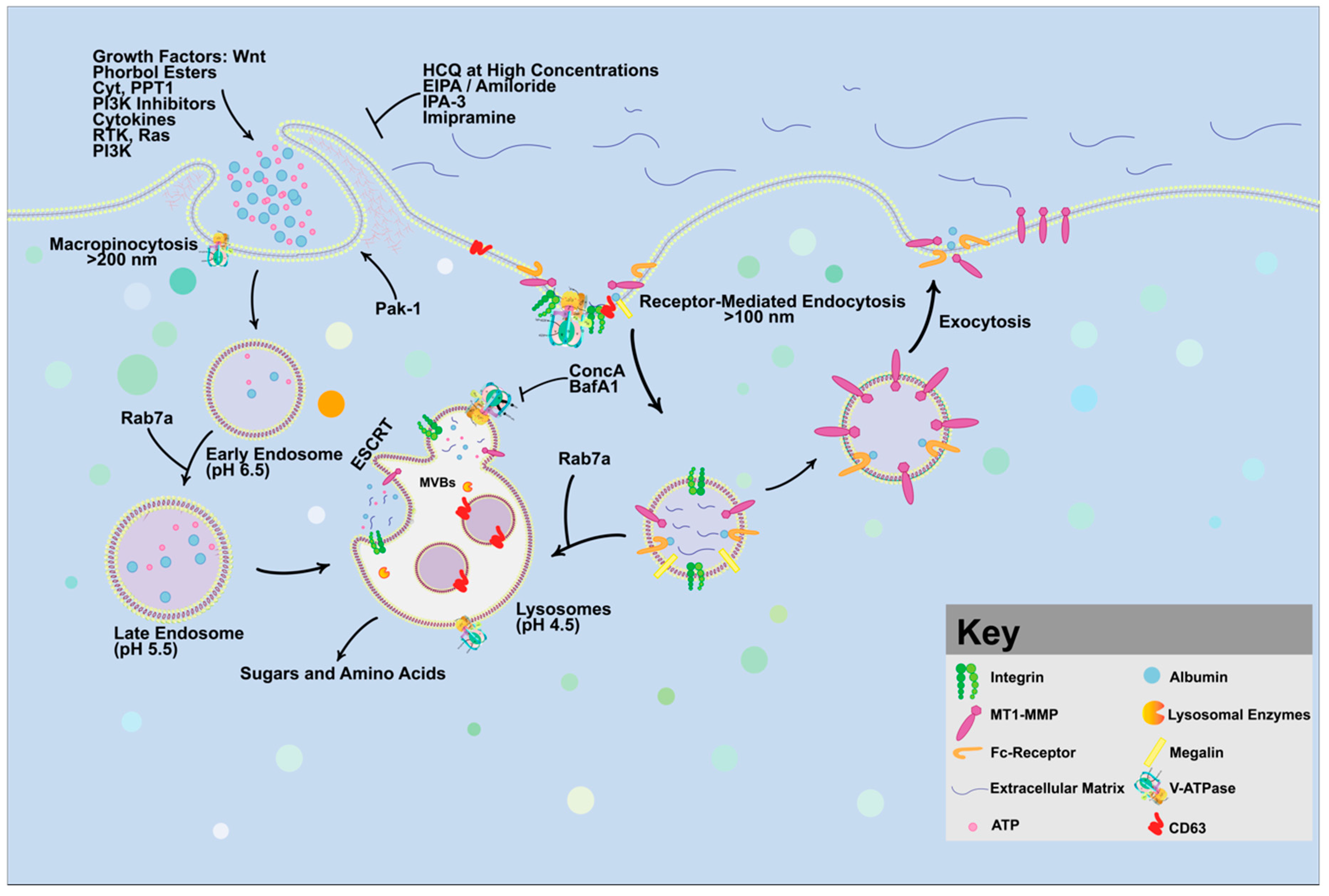
4. Targeting Macropinocytosis as a Strategy for Cancer Treatment
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Di Guglielmo, G.M.; Le Roy, C.; Goodfellow, A.F.; Wrana, J.L. Distinct Endocytic Pathways Regulate TGF-β Receptor Signalling and Turnover. Nat. Cell Biol. 2003, 5, 410–421. [Google Scholar] [CrossRef] [PubMed]
- Komiya, Y.; Habas, R. Wnt Signal Transduction Pathways. Organogenesis 2008, 4, 68–75. [Google Scholar] [CrossRef] [Green Version]
- Cabrera, C.V.; Alonso, M.C.; Johnston, P.; Phillips, R.G.; Lawrence, P.A. Phenocopies Induced with Antisense RNA Identify the Wingless Gene. Cell 1987, 50, 659–663. [Google Scholar] [CrossRef]
- Nusse, R.; Varmus, H.E. Many Tumors Induced by the Mouse Mammary Tumor Virus Contain a Provirus Integrated in the Same Region of the Host Genome. Cell 1982, 31, 99–109. [Google Scholar] [CrossRef]
- Takada, S.; Fujimori, S.; Shinozuka, T.; Takada, R.; Mii, Y. Differences in the secretion and transport of Wnt proteins. J. Biochem. 2017, 161, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilić, J.; Huang, Y.-L.; Davidson, G.; Zimmermann, T.; Cruciat, C.-M.; Bienz, M.; Niehrs, C. Wnt Induces LRP6 Signalosomes and Promotes Dishevelled-Dependent LRP6 Phosphorylation. Science 2007, 316, 1619–1622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blitzer, J.T.; Nusse, R. A Critical Role for Endocytosis in Wnt Signaling. BMC Cell Biol. 2006, 7, 28. [Google Scholar] [CrossRef] [Green Version]
- Stamos, J.L.; Weis, W.I. The β-Catenin Destruction Complex. Cold Spring Harb. Perspect. Biol. 2012, 5, a007898. [Google Scholar]
- Behrens, J.; von Kries, J.P.; Kühl, M.; Bruhn, L.; Wedlich, D.; Grosschedl, R.; Birchmeier, W. Functional Interaction of β-Catenin with the Transcription Factor LEF-1. Nature 1996, 382, 638–642. [Google Scholar] [CrossRef]
- Taelman, V.F.; Dobrowolski, R.; Plouhinec, J.-L.; Fuentealba, L.C.; Vorwald, P.P.; Gumper, I.; Sabatini, D.D.; De Robertis, E.M. Wnt Signaling Requires Sequestration of Glycogen Synthase Kinase 3 inside Multivesicular Endosomes. Cell 2010, 143, 1136–1148. [Google Scholar] [CrossRef] [Green Version]
- Acebron, S.P.; Karaulanov, E.; Berger, B.S.; Huang, Y.-L.; Niehrs, C. Mitotic Wnt Signaling Promotes Protein Stabilization and Regulates Cell Size. Mol. Cell 2014, 54, 663–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobrowolski, R.; Vick, P.; Ploper, D.; Gumper, I.; Snitkin, H.; Sabatini, D.D.; De Robertis, E.M. Presenilin Deficiency or Lysosomal Inhibition Enhances Wnt Signaling through Relocalization of GSK3 to the Late-Endosomal Compartment. Cell Rep. 2012, 2, 1316–1328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, A.; Pagare, P.P.; Li, J.; Zhang, Y. Drug discovery efforts toward inhibitors of canonical Wnt/β-catenin signaling pathway in the treatment of cancer: A composition-of-matter review (2010–2020). Drug Discov. Today 2021, 27, 1115–1127. [Google Scholar] [CrossRef]
- Mosesson, Y.; Mills, G.B.; Yarden, Y. Derailed Endocytosis: An Emerging Feature of Cancer. Nat. Rev. Cancer 2008, 8, 835–850. [Google Scholar] [CrossRef]
- Tejeda-Muñoz, N.; De Robertis, E.M. Wnt, GSK3, and Macropinocytosis. Subcell. Biochem. 2022, 98, 169–187. [Google Scholar] [PubMed]
- Colozza, G.; Jami-Alahmadi, Y.; Dsouza, A.; Tejeda-Muñoz, N.; Albrecht, L.V.; Sosa, E.A.; Wohlschlegel, J.A.; De Robertis, E.M. Wnt-inducible Lrp6-APEX2 interacting proteins identify ESCRT machinery and Trk-fused gene as components of the Wnt signaling pathway. Sci. Rep. 2020, 10, 21555. [Google Scholar] [CrossRef]
- Stenmark, H. Rab GTPases as Coordinators of Vesicle Traffic. Nat. Rev. Mol. Cell Biol. 2009, 10, 513–525. [Google Scholar] [CrossRef]
- Chavrier, P.; Goud, B. The Role of Arf and Rab Gtpases in Membrane Transport. Curr. Opin. Cell 1999, 11, 466–475. [Google Scholar] [CrossRef]
- Pereira-Leal, J.B.; Seabra, M.C. The Mammalian Rab Family of Small GTPases: Definition of Family and Subfamily Sequence Motifs Suggests a Mechanism for Functional Specificity in the Ras Superfamily. J. Mol. Biol. 2000, 301, 1077–1087. [Google Scholar] [CrossRef] [Green Version]
- Tzeng, H.-T.; Wang, Y.-C. Rab-Mediated Vesicle Trafficking in Cancer. J. Biomed. Sci. 2016, 23, 70. [Google Scholar] [CrossRef] [Green Version]
- Gopal Krishnan, P.D.; Golden, E.; Woodward, E.A.; Pavlos, N.J.; Blancafort, P. Rab GTPases: Emerging Oncogenes and Tumor Suppressive Regulators for the Editing of Survival Pathways in Cancer. Cancers 2020, 12, 259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tejeda-Muñoz, N.; Albrecht, L.V.; Bui, M.H.; De Robertis, E.M. Wnt Canonical Pathway Activates Macropinocytosis and Lysosomal Degradation of Extracellular Proteins. Proc. Natl. Acad. Sci. USA 2019, 116, 10402–10411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.P.; Gleeson, P.A. Macropinocytosis: An Endocytic Pathway for Internalising Large Gulps. Immunol. Cell Biol. 2011, 89, 836–843. [Google Scholar] [CrossRef] [PubMed]
- Swanson, J.A.; King, J.S. The Breadth of Macropinocytosis Research. Philos. Trans. R. Soc. 2018, 374, 20180146. [Google Scholar] [CrossRef]
- Albrecht, L.V.; Tejeda-Muñoz, N.; Bui, M.H.; Cicchetto, A.C.; Di Biagio, D.; Colozza, G.; Schmid, E.; Piccolo, S.; Christofk, H.R.; De Robertis, E.M. GSK3 Inhibits Macropinocytosis and Lysosomal Activity through the Wnt Destruction Complex Machinery. Cell Rep. 2020, 32, 107973. [Google Scholar] [CrossRef]
- Tejeda-Munoz, N.; De Robertis, E.M. Lysosomes are required for early dorsal signaling in the Xenopus embryo. Proc. Natl. Acad. Sci. USA 2022, 119, e2201008119. [Google Scholar] [CrossRef]
- Yan, L.; Chen, J.; Zhu, X.; Sun, J.; Wu, X.; Shen, W.; Zhang, W.; Tao, Q.; Meng, A. Maternal Huluwa dictates the embryonic body axis through β-catenin in vertebrates. Science 2018, 23, 362. [Google Scholar] [CrossRef]
- Tejeda-Muñoz, N.; Morselli, M.; Moriyama, Y.; Sheladiya, P.; Pellegrini, M.; De Robertis, E.M. Canonical Wnt Signaling Induces Focal Adhesion and Integrin Beta-1 Endocytosis. iScience 2022, 25, 104123. [Google Scholar] [CrossRef]
- Ramirez, C.; Hauser, A.D.; Vucic, E.A.; Bar-Sagi, D. Plasma membrane V-ATPase controls oncogenic RAS-induced macropinocytosis. Nature 2019, 576, 477–481. [Google Scholar] [CrossRef]
- Stevens, T.H.; Forgac, M. Structure, Function and Regulation of the Vacuolar (H+)-ATPase. Annu. Rev. Cell Dev. Biol. 1997, 13, 779–808. [Google Scholar] [CrossRef]
- Kotyk, A.; Dvoráková, M.; Koryta, J. Deuterons cannot replace protons in active transport processes in yeast. FEBS Lett. 1990, 264, 203–205. [Google Scholar] [CrossRef] [Green Version]
- Boros, L.G.; D’Agostino, D.P.; Katz, H.E.; Roth, J.P.; Meuillet, E.J.; Somlyai, G. Submolecular regulation of cell transformation by deuterium depleting water exchange reactions in the tricarboxylic acid substrate cycle. Med. Hypotheses 2016, 87, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Arata, Y.; Nishi, T.; Kawasaki-Nishi, S.; Shao, E.; Wilkens, S.; Forgac, M. Structure, Subunit Function and Regulation of the Coated Vesicle and Yeast Vacuolar (H+)-ATPases. Biochim. Biophys. Acta Bioenerg. 2002, 1555, 71–74. [Google Scholar] [CrossRef] [Green Version]
- Eaton, A.F.; Merkulova, M.; Brown, D. The H+-ATPase (V-ATPase): From Proton Pump to Signaling Complex in Health and Disease. Am. J. Physiol. Cell Physiol. 2021, 320, C392–C414. [Google Scholar] [CrossRef] [PubMed]
- Stransky, L.; Cotter, K.; Forgac, M. The Function of V-Atpases in Cancer. Physiol. Rev. 2016, 96, 1071–1091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perona, R.; Serrano, R. Increased pH and tumorigenicity of fibroblasts expressing a yeast proton pump. Nature 1988, 334, 438–440. [Google Scholar] [CrossRef]
- Hinton, A.; Sennoune, S.R.; Bond, S.; Fang, M.; Reuveni, M.; Sahagian, G.G.; Jay, D.; Martinez-Zaguilan, R.; Forgac, M. Function of a Subunit Isoforms of the V-Atpase in Ph Homeostasis and in Vitro Invasion of MDA-MB231 Human Breast Cancer Cells. JBC 2009, 284, 16400–16408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capecci, J.; Forgac, M. The Function of Vacuolar Atpase (V-Atpase) a Subunit Isoforms in Invasiveness of MCF10a and mcf10ca1a Human Breast Cancer Cells. JBC 2013, 288, 32731–32741. [Google Scholar] [CrossRef] [Green Version]
- García-García, A.; Pérez-Sayáns, M.; Rodríguez, M.J.; Antúnez-López, J.; Barros-Angueira, F.; Somoza-Martín, M.; Gándara-Rey, J.M.; Aguirre-Urízar, J.M. Immunohistochemical localization of C1 subunit of V-ATPase (ATPase C1) in oral squamous cell cancer and normal oral mucosa. Biotech. Histochem. 2012, 7, 133–139. [Google Scholar] [CrossRef]
- Torigoe, T.; Izumi, H.; Ishiguchi, H.; Uramoto, H.; Murakami, T.; Ise, T.; Yoshida, Y.; Tanabe, M.; Nomoto, M.; Itoh, H.; et al. Enhanced expression of the human vacuolar H+-ATPase c subunit gene (ATP6L) in response to anticancer agents. J. Biol. Chem. 2002, 277, 36534–36543. [Google Scholar] [CrossRef] [Green Version]
- Laplante, M.; Sabatini, D.M. mTOR signaling in growth control and disease. Cell 2012, 149, 274–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Schwarzenberg, K.; Wiedmann, R.M.; Oak, P.; Schulz, S.; Zischka, H.; Wanner, G.; Efferth, T.; Trauner, D.; Vollmar, A.M. Mode of cell death induction by pharmacological vacuolar H(+)-ATPase (V-ATPase) inhibition. JBC 2013, 288, 1385–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotter, K.; Capecci, J.; Sennoune, S.; Huss, M.; Maier, M.; Martinez-Zaguilan, R.; Forgac, M. Activity of Plasma Membrane V-Atpases Is Critical for the Invasion of MDA-MB231 Breast Cancer Cells. JBC 2015, 290, 3680–3692. [Google Scholar] [CrossRef] [Green Version]
- Sennoune, S.R.; Bakunts, K.; Martínez, G.M.; Chua-Tuan, J.L.; Kebir, Y.; Attaya, M.N.; Martínez-Zaguilán, R. Vacuolar H+-ATPase in Human Breast Cancer Cells with Distinct Metastatic Potential: Distribution and Functional Activity. Am. J. Physiol. Cell Physiol. 2004, 286, C1443–C1452. [Google Scholar] [CrossRef] [PubMed]
- Cruciat, C.-M.; Ohkawara, B.; Acebron, S.P.; Karaulanov, E.; Reinhard, C.; Ingelfinger, D.; Boutros, M.; Niehrs, C. Requirement of Prorenin Receptor and Vacuolar H+-ATPase-Mediated Acidification for Wnt Signaling. Science 2010, 327, 459–463. [Google Scholar] [CrossRef]
- Jung, Y.-S.; Jun, S.; Kim, M.J.; Lee, S.H.; Suh, H.N.; Lien, E.M.; Jung, H.-Y.; Lee, S.; Zhang, J.; Yang, J.-I.; et al. TMEM9 Promotes Intestinal Tumorigenesis through Vacuolar-Atpase-Activated Wnt/β-Catenin Signalling. Nat. Cell Biol. 2018, 20, 1421–1433. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.S.; Stratton, S.A.; Lee, S.H.; Kim, M.J.; Jun, S.; Zhang, J.; Zheng, B.; Cervantes, C.L.; Cha, J.H.; Barton, M.C.; et al. TMEM9-V-ATPase Activates Wnt/β-Catenin Signaling via APC Lysosomal Degradation for Liver Regeneration and Tumorigenesis. Hepatology 2020, 73, 776–794. [Google Scholar] [CrossRef]
- Nakase, I.; Kobayashi, N.B.; Takatani-Nakase, T.; Yoshida, T. Active Macropinocytosis Induction by Stimulation of Epidermal Growth Factor Receptor and Oncogenic RAS Expression Potentiates Cellular Uptake Efficacy of Exosomes. Sci. Rep. 2015, 5, 10300. [Google Scholar] [CrossRef] [Green Version]
- Ghoshal, P.; Singla, B.; Lin, H.; Feck, D.M.; Cantu-Medellin, N.; Kelley, E.E.; Haigh, S.; Fulton, D.; Csányi, G. Nox2-Mediated PI3K and Cofilin Activation Confers Alternate Redox Control of Macrophage Pinocytosis. ARS 2017, 26, 902–916. [Google Scholar] [CrossRef]
- Kitazawa, S.; Nishizawa, S.; Nakagawa, H.; Funata, M.; Nishimura, K.; Soga, T.; Hara, T. Cancer with Low Cathepsin D Levels Is Susceptible to Vacuolar (H+)-ATPase Inhibition. Cancer Sci. 2017, 108, 1185–1193. [Google Scholar] [CrossRef] [Green Version]
- Takenaka, T.; Nakai, S.; Katayama, M.; Hirano, M.; Ueno, N.; Noguchi, K.; Takatani-Nakase, T.; Fujii, I.; Kobayashi, S.S.; Nakase, I. Effects of Gefitinib Treatment on Cellular Uptake of Extracellular Vesicles in EGFR-Mutant Non-Small Cell Lung Cancer Cells. Int. J. Pharm. 2019, 572, 118762. [Google Scholar] [CrossRef] [PubMed]
- Tajiri, H.; Uruno, T.; Shirai, T.; Takaya, D.; Matsunaga, S.; Setoyama, D.; Watanabe, M.; Kukimoto-Niino, M.; Oisaki, K.; Ushijima, M.; et al. Targeting Ras-Driven Cancer Cell Survival and Invasion through Selective Inhibition of dock1. Cell Rep. 2017, 19, 969–980. [Google Scholar] [CrossRef] [Green Version]
- Amyere, M.; Payrastre, B.; Krause, U.; Smissen, P.V.; Veithen, A.; Courtoy, P.J. Constitutive Macropinocytosis in Oncogene-Transformed Fibroblasts Depends on Sequential Permanent Activation of Phosphoinositide 3-Kinase and Phospholipase C. Mol. Biol. Cell 2000, 11, 3453–3467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Straight, A.F.; Cheung, A.; Limouze, J.; Chen, I.; Westwood, N.J.; Sellers, J.R.; Mitchison, T.J. Dissecting Temporal and Spatial Control of Cytokinesis with a Myosin II Inhibitor. Science 2003, 299, 1743–1747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araki, N.; Egami, Y.; Watanabe, Y.; Hatae, T. Phosphoinositide Metabolism during Membrane Ruffling and Macropinosome Formation in EGF-Stimulated A431 Cells. Exp. Cell Res. 2007, 313, 1496–1507. [Google Scholar] [CrossRef]
- Towers, C.G.; Thorburn, A. Targeting the Lysosome for Cancer Therapy. Cancer Discov. 2017, 7, 1218–1220. [Google Scholar] [CrossRef] [Green Version]
- Michalopoulou, E.; Auciello, F.R.; Bulusu, V.; Strachan, D.; Campbell, A.D.; Tait-Mulder, J.; Karim, S.A.; Morton, J.P.; Sansom, O.J.; Kamphorst, J.J. Macropinocytosis Renders a Subset of Pancreatic Tumor Cells Resistant to Mtor Inhibition. Cell Rep. 2020, 30, 2729–2742. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wnt/β-catenin Pathway Inhibitors | Name |
|---|---|
| Repressor targeting Wnt ligand | sFRP1 (FRP, SARP2, FrzA) SFRP1, sFRP2 (SARP1) SFRP2 sFRP3 (FrzB, Fritz) FRZB, sFRP4 (FrzB-2) SFRP4, sFRP5 (SARP3) SFRP5 Sizzled, Sizzled2, Crescent, WIF-1, Tiki, Cerberus, Notum, Coco, Dkk-3 (REIC) (DKK3), Soggy (DKKL2), Ipafricept, OMP-18R5, F2.A, IGFBP4, Fz7-21, OTSA-101, Gpr177, Wise, 90γ-OTSA-101, OMP-54F28 |
| Repressor targeting Lrps | Dkk-1 (DKK1), Sost, Dkk-2 (DKK2), Dkk-4 (DKK4) |
| Repressor targeting Fzl | sFRP1 (FRP, SARP2, FrzA) SFRP1 (inhibits at high concentrations), IGFBP4, OTSA101, OMP-18R5, OMP-54F28 |
| PORC inhibitors | WNT974, CGX1321, IWP-2, ETC-159, RXC004, GNF-6231, 90γ-OTSA-101, LGK974 |
| β-catenin/TCF inhibitors | PFK115-584, CGP049090, CWP291, FL3, ZINC02092166, NC043, iCRT14 |
| CBP/ β-catenin binding inhibitors | PRI-724, ICG001, GNE-781, JW67, JW74, NLS-StAx-h, INT-01 |
| DVL inhibitors | FJ9, NSC668036, 3289-8625, Niclosamide, J01-017a, sulindac, LM02 |
| Repressor targeting Axin | Tankyrase inhibitors; XAV939, IWR-1, NVP-TNKS656, LZZ-02, JW74, WIKI14, K-756, G007-LK, G244-LM, FL3 |
| β-catenin inhibitors | COX inhibitors; Aspirin, Celecoxib, Sulindac, 1,25(OH)2D35R, SM08502, PKF115-584, PKF118-310, SAH-BCL9 |
| Repressor targeting CKI | Pyrivinium |
| Repressor targeting GSK3β | Genistein |
| TCF/LEF inhibitors | TNIK inhibitor, NCB-0846, PKF115-584, CGP049090 |
| Repressor targeting DKK | DKN-01 |
| Regulates alternative splicing of TCF inhibitors | SAM68, OMP-54F28 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tejeda-Muñoz, N.; Mei, K.-C.; Sheladiya, P.; Monka, J. Targeting Membrane Trafficking as a Strategy for Cancer Treatment. Vaccines 2022, 10, 790. https://doi.org/10.3390/vaccines10050790
Tejeda-Muñoz N, Mei K-C, Sheladiya P, Monka J. Targeting Membrane Trafficking as a Strategy for Cancer Treatment. Vaccines. 2022; 10(5):790. https://doi.org/10.3390/vaccines10050790
Chicago/Turabian StyleTejeda-Muñoz, Nydia, Kuo-Ching Mei, Pooja Sheladiya, and Julia Monka. 2022. "Targeting Membrane Trafficking as a Strategy for Cancer Treatment" Vaccines 10, no. 5: 790. https://doi.org/10.3390/vaccines10050790
APA StyleTejeda-Muñoz, N., Mei, K.-C., Sheladiya, P., & Monka, J. (2022). Targeting Membrane Trafficking as a Strategy for Cancer Treatment. Vaccines, 10(5), 790. https://doi.org/10.3390/vaccines10050790