
Protease-Based Subunit Vaccine in Mice Boosts BCG Protection against Mycobacterium tuberculosis

 ,
, 
 , ,
, ,  and
and 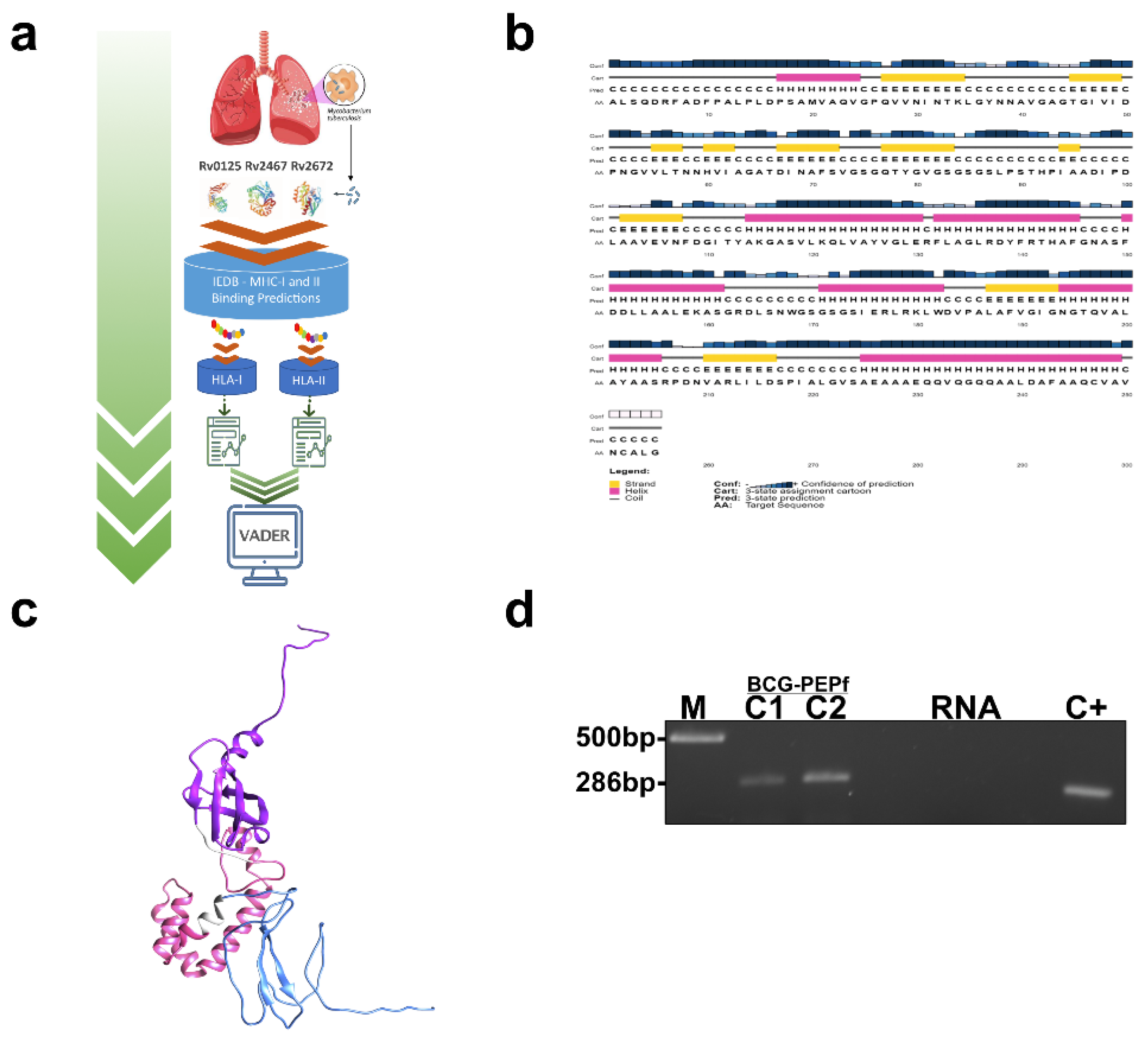{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Prediction of MHC (Major Histocompatibility Complex) Class I and Class II Binding Epitopes and Fusion Protein Design
2.2. Secondary and Tertiary Structure Prediction of PEPf
2.3. Bacterial Strains Used in This Study
2.4. Expression and Purification of Recombinant PEPf Protein
2.5. Animal Care and Use
2.6. PEPf Immunogenicity Evaluation Strategy
2.7. Construction of BCG-PEPf
2.8. Live Vaccine Preparation, Immunization, and Mtb Challenge
2.9. Booster Vaccine Strategy
2.10. Mycobacterium Tuberculosis Infection
2.11. Colony-Forming Unit (CFU) Determination of Infected Mice
2.12. Histopathological Evaluations
2.13. Immune Response Analysis by Flow Cytometry
2.14. Statistical Analysis
3. Results
3.1. Epitope Predictions
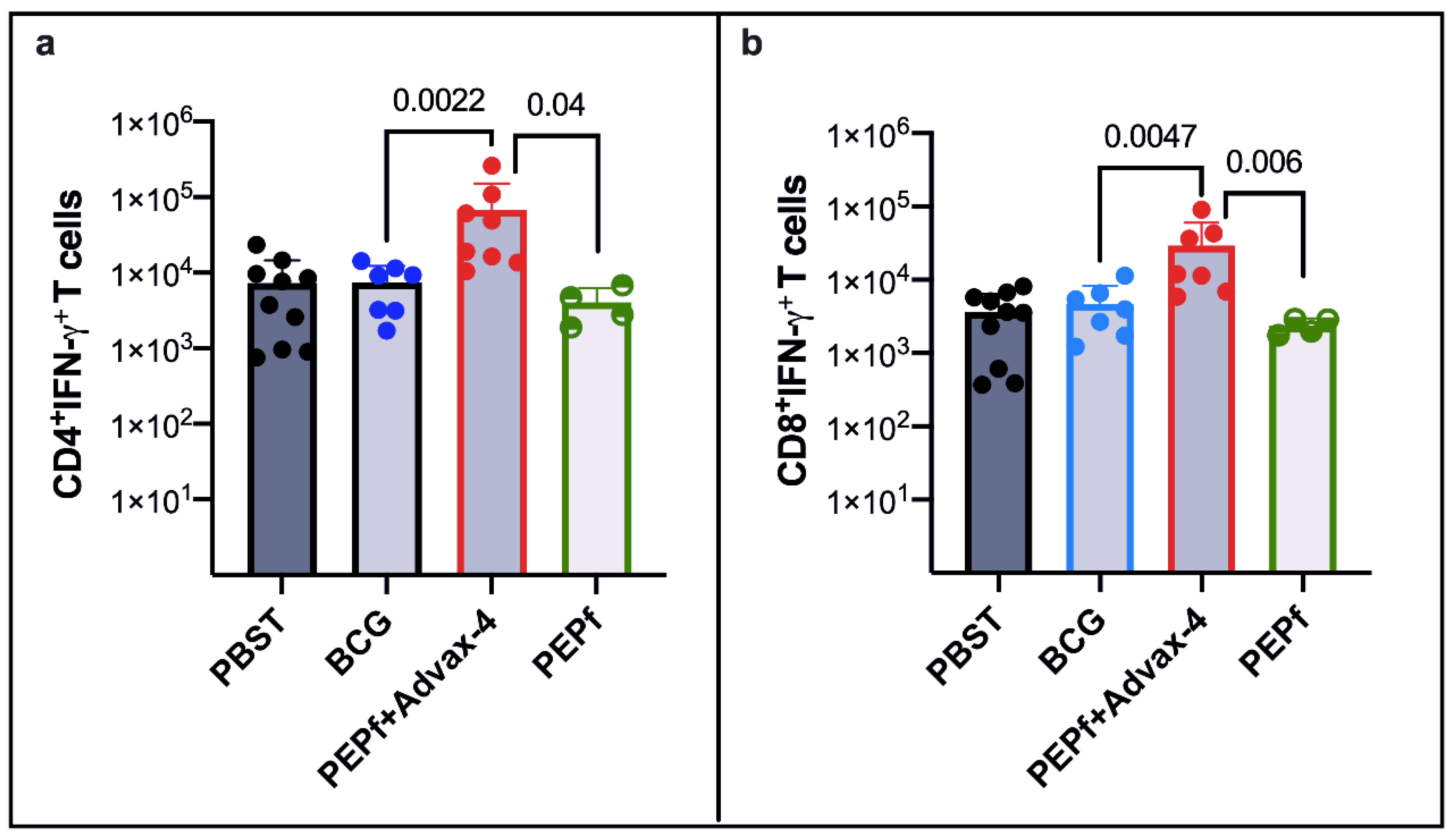
3.2. PEPf Protein Used as a Subunit Vaccine Is Immunogenic in the Murine Vaccination Model
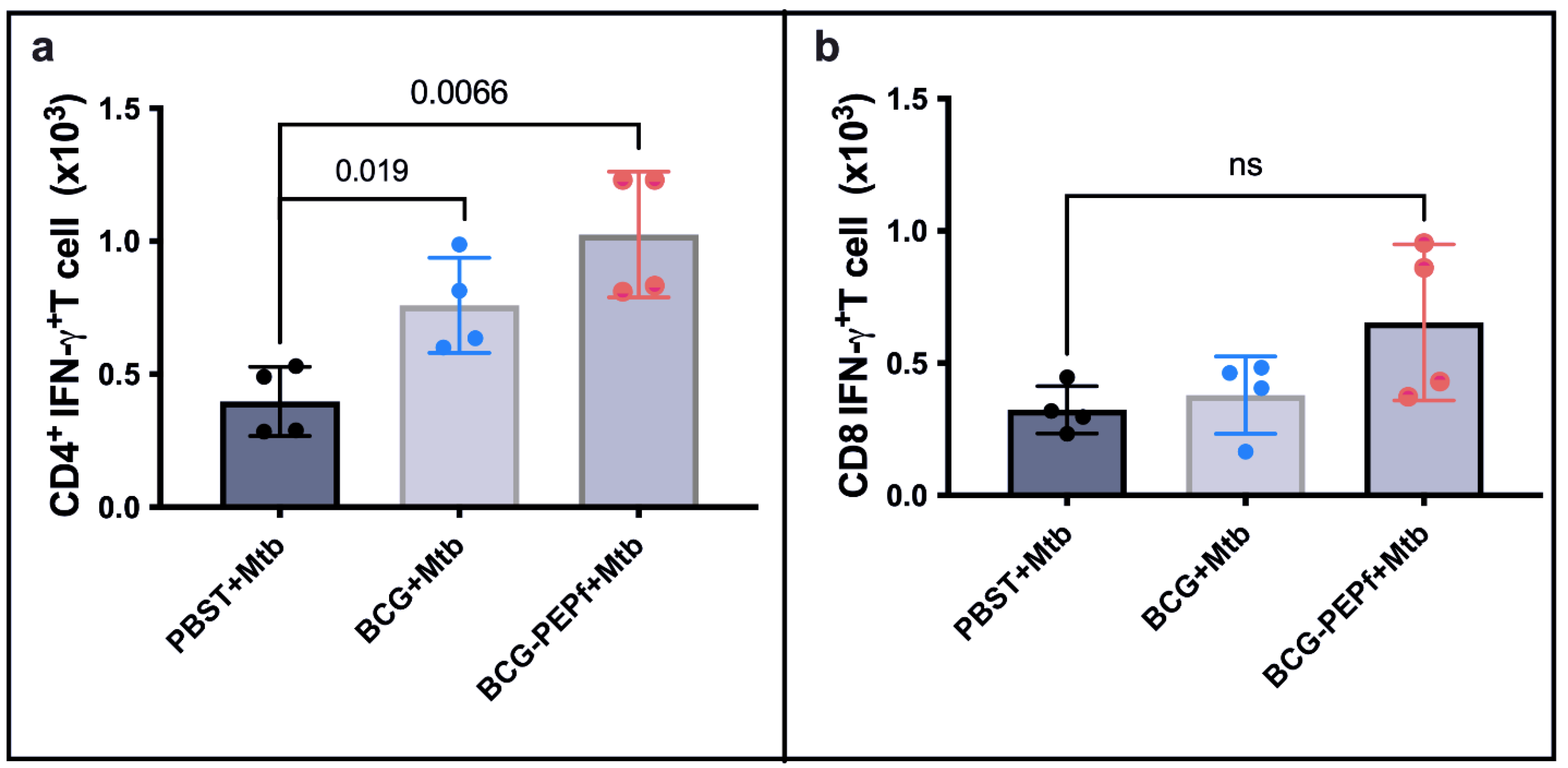
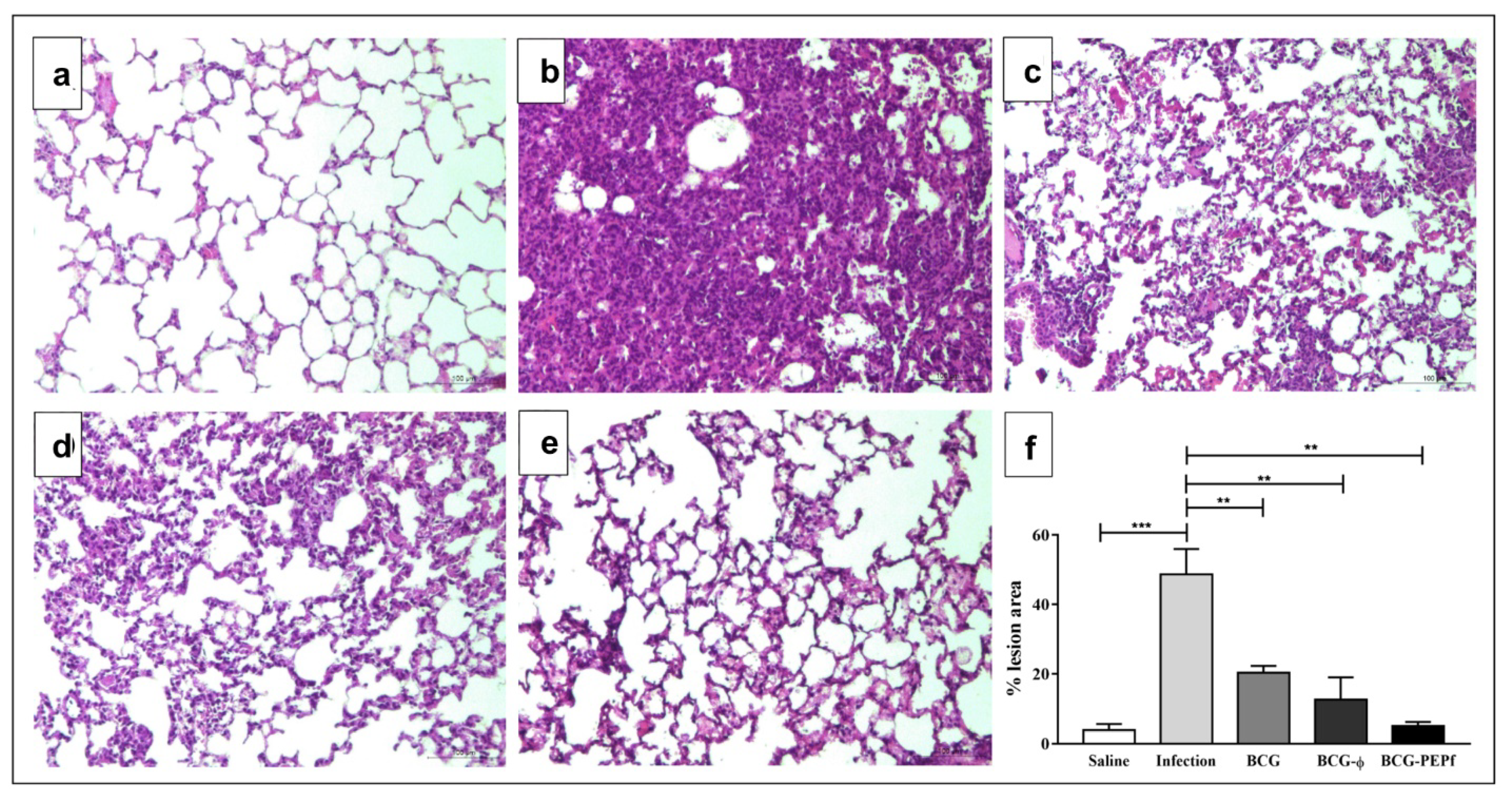
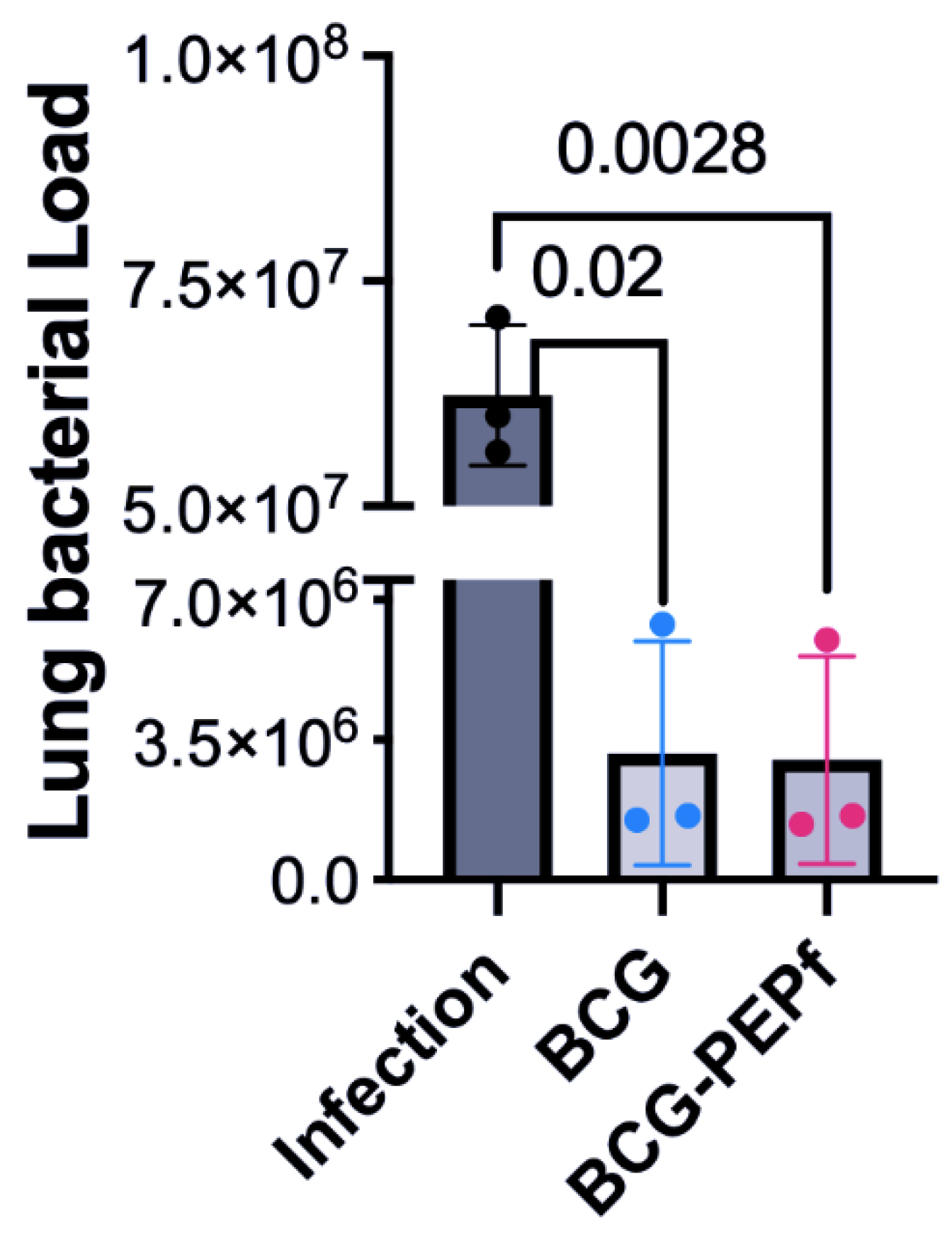
3.3. BCG-PEPf Vaccine Recalls Th1-Specific Immune Response after the M. tuberculosis Challenge and Diminishes the Lung Lesion Areas Caused by Infection
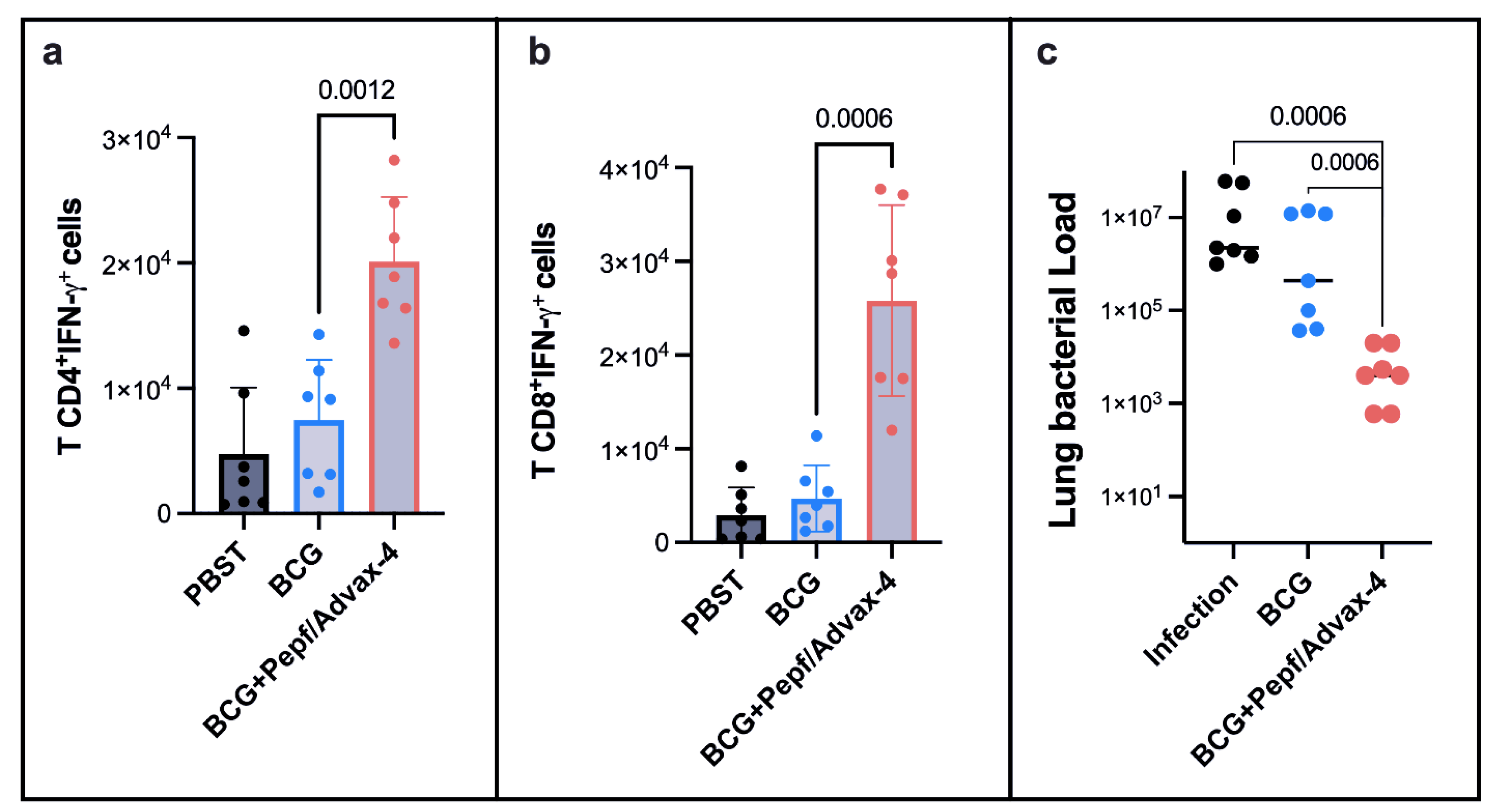
3.4. Prime-Boost Vaccine Strategies Using BCG and PEPf + Advax4 Reduced Lung Lesions and Enhanced BCG Protection against Tuberculosis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fletcher, H.A.; Schrager, L. TB Vaccine Development and the End TB Strategy: Importance and Current Status. Trans. R. Soc. Trop. Med. Hyg. 2016, 110, 212–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Global Tuberculosis Report 2021; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Husain, A.A.; Daginawala, H.F.; Singh, L.; Kashyap, R.S. Current Perspective in Tuberculosis Vaccine Development for High TB Endemic Regions. Tuberculosis 2016, 98, 149–158. [Google Scholar] [CrossRef]
- Luca, S.; Mihaescu, T. History of BCG Vaccine. Maedica 2013, 8, 53–58. [Google Scholar] [PubMed]
- Zwerling, A.; Behr, M.A.; Verma, A.; Brewer, T.F.; Menzies, D.; Pai, M. The BCG World Atlas: A Database of Global BCG Vaccination Policies and Practices. PLoS Med. 2011, 8, e1001012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butkeviciute, E.; Jones, C.E.; Smith, S.G. Heterologous Effects of Infant BCG Vaccination: Potential Mechanisms of Immunity. Future Microbiol. 2018, 13, 1193–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sette, A.; Rappuoli, R. Reverse Vaccinology: Developing Vaccines in the Era of Genomics. Immunity 2010, 33, 530–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rappuoli, R.; Bottomley, M.J.; D’Oro, U.; Finco, O.; De Gregorio, E. Reverse Vaccinology 2.0: Human Immunology Instructs Vaccine Antigen Design. J. Exp. Med. 2016, 213, 469–481. [Google Scholar] [CrossRef]
- Lata, K.S.; Kumar, S.; Vaghasia, V.; Sharma, P.; Bhairappanvar, S.B.; Soni, S.; Das, J. Exploring Leptospiral Proteomes to Identify Potential Candidates for Vaccine Design Against Leptospirosis Using an Immunoinformatics Approach. Sci. Rep. 2018, 8, 6935. [Google Scholar] [CrossRef] [Green Version]
- Sharma, N.; Aggarwal, S.; Kumar, S.; Sharma, R.; Choudhury, K.; Singh, N.; Jayaswal, P.; Goel, R.; Wajid, S.; Yadav, A.K.; et al. Comparative Analysis of Homologous Aminopeptidase PepN from Pathogenic and Non-pathogenic Mycobacteria Reveals Divergent Traits. PLoS ONE 2019, 14, e0215123. [Google Scholar] [CrossRef]
- Skeiky, Y.A.; Lodes, M.J.; Guderian, J.A.; Mohamath, R.; Bement, T.; Alderson, M.R.; Reed, S.G. Cloning, Expression, and Immunological Evaluation of Two Putative Secreted Serine Protease Antigens of Mycobacterium tuberculosis. Infect. Immun. 1999, 67, 3998–4007. [Google Scholar] [CrossRef] [Green Version]
- Tait, D.R.; Hatherill, M.; Der Meeren, O.V.; Ginsberg, A.M.; Brakel, E.V.; Salaun, B.; Scriba, T.J.; Akite, E.J.; Ayles, H.M.; Bollaerts, A.; et al. Final Analysis of a Trial of M72/AS01 E Vaccine to Prevent Tuberculosis. N. Engl. J. Med. 2019, 381, 2429–2439. [Google Scholar] [CrossRef] [PubMed]
- Dejesus, M.A.; Gerrick, E.R.; Xu, W.; Park, S.W.; Long, J.E.; Boutte, C.C.; Rubin, E.J.; Schnappinger, D.; Ehrt, S.; Fortune, S.M.; et al. Comprehensive Essentiality Analysis of the Mycobacterium Tuberculosis Genome via Saturating Transposon Mutagenesis. mBio 2017, 8, e02133-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, K.H.; Jha, B.; Dwivedy, A.; Choudhary, E.; Arpitha, G.N.; Ashraf, A.; Arora, D.; Agarwal, N.; Biswal, B.K. Characterization of a secretory hydrolase from Mycobacterium tuberculosis Sheds Critical Insight Into Host Lipid Utilization by M. tuberculosis. J. Biol. Chem. 2017, 292, 11326–11335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreatta, M.; Nielsen, M. Gapped Sequence Alignment Using Artificial Neural Networks: Application to the MHC Class I System. Bioinformatics 2016, 32, 511–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, B.; Sette, A. Generating Quantitative Models Describing the Sequence Specificity of Biological Processes with the Stabilized Matrix Method. BMC Bioinform. 2005, 6, 132. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, M.; Lund, O. NN-align. An Artificial Neural Network-based Alignment Algorithm for MHC Class II Peptide Binding Prediction. BMC Bioinform. 2009, 10, 296. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, M.; Lundegaard, C.; Lund, O. Prediction of MHC Class II Binding Affinity Using SMM-align, a Novel Stabilization Matrix Alignment Method. BMC Bioinform. 2007, 8, 238. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Sidney, J.; Dow, C.; Mothé, B.; Sette, A.; Peters, B. A Systematic Assessment of MHC Class II Peptide Binding Predictions and Evaluation of a Consensus Approach. PLoS Comput. Biol. 2008, 4, e1000048. [Google Scholar] [CrossRef] [Green Version]
- Dimitrov, I.; Naneva, L.; Doytchinova, I.; Bangov, I. AllergenFP: Allergenicity prediction by descriptor fingerprints. Bioinformatics 2014, 30, 846–851. [Google Scholar] [CrossRef]
- Saha, S.; Raghava, G.P.S. AlgPred: Prediction of Allergenic Proteins and Mapping of IgE Epitopes. Nucleic Acids Res. 2006, 34, W202–W209. [Google Scholar] [CrossRef]
- Venkatarajan, M.S.; Braun, W. New Quantitative Descriptors of Amino Acids Based on Multidimensional Scaling of a Large Number of Physical-Chemical Properties. J. Mol. Model. 2001, 7, 445–453. [Google Scholar] [CrossRef]
- Gupta, S.; Kapoor, P.; Chaudhary, K.; Gautam, A.; Kumar, R.; Raghava, G.P.S. In Silico Approach for Predicting Toxicity of Peptides and Proteins. PLoS ONE 2013, 8, e73957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Li, W.; Liu, S.; Xu, J. RaptorX-Property: A Web Server for Protein Structure Property Prediction. Nucleic Acids Res. 2016, 44, W430–W435. [Google Scholar] [CrossRef]
- Heo, L.; Park, H.; Seok, C. GalaxyRefine: Protein Structure Refinement Driven by Side-Chain Repacking. Nucleic Acids Res. 2013, 41, W384–W388. [Google Scholar] [CrossRef] [Green Version]
- Stover, C.K.; de la Cruz, V.F.; Fuerst, T.R.; Burlein, J.E.; Benson, L.A.; Bennett, L.T.; Bansal, G.P.; Young, J.F.; Lee, M.H.; Hatfull, G.F.; et al. New Use of BCG for Recombinant Vaccines. Nature 1991, 351, 456–460. [Google Scholar] [CrossRef]
- Goude, R.; Roberts, D.M.; Parish, T. Electroporation of Mycobacteria. Methods Mol. Biol. 2015, 1285, 117–1130. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, F.M.; Da Costa, A.C.; Procopio, V.O.; Garcia, W.; Araújo, J.N.; Da Silva, R.A.; Junqueira-Kipnis, A.P.; Kipnis, A. Mycobacterium abscessus Subsp. Massiliense mycma_0076 and mycma_0077 Genes Code for Ferritins That Are Modulated by Iron Concentration. Front. Microbiol. 2018, 9, 1072. [Google Scholar] [CrossRef] [Green Version]
- Junqueira-Kipnis, A.P.; De Oliveira, F.M.; Trentini, M.M.; Tiwari, S.; Chen, B.; Resende, D.P.; Silva, B.D.S.; Chen, M.; Tesfa, L.; Jacobs, W.R., Jr.; et al. Prime-boost with Mycobacterium smegmatis Recombinant Vaccine Improves Protection in Mice Infected with Mycobacterium tuberculosis. PLoS ONE 2013, 8, e78639. [Google Scholar] [CrossRef]
- Niuewenhuizen, N.E.; Kaufmann, S.H.E. Next-Generation Vaccines Based on Bacille Calmette-Guérin. Front. Immunol. 2018, 9, 121. [Google Scholar] [CrossRef]
- De Paula Oliveira Santos, B.; Trentini, M.M.; Machado, R.B.; Celes, M.R.N.; Kipnis, A.; Petrovsky, N.; Junqueira-Kipnis, A.P. Advax4 Delta Inulin Combination Adjuvant Together with ECMX, a Fusion Construct of Four Protective mTB Antigens, Induces a Potent Th1 Immune Response and Protects Mice against Mycobacterium tuberculosis Infection. Hum. Vaccines Immunother. 2017, 13, 2967–2976. [Google Scholar] [CrossRef] [Green Version]
- Da Costa, A.C.; Costa-Júnior, A.O.; Oliveira, F.M.; Nogueira, S.V.; Rosa, J.D.; Resende, D.P.; Kipnis, A.; Junqueira-Kipnis, A.P. A New Recombinant BCG Vaccine Induces Specific Th17 and Th1 Effector Cells with Higher Protective Efficacy against Tuberculosis. PLoS ONE 2014, 9, e112848. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Wang, X.; Zhang, H.; Xu, Z.; Li, Y.; Yuan, L. Immune Responses and Protective Efficacy Induced by 85B Antigen and Early Secreted Antigenic Target-6 kDa Antigen Fusion Protein Secreted by Recombinant Bacille Calmette-Guérin. Acta Biochim. Biophys. Sin. 2007, 39, 290–296. [Google Scholar] [CrossRef] [Green Version]
- Blevins, L.K.; Parsonage, D.; Oliver, M.B.; Domzalski, E.; Swords, W.E.; Alexander-Miller, M.A. A Novel Function for the Streptococcus pneumoniae Aminopeptidase N: Inhibition of T Cell Effector Function through Regulation of TCR Signaling. Front. Immunol. 2017, 8, 1610. [Google Scholar] [CrossRef] [Green Version]
- Kaushal, D.; Foreman, T.W.; Gautam, U.S.; Alvarez, X.; Adekambi, T.; Rangel-Moreno, J.; Golden, N.A.; Johnson, A.M.F.; Phillips, B.L.; Ahsan, M.H.; et al. Mucosal Vaccination with Attenuated Mycobacterium tuberculosis Induces Strong Central Memory Responses and Protects against Tuberculosis. Nat. Commun. 2015, 6, 8533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, A.M.; Difazio, R.; Flynn, J.L. IFN-γ From CD4 T Cells is Essential for Host Survival and Enhances CD8 T Cell Function During Mycobacterium Tuberculosis Infection. J. Immunol. 2013, 190, 270–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.D.; Mott, D.; Sutiwisesak, R.; Lu, Y.-J.; Raso, F.; Stowell, B.; Babunovic, G.H.; Lee, J.; Carpenter, S.M.; Way, S.S.; et al. Mycobacterium tuberculosis-specific CD4+ and CD8+ T Cells Differ in Their Capacity to Recognize Infected Macrophages. PLoS Pathog. 2018, 14, e1007060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saunders, B.M.; Frank, A.A.; Orme, I.M.; Cooper, A.M. CD4 is Required for the Development of a Protective Granulomatous Response to Pulmonary Tuberculosis. Cell. Immunol. 2002, 216, 65–72. [Google Scholar] [CrossRef]
- Orme, I.M.; Basaraba, R.J. The Formation of the Granuloma in Tuberculosis Infection. Semin. Immunol. 2014, 26, 601–609. [Google Scholar] [CrossRef]
- Cruz, A.; Khader, S.A.; Torrado, E.; Fraga, A.; Pearl, J.E.; Pedrosa, J.; Cooper, A.M.; Castro, A.G. Cutting Edge: IFN- regulates the Induction and Expansion of IL-17-producing CD4 T Cells during Mycobacterial Infection. J. Immunol. 2006, 177, 1416–1420. [Google Scholar] [CrossRef] [Green Version]
- Ottenhoff, T.H.; Kaufmann, S.H. Vaccines against Tuberculosis: Where Are We and Where Do We Need to Go? PLoS Pathog. 2012, 8, e1002607. [Google Scholar] [CrossRef]
- Behar, S.M. Antigen-specific CD8+ T Cells and Protective Immunity to Tuberculosis. Adv. Exp. Med. Biol. 2013, 783, 141–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Counoupas, C.; Pinto, R.; Nagalingam, G.; Britton, W.J.; Petrovsky, N.; Triccas, J.A. Delta Inulin-Based Adjuvants Promote the Generation of Polyfunctional CD4+ T Cell Responses and Protection against Mycobacterium tuberculosis Infection. Sci. Rep. 2017, 7, 8582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa, E.M.; da Costa, A.C.; Trentini, M.M.; de Araújo Filho, J.A.; Kipnis, A.; Junqueira-Kipnis, A.P. Immunogenicity of a Fusion Protein Containing Immunodominant Epitopes of Ag85C, MPT51, and HspX from Mycobacterium tuberculosis in Mice and Active TB Infection. PLoS ONE 2012, 7, e47781. [Google Scholar] [CrossRef] [PubMed]
- Lewinsohn, D.A.; Swarbrick, G.M.; Park, B.; Cansler, M.E.; Null, M.D.; Toren, K.G.; Baseke, J.; Zalwango, S.; Mayanja-Kizza, H.; Malone, L.L.; et al. Comprehensive Definition of Human Immunodominant CD8 Antigens in Tuberculosis. Npj Vaccines 2017, 2, 8. [Google Scholar] [CrossRef]
- Rao, M.; Cadieux, N.; Fitzpatrick, M.; Reed, S.; Arsenian, S.; Valentini, D.; Parida, S.; Dodoo, E.; Zumla, A.; Maeurer, M. Mycobacterium tuberculosis Proteins Involved in Cell Wall Lipid Biosynthesis Improve BCG Vaccine Efficacy in a Murine TB Model. Int. J. Infect. Dis. 2017, 56, 274–282. [Google Scholar] [CrossRef] [Green Version]
- Lin, P.L.; Flynn, J.L. CD8 T cells and Mycobacterium tuberculosis infection. Semin. Immunopathol. 2015, 37, 239–249. [Google Scholar] [CrossRef] [Green Version]
- Nieuwenhuizen, N.E.; Kulkarni, P.S.; Shaligram, U.; Cotton, M.F.; Rentsch, C.A.; Eisele, B.; Grode, L.; Kaufmann, S.H.E. The Recombinant Bacille Calmette-Guérin Vaccine VPM1002: Ready for Clinical Efficacy Testing. Front. Immunol. 2017, 8, 1147. [Google Scholar] [CrossRef]
- Tiwari, S.; Dutt, T.S.; Chen, B.; Chen, M.; Kim, J.; Dai, A.Z.; Lukose, R.; Shanley, C.; Fox, A.; Karger, B.R.; et al. BCG-Prime and Boost with Esx-5 Secretion System Deletion Mutant Leads to Better Protection against Clinical Strains of Mycobacterium tuberculosis. Vaccine 2020, 38, 7156–7165. [Google Scholar] [CrossRef]
- Kipnis, A.; Irwin, S.; Izzo, A.A.; Basaraba, R.J.; Orme, I.M. Memory T Lymphocytes Generated by Mycobacterium bovis BCG Vaccination Reside within a CD4 CD44lo CD62 Ligandhi Population. Infect. Immun. 2005, 73, 7759–7764. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Junqueira-Kipnis, A.P.; de Castro Souza, C.; de Oliveira Carvalho, A.C.; de Oliveira, F.M.; Almeida, V.P.; de Paula, A.R.; Celes, M.R.; Kipnis, A. Protease-Based Subunit Vaccine in Mice Boosts BCG Protection against Mycobacterium tuberculosis. Vaccines 2022, 10, 306. https://doi.org/10.3390/vaccines10020306
Junqueira-Kipnis AP, de Castro Souza C, de Oliveira Carvalho AC, de Oliveira FM, Almeida VP, de Paula AR, Celes MR, Kipnis A. Protease-Based Subunit Vaccine in Mice Boosts BCG Protection against Mycobacterium tuberculosis. Vaccines. 2022; 10(2):306. https://doi.org/10.3390/vaccines10020306
Chicago/Turabian StyleJunqueira-Kipnis, Ana Paula, Carine de Castro Souza, Ana Carolina de Oliveira Carvalho, Fabio Muniz de Oliveira, Vinnycius Pereira Almeida, Alisson Rodrigues de Paula, Mara Rubia Celes, and André Kipnis. 2022. "Protease-Based Subunit Vaccine in Mice Boosts BCG Protection against Mycobacterium tuberculosis" Vaccines 10, no. 2: 306. https://doi.org/10.3390/vaccines10020306
APA StyleJunqueira-Kipnis, A. P., de Castro Souza, C., de Oliveira Carvalho, A. C., de Oliveira, F. M., Almeida, V. P., de Paula, A. R., Celes, M. R., & Kipnis, A. (2022). Protease-Based Subunit Vaccine in Mice Boosts BCG Protection against Mycobacterium tuberculosis. Vaccines, 10(2), 306. https://doi.org/10.3390/vaccines10020306