
A Triple Gene-Deleted Pseudorabies Virus-Vectored Subunit PCV2b and CSFV Vaccine Protects Pigs against PCV2b Challenge and Induces Serum Neutralizing Antibody Response against CSFV

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Medium
2.2. Viruses
2.3. Antibodies
2.4. Construction of PRVtmv+ Vector Virus Expressing the CSFV E2, Erns-GM-CSF, and PCV2b Cap Proteins
2.4.1. Construction of PRV gE-Deleted Virus (PRV gEΔ)
2.4.2. Construction of PRV gE/TK Dual Gene-Deleted Virus
2.4.3. Construction of PRV gE/TK/gG Triple Gene-Deleted (PRVtmv) Vector
2.5. Construction of PRV gEΔ CSFV-E2, PRV TKΔ PCV2 Cap and gGΔ CSFV Erns-GM-CSF Insertion Plasmids
2.5.1. Construction of PRV gEΔ CSFV-E2 Insertion Plasmid (pPRVgEΔ/CSFV E2-INS)
2.5.2. Construction of PRV TK-Deleted PCV2 Cap Insertion Plasmid (pPRV TKΔ/PCV2 Cap-INS)
2.5.3. Construction of the gG Deletion/CSFV Erns-GM-CSF Insertion Plasmid (pPRV gGΔ/CSFV Erns-GM-CSF-INS)
2.5.4. Construction of PRVtmv Vector Virus Expressing CSFV E2 and Erns-GM-CSF, and PCV2b Cap Chimeric Genes (PRVtmv+)
2.6. Virus Titrations
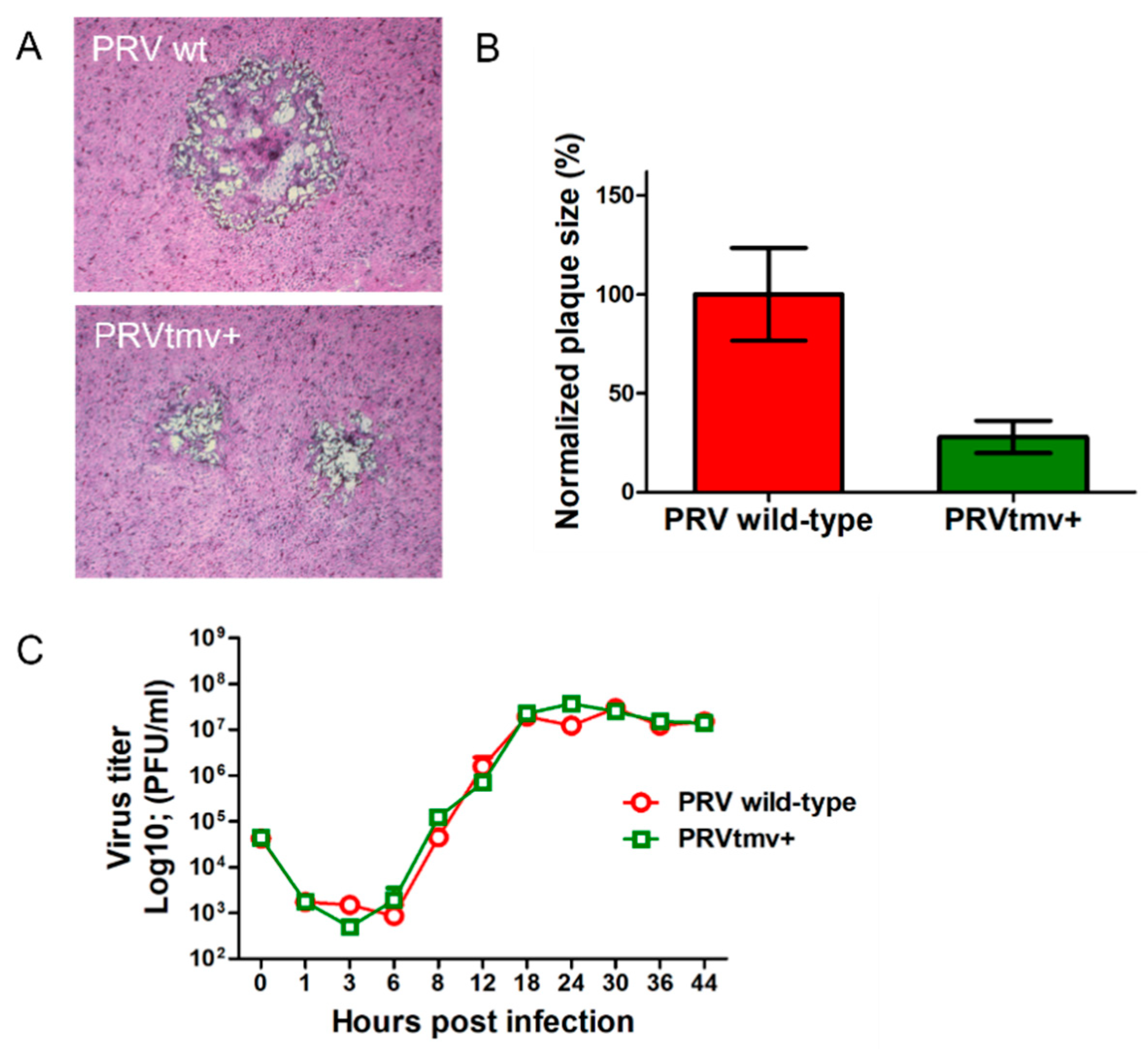
2.7. Growth Kinetics and Plaque Size Assay
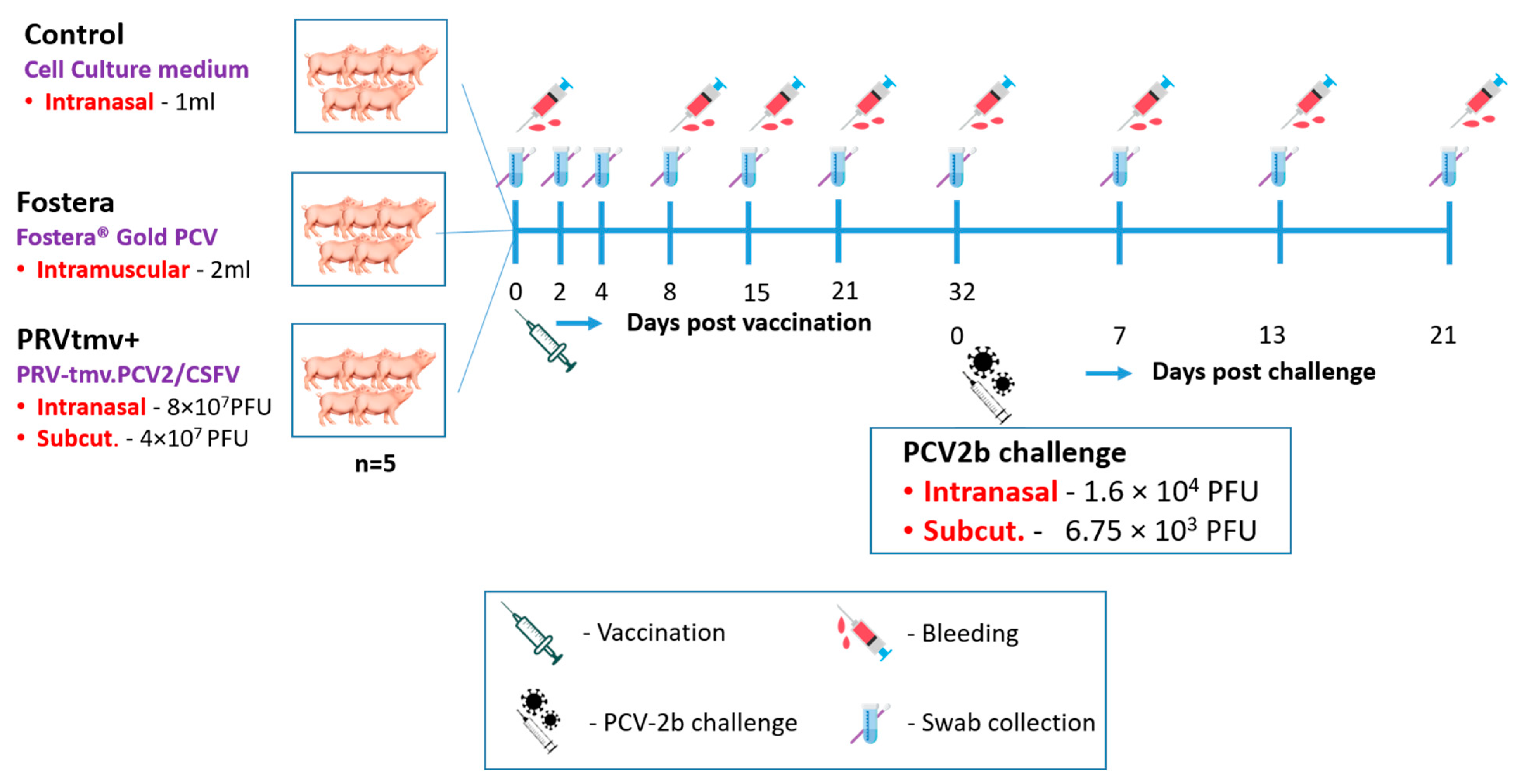
2.8. Animals and Experimental Design
2.8.1. Vaccination and Challenge
2.8.2. Clinical Examination of Pigs Following Vaccination and Challenge
2.8.3. Sample Collection and Processing from the Vaccinated and Control Pigs following Vaccination and PCV2b Challenge
2.8.4. Leukocyte and Lymphocyte Counting in Whole Blood
2.8.5. Euthanasia, Necropsy, Tissue Sample Collection, and Processing
2.9. Serum Virus Neutralization (SN) by Plaque Reduction Assay for PRV and PCV2b
2.10. CSFV-Specific SN Assay
2.11. DNA Isolation and Quantitative PCR (qPCR)
2.12. Immunohistochemistry
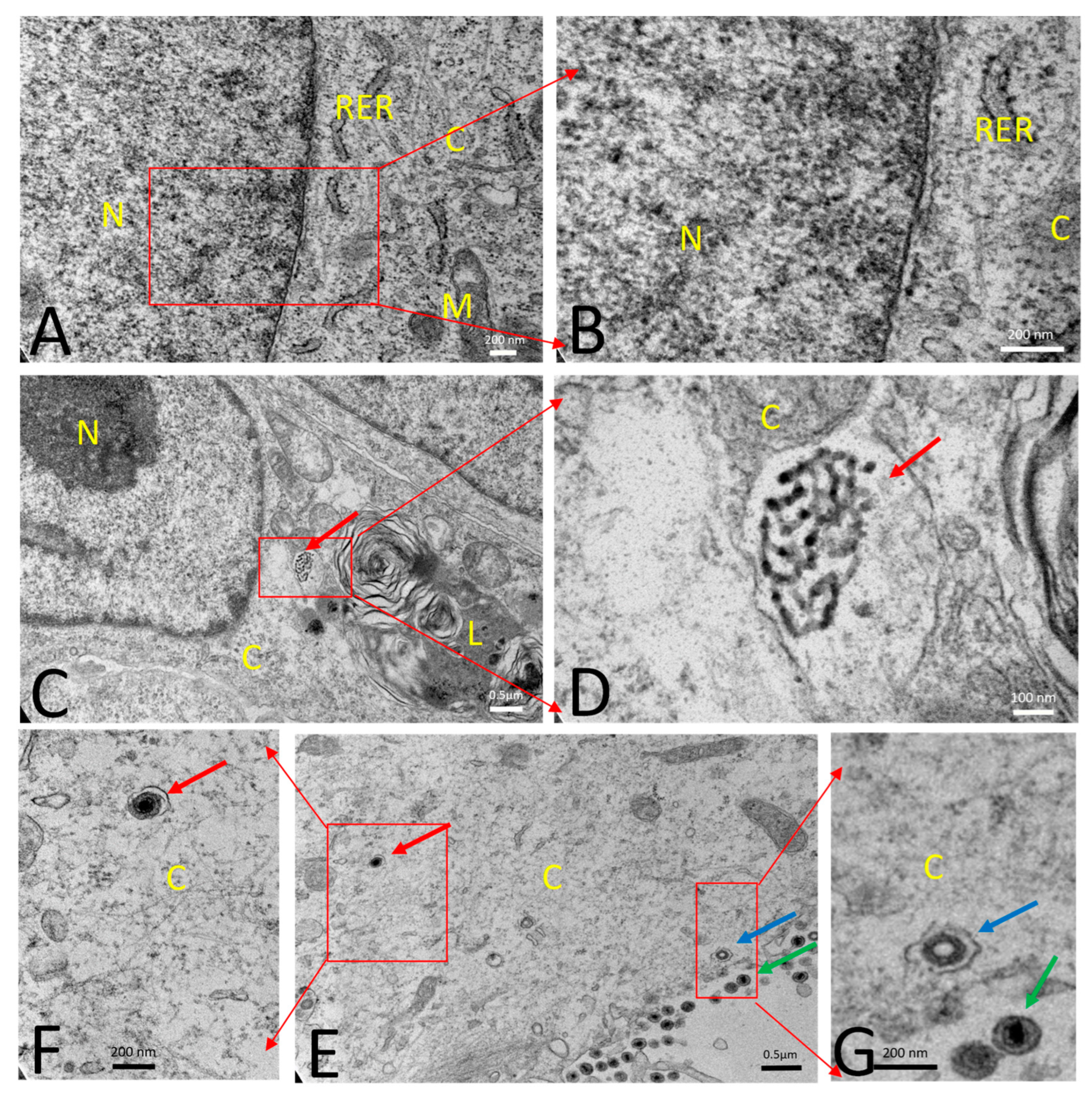
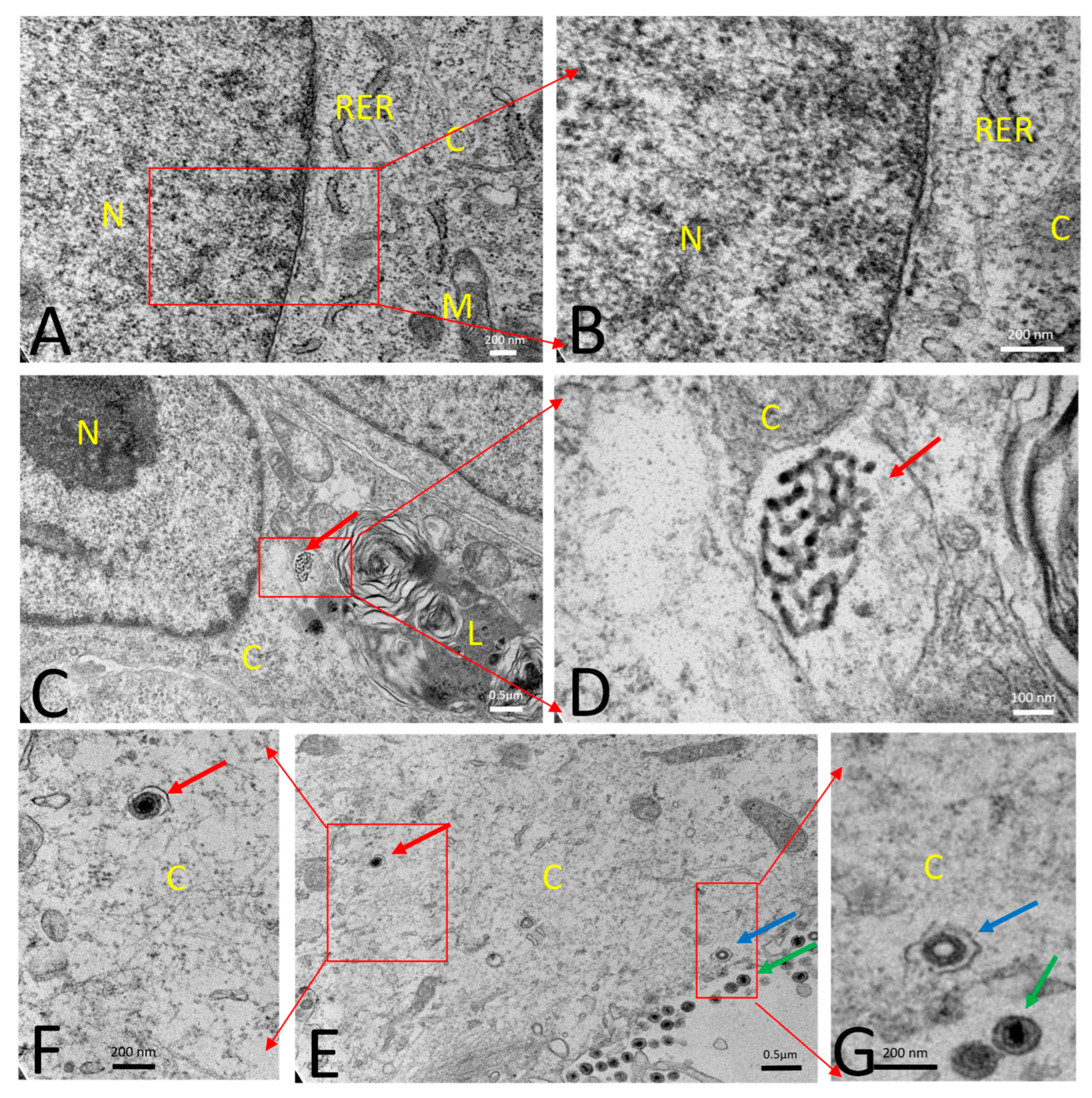
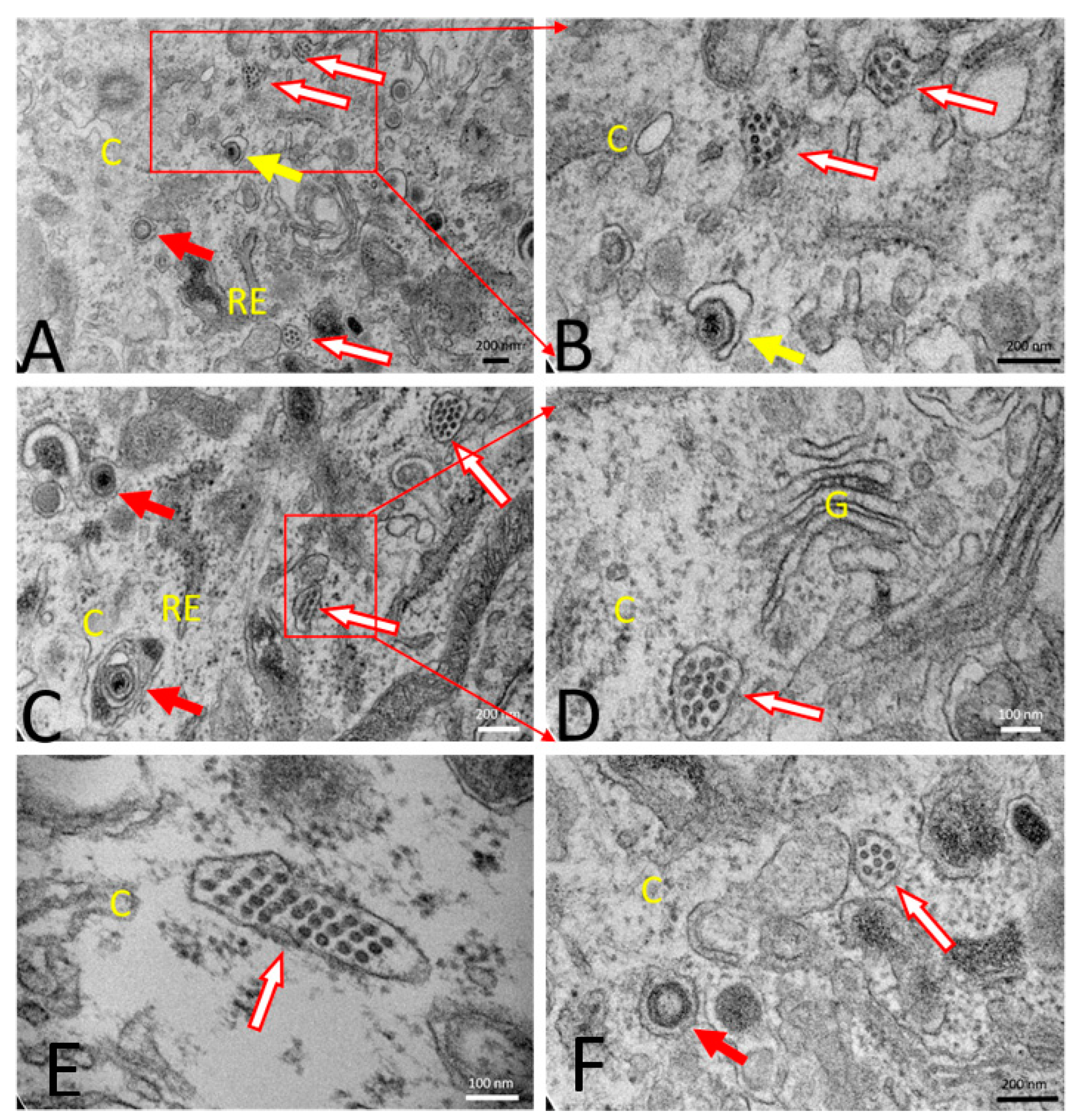
2.13. Transmission Electron Microscopy (TEM)
2.14. Histopathology
2.15. Statistical Analysis
3. Results
3.1. Characterization of PRVtmv+
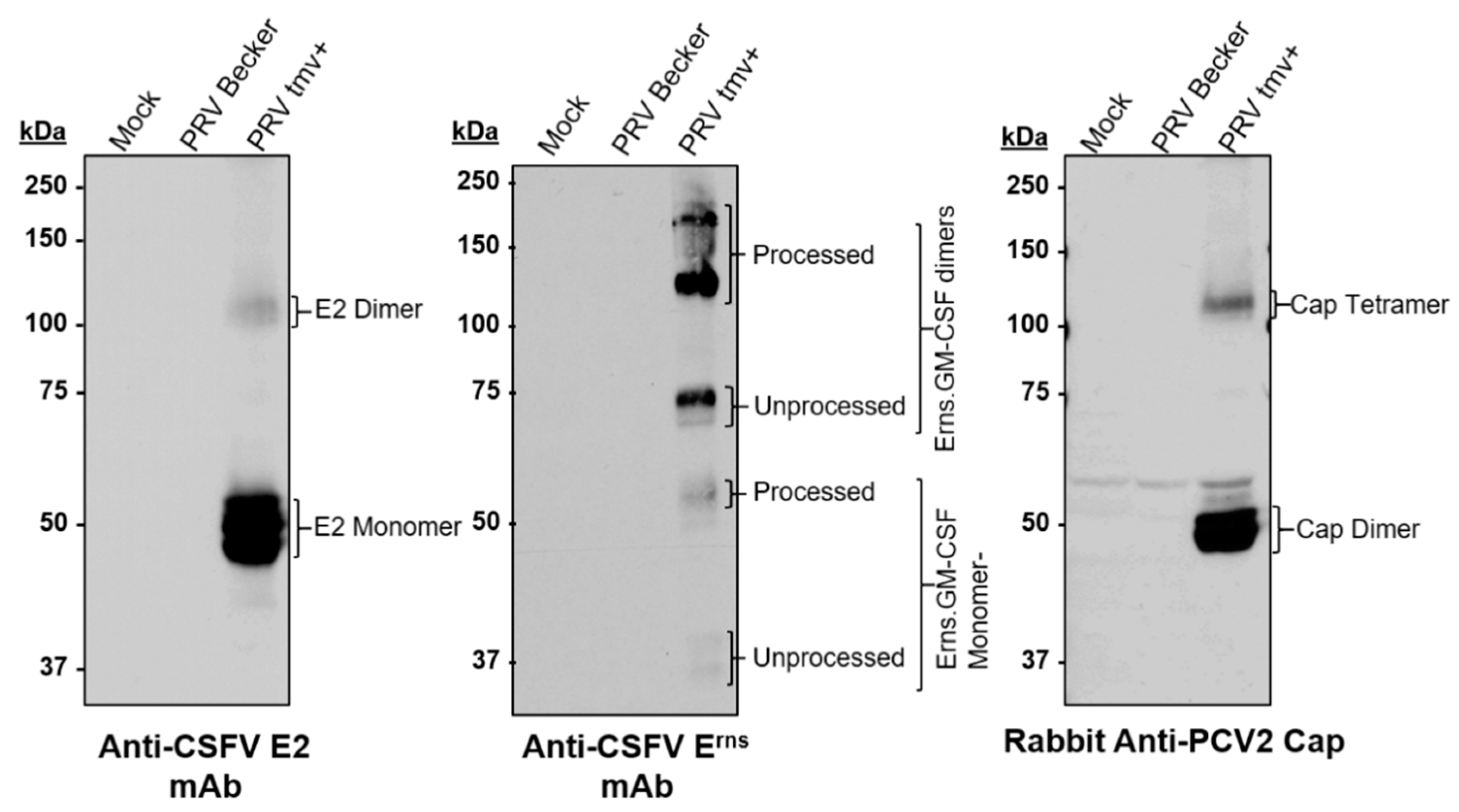
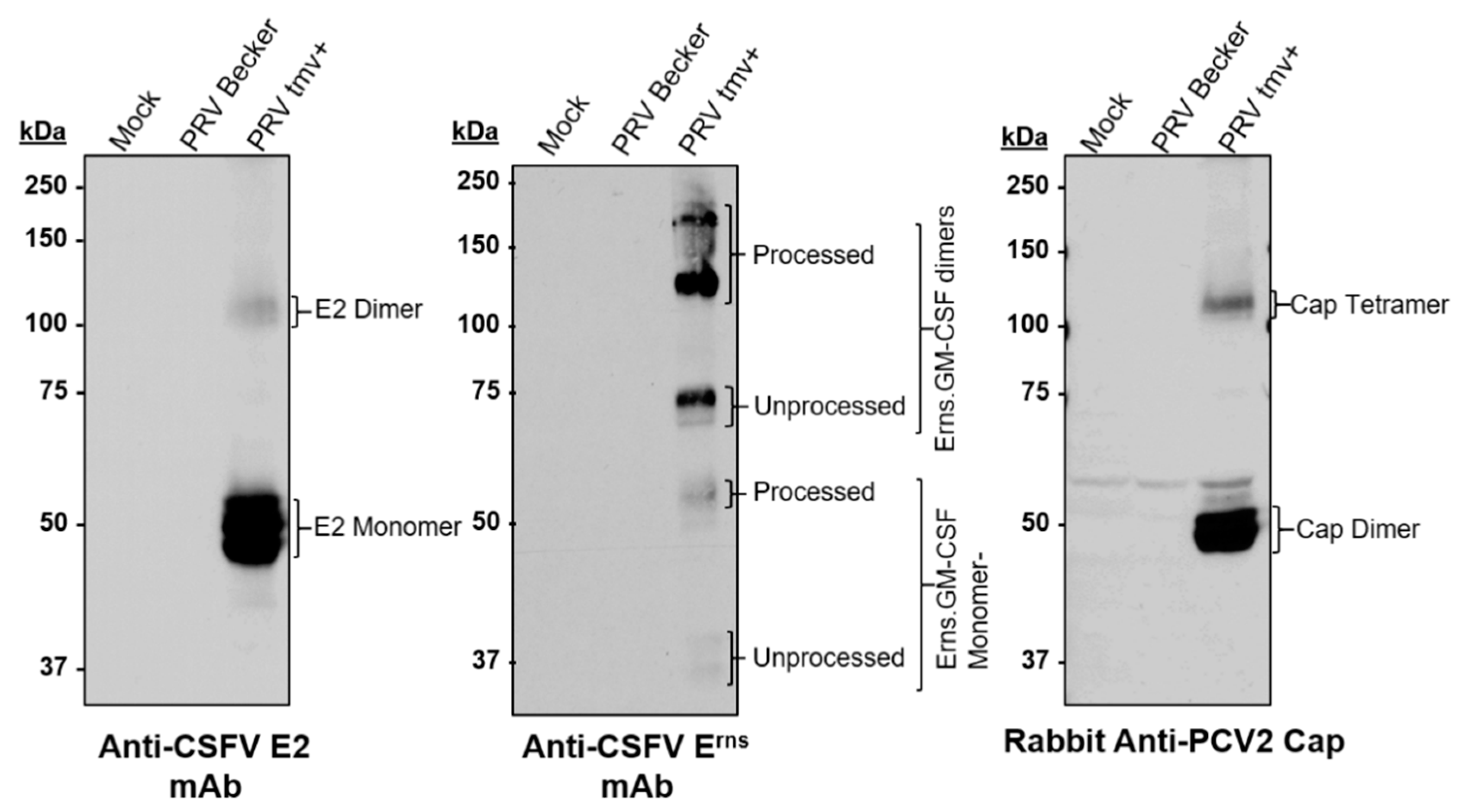
3.1.1. Characterization of PRVtmv+ for the Expression of the CSFV E2, PCV2b Cap, and Chimeric CSFV Erns-GM-CSF
3.1.2. PRVtmv+ Expressed PCV2b Cap Self-Assembles into Virus-like Particles (VLPs) In Vitro
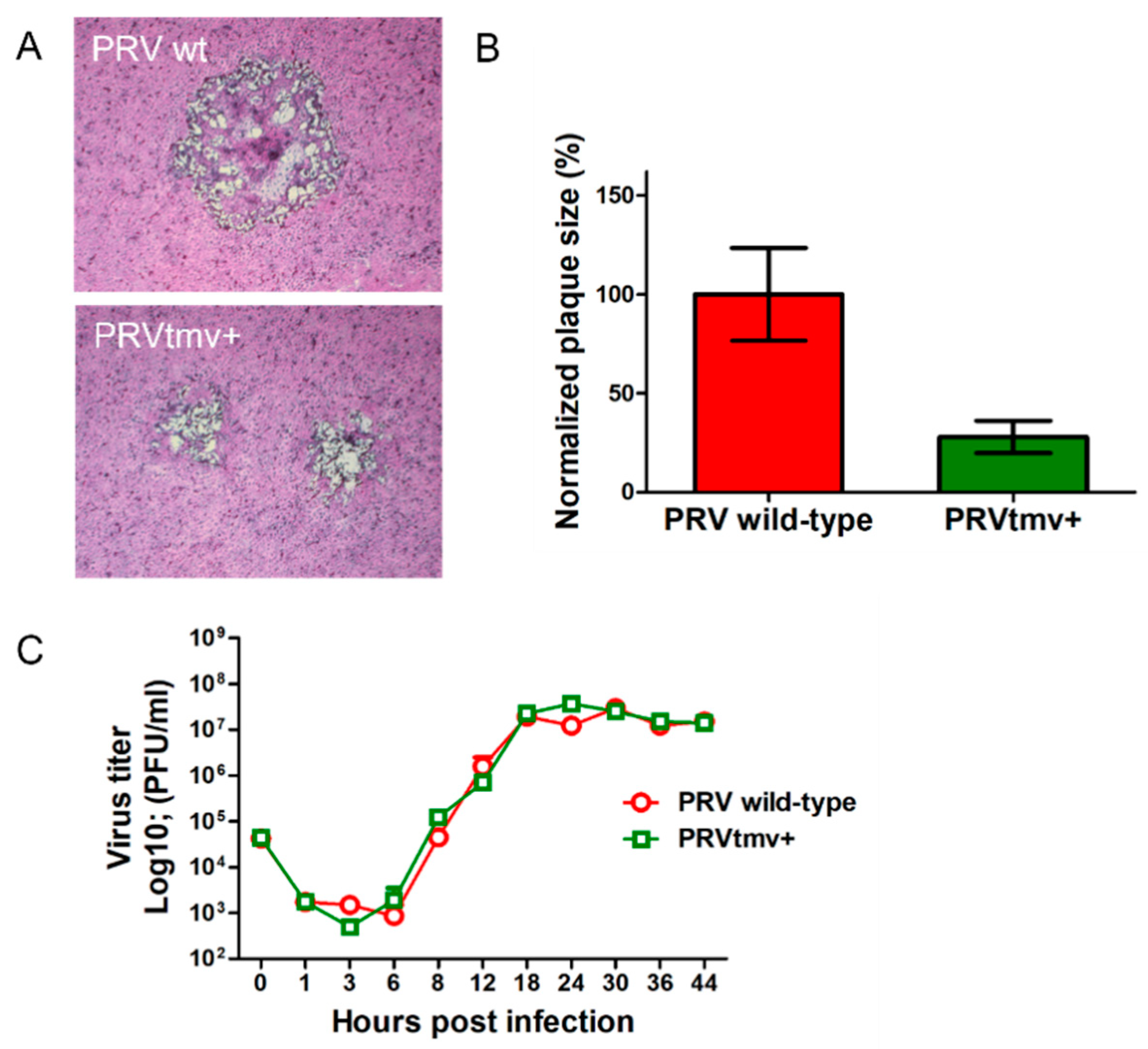
3.2. PRVtmv+ Vaccine Virus Replicates with a Similar Kinetics and Virus Yield in SK Cells, In Vitro, Like the PRV wt but Produces Smaller Plaques
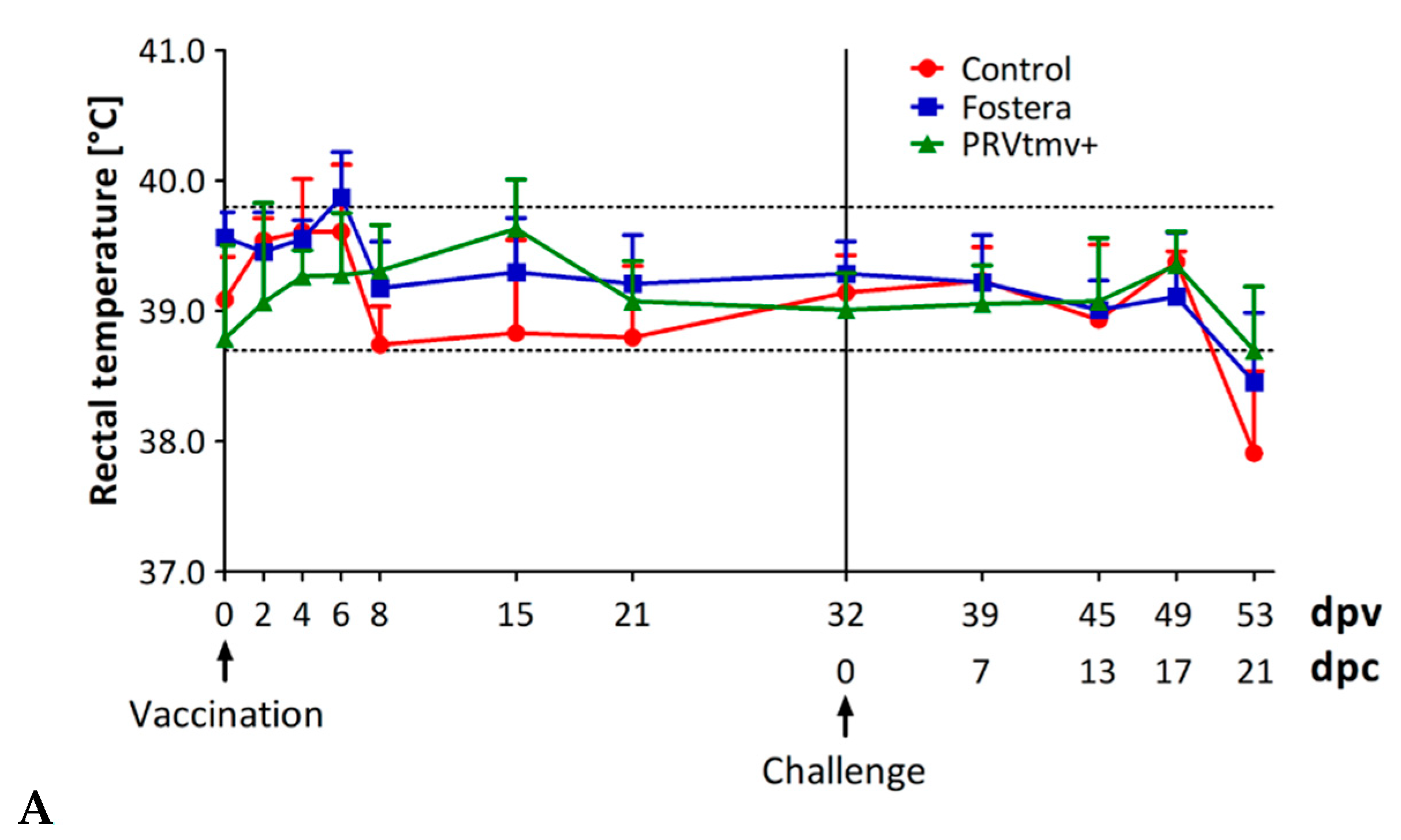
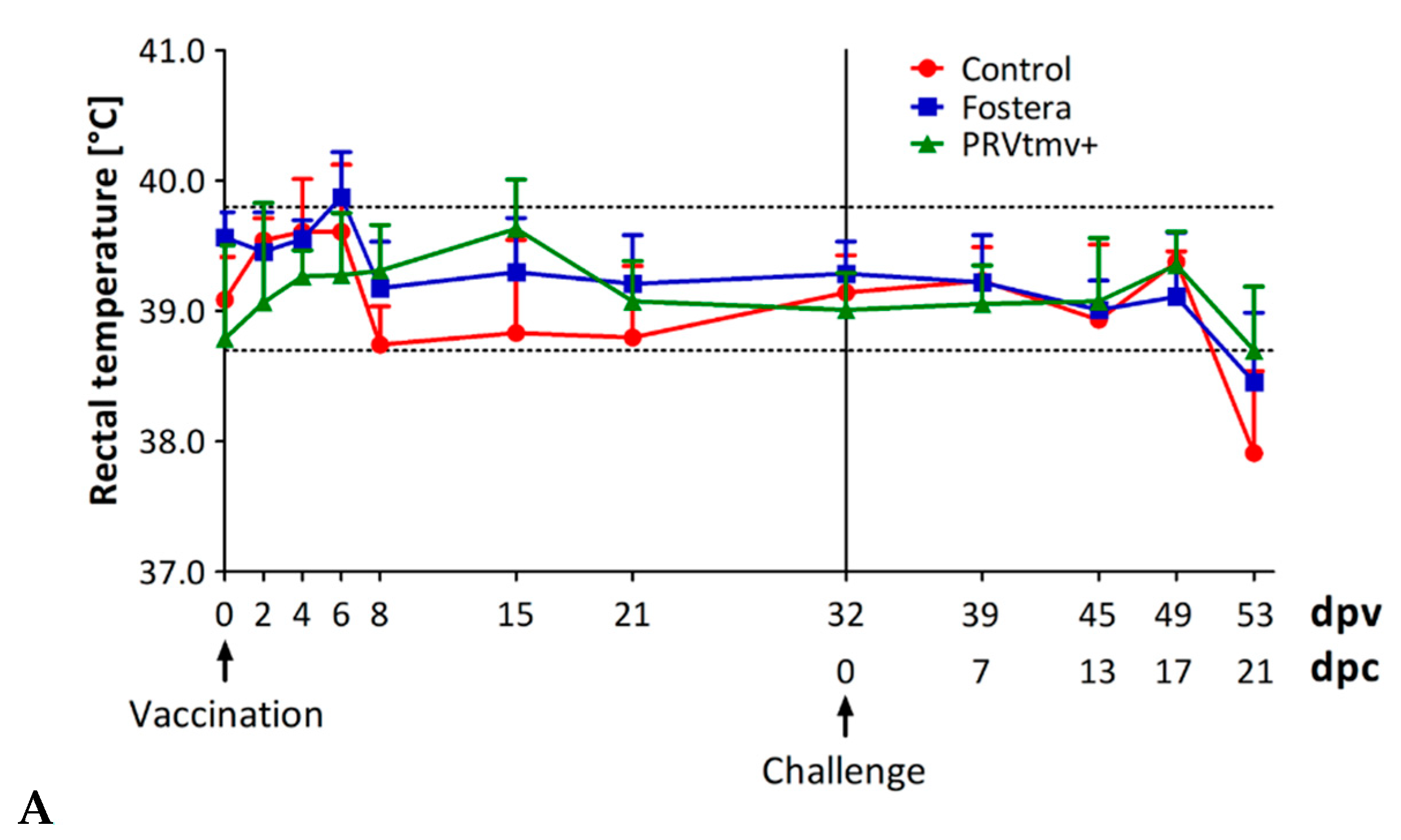
3.3. PRVtmv+ Vaccine Virus Is Highly Attenuated, Safe, and Retains Its Stability in Pigs to Express the PCV2b and CSFV Chimeric Genes
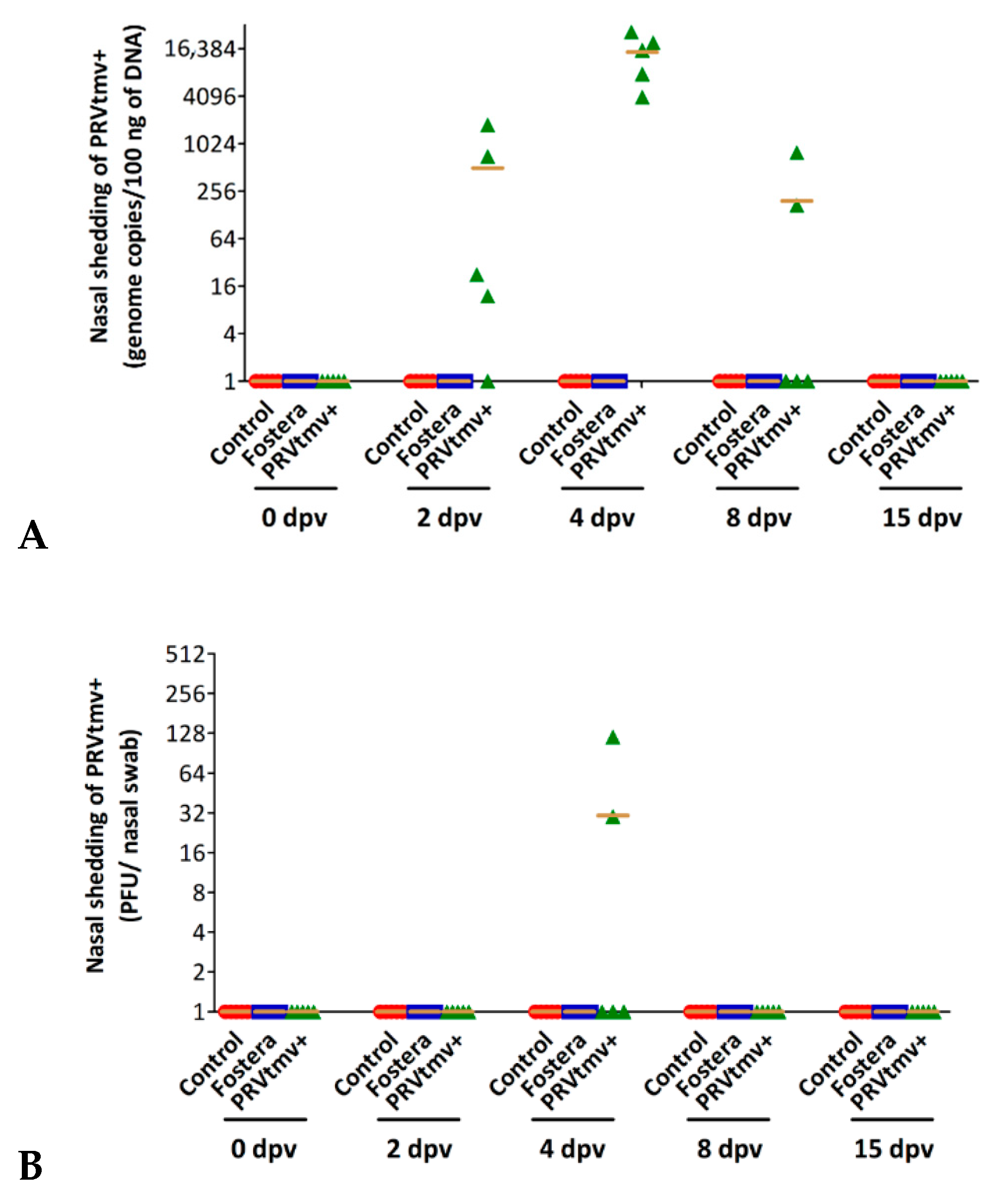
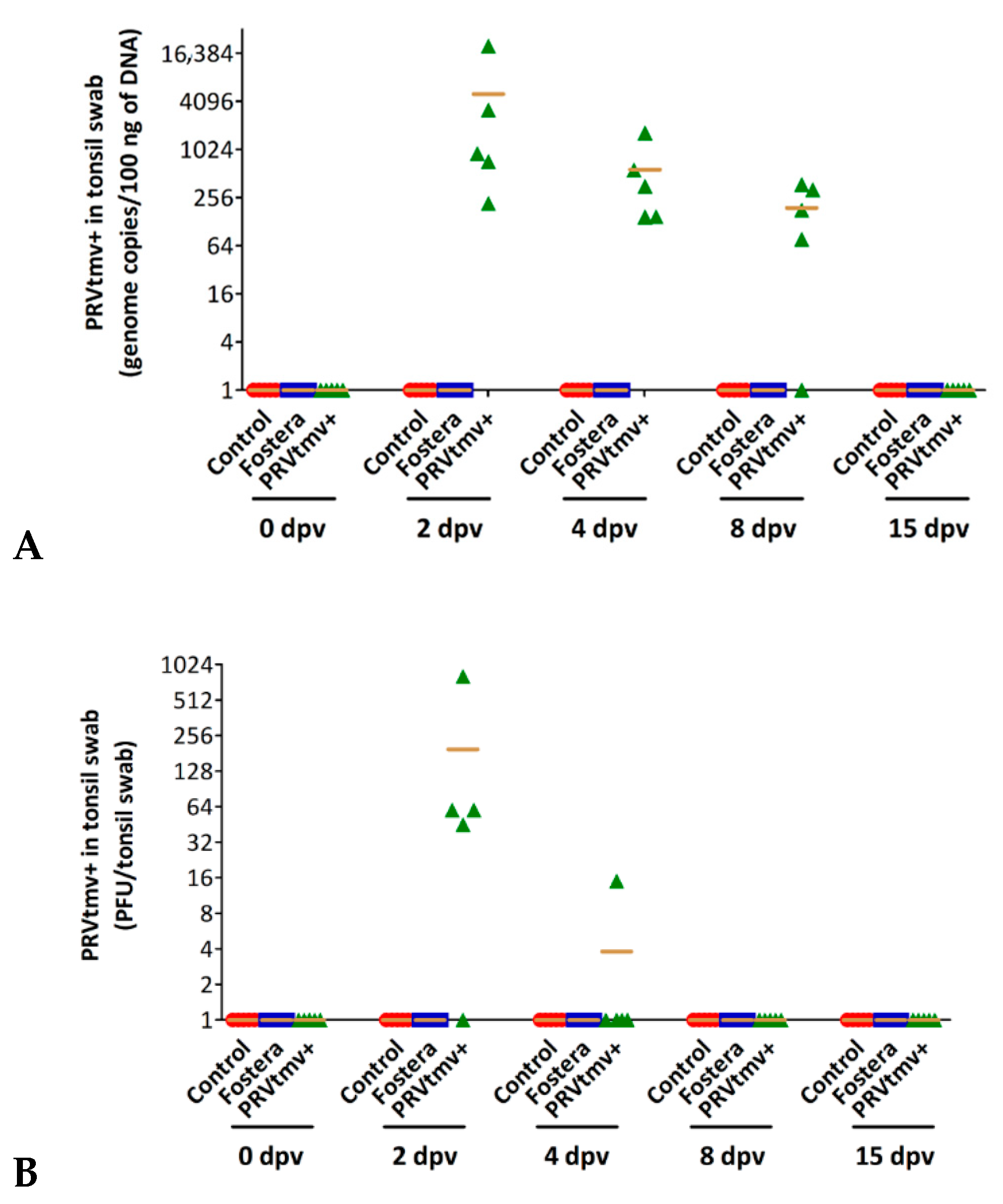
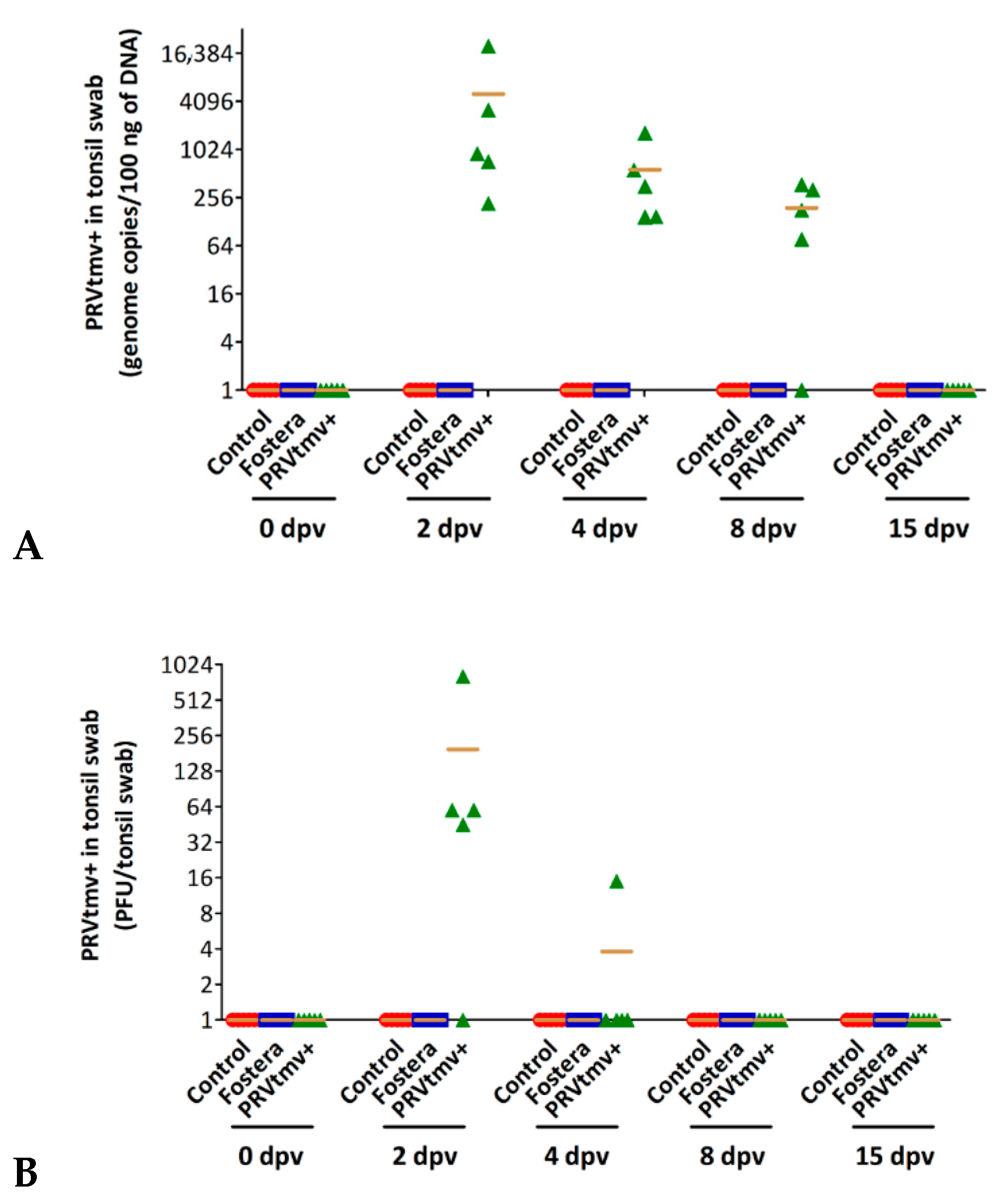
3.4. Nasal Virus Shedding following IN/Subcut Vaccination with PRVtmv+ Vaccine
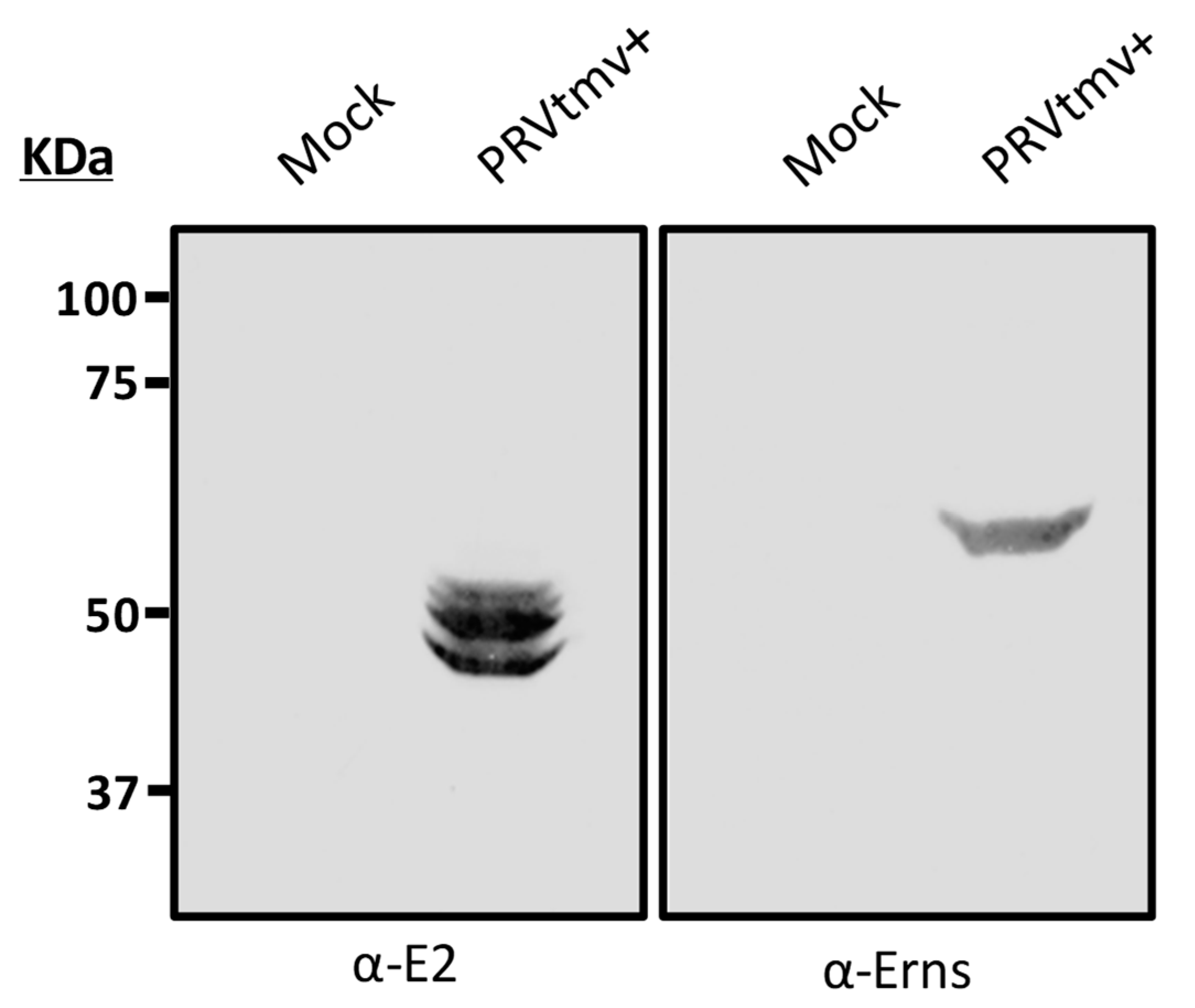
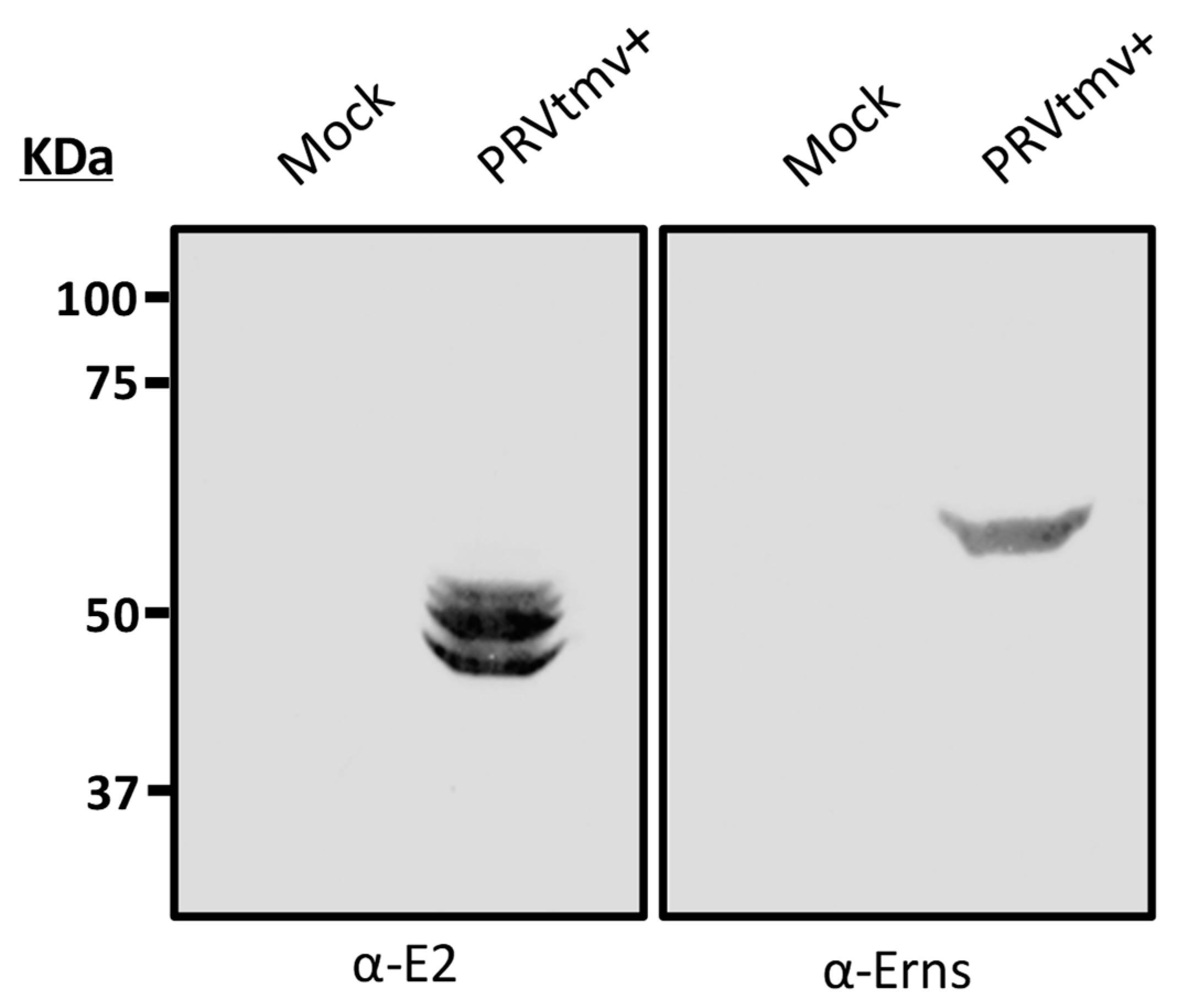
3.5. The Chimeric E2 and Erns Proteins Are Intact in the PRVtmv+ Virus Isolated from the Vaccinated Animals at 4 dpv
3.6. PRVtmv+ Vaccinated Pigs Induced PC2b-Specific Antibodies in the Vaccinated Pigs, Which Detected PCV2b-Infected Cells in Culture
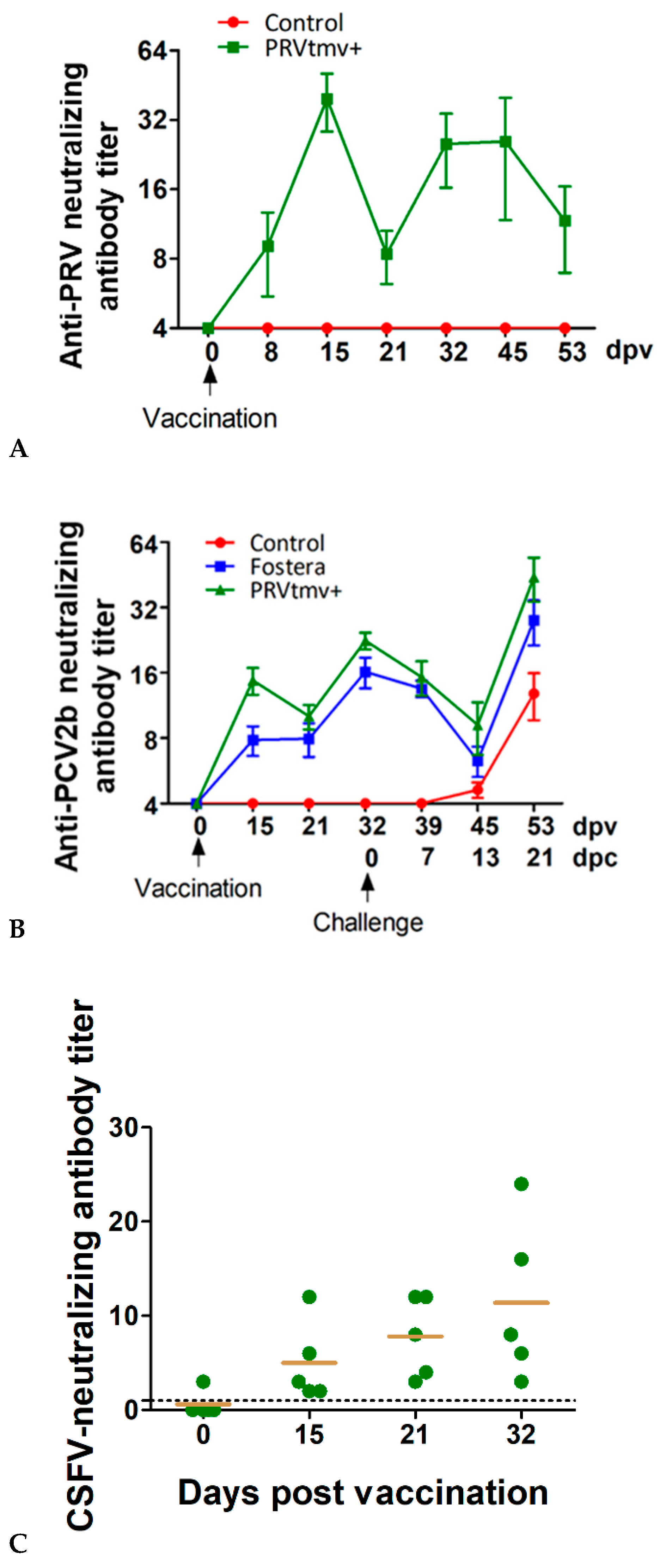
3.7. PRVtmv+ Vaccine Stimulated a Reasonable PRV-Specific Neutralizing Antibody Response in the Vaccinated Pigs
3.8. A Single Dose of PRVtmv+ Vaccine Is Sufficient to Elicit a Higher PCV2b-Specific Antibody Response Than the Inactivated Fostera Gold PCV Vaccine
3.9. PRVtmv+ Immunized Pigs Generated CSFV-Specific Neutralizing Antibody Titers
3.10. Both the PRVtmv+ and the Fostera Vaccinated Pigs Had Similar PCV2b-Specific seroconversion after the Challenge, but the SN Antibody Titers in the PRVtmv+ Vaccine Group Were Two-Fold Higher than the Commercial Vaccine Group
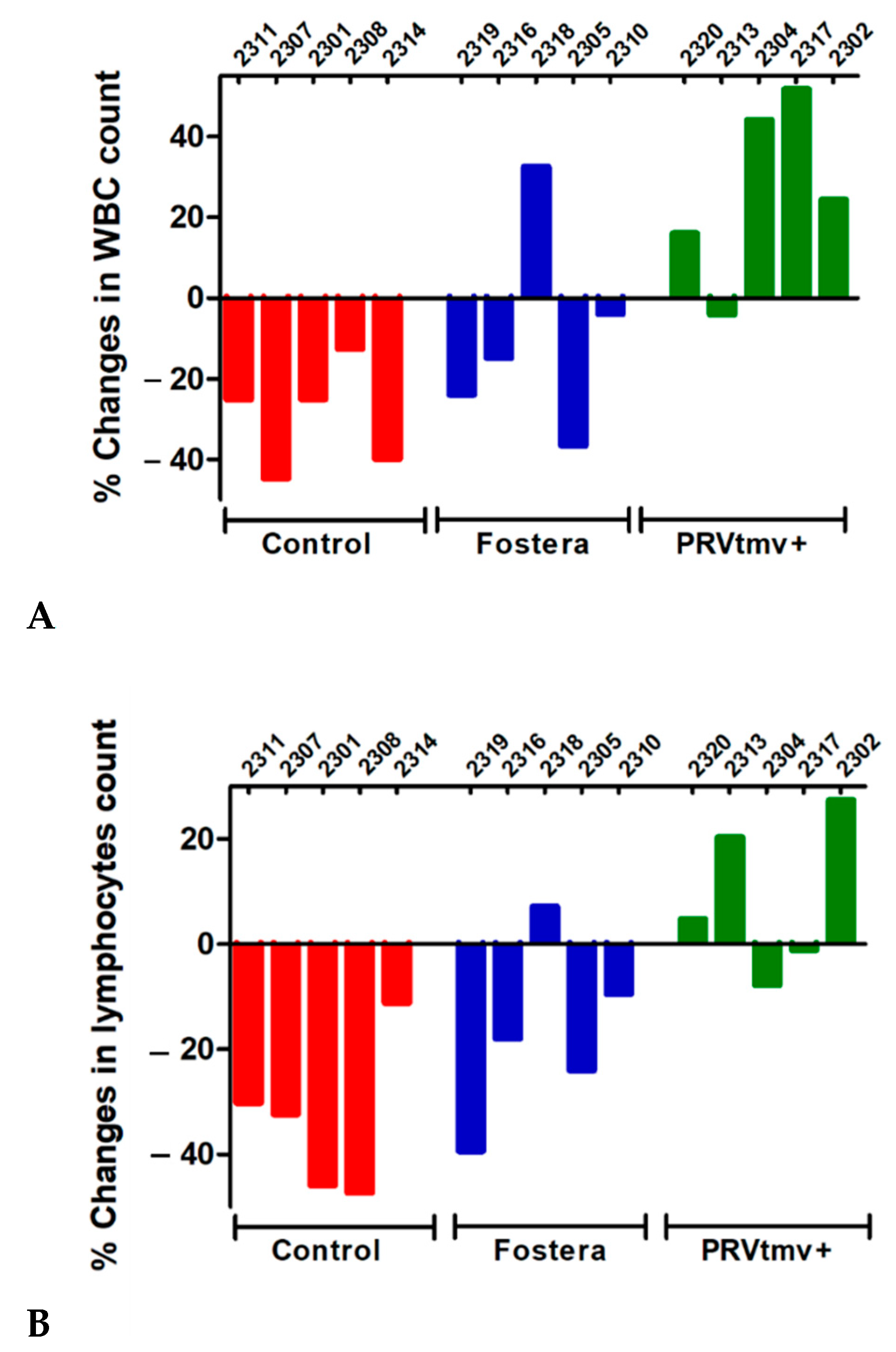
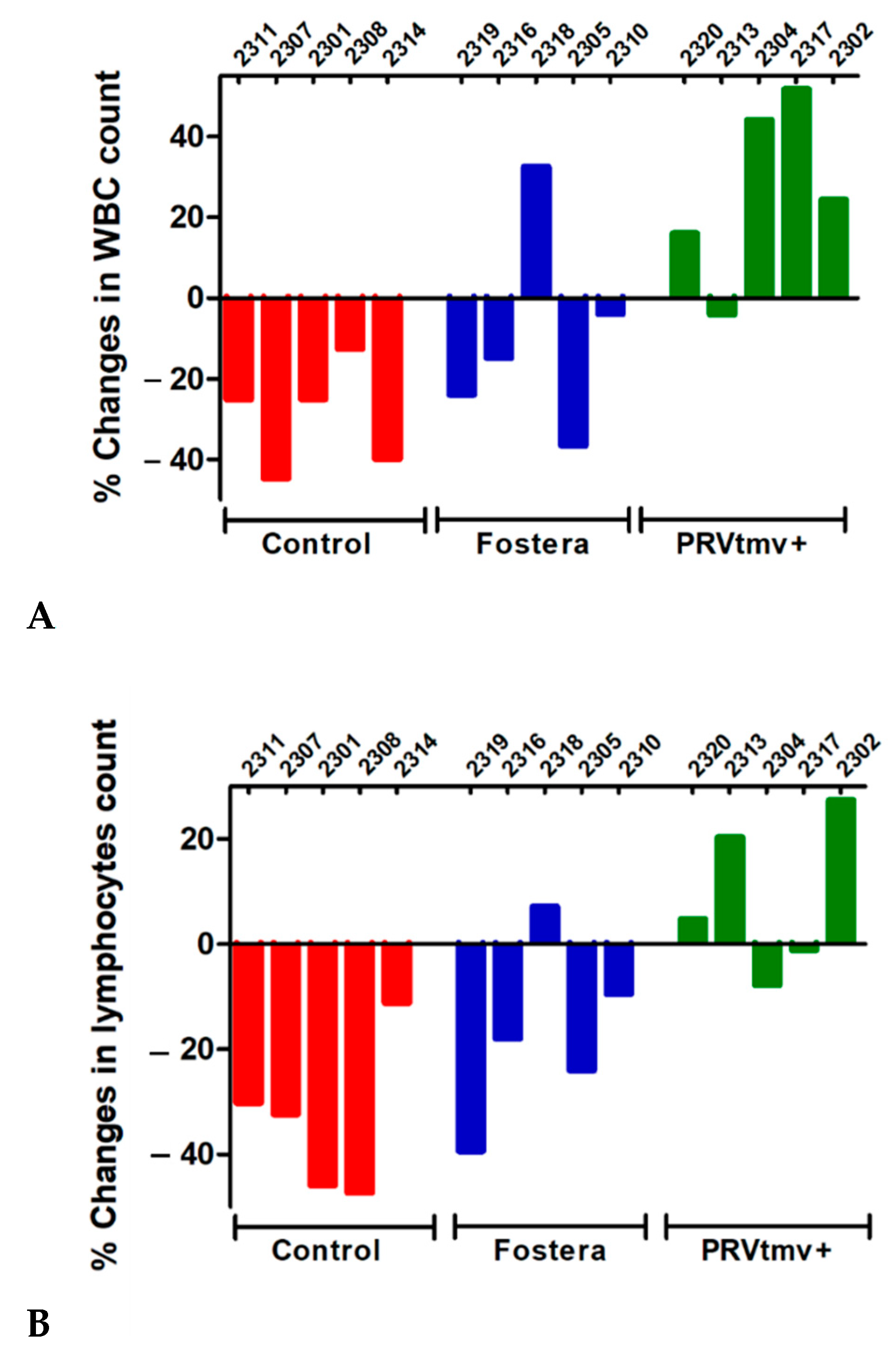
3.11. While PRVtmv+ Vaccinated Pigs Had a Moderate Increase in Leukocyte and Lymphocyte Counts following the PCV2b Challenge, Pigs in the Control Unvaccinated and Commercial, Fostera Vaccine Groups Had a Reduction in Both the Counts (a Moderate to Low Level of Leukopenia and Lymphopenia)
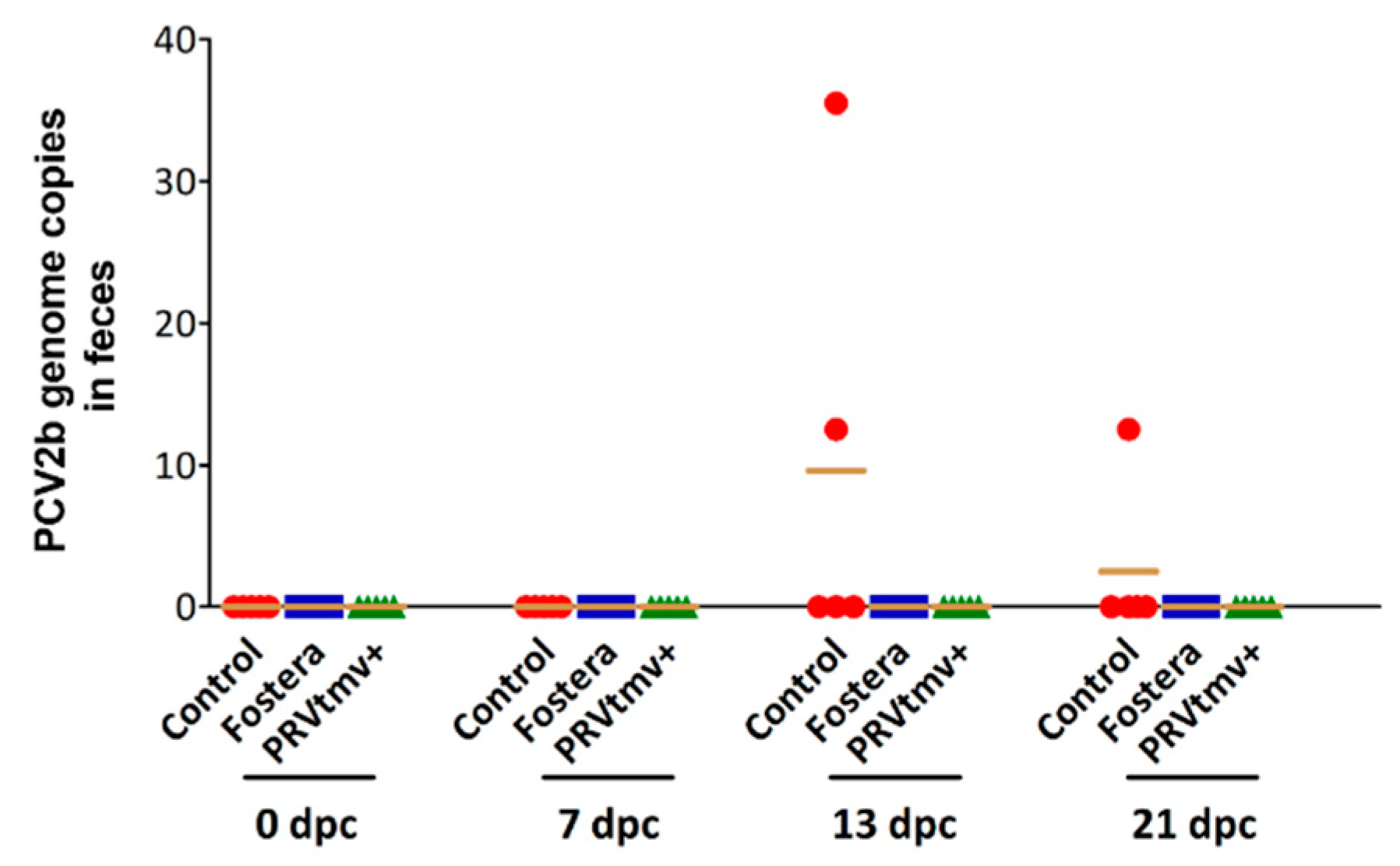
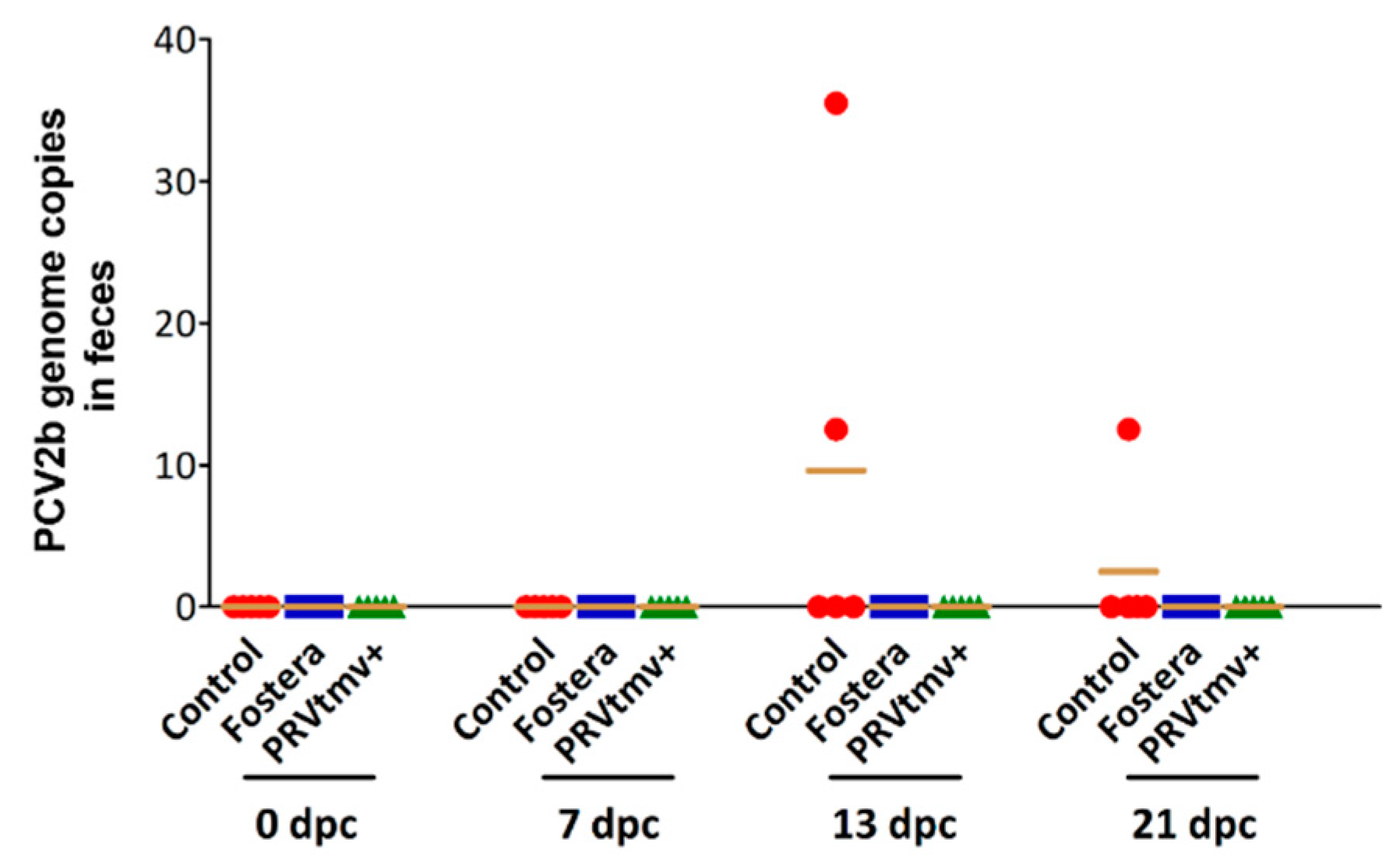
3.12. Both PRVtmv+ and Fostera Immunized Pigs Did Not Shed the PCV2b Challenge Virus in Feces
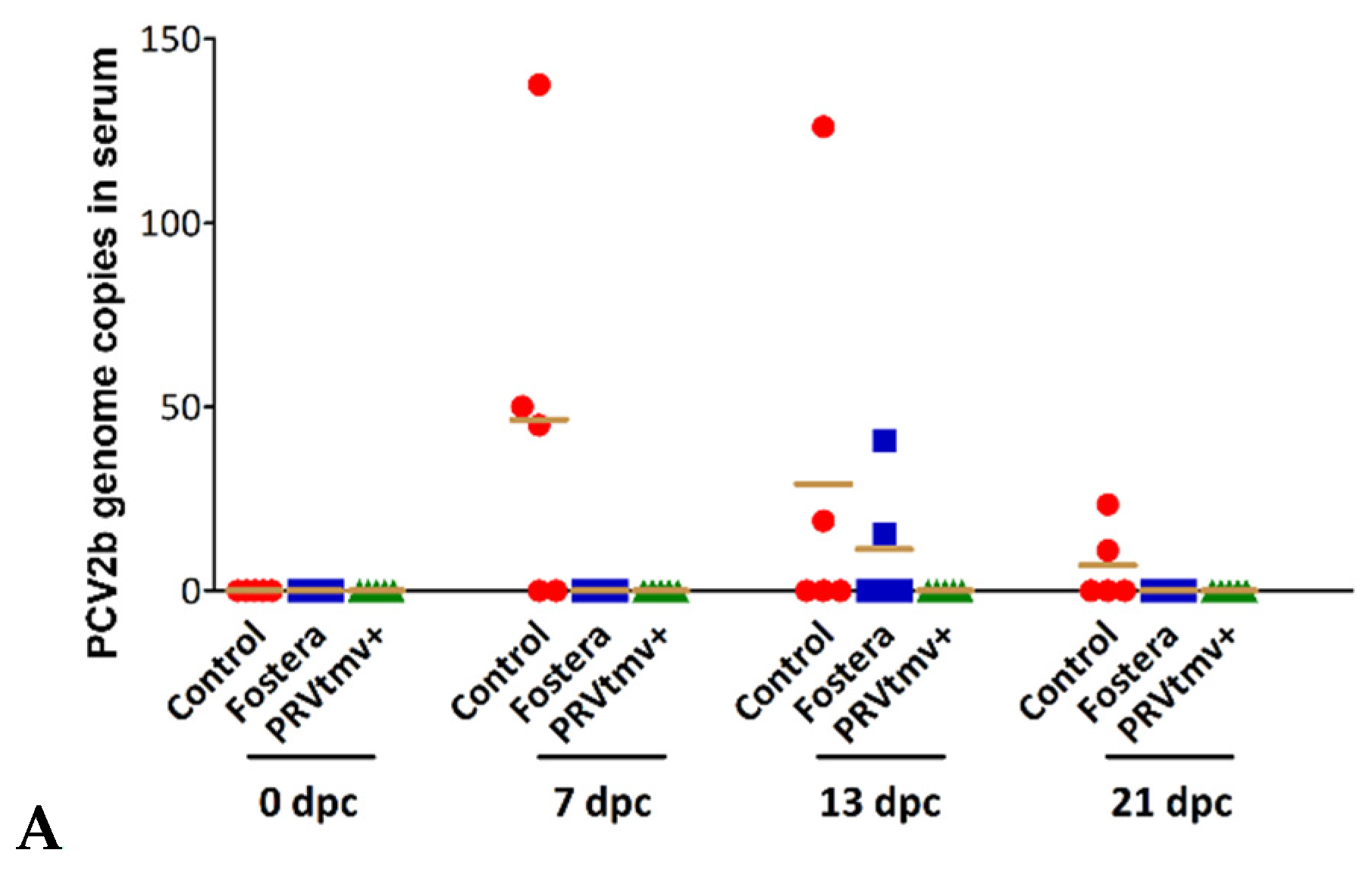
3.13. PRVtmv+ Protected the Vaccinated Pigs from Both Cell-Free and Cell-Associated Viremia after PCV2b Challenge
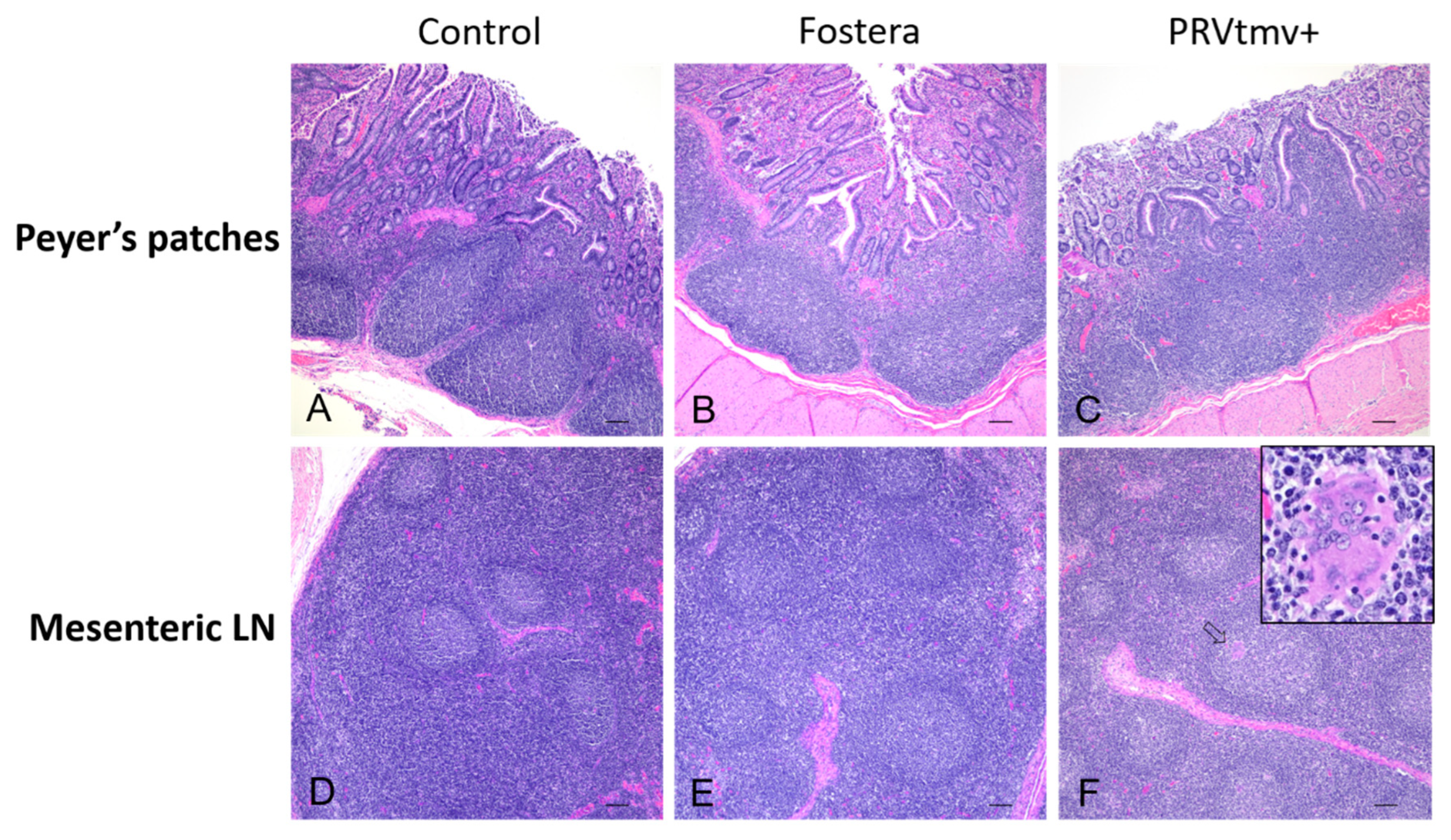
3.14. Gross and Histopathology Lesions in Pigs
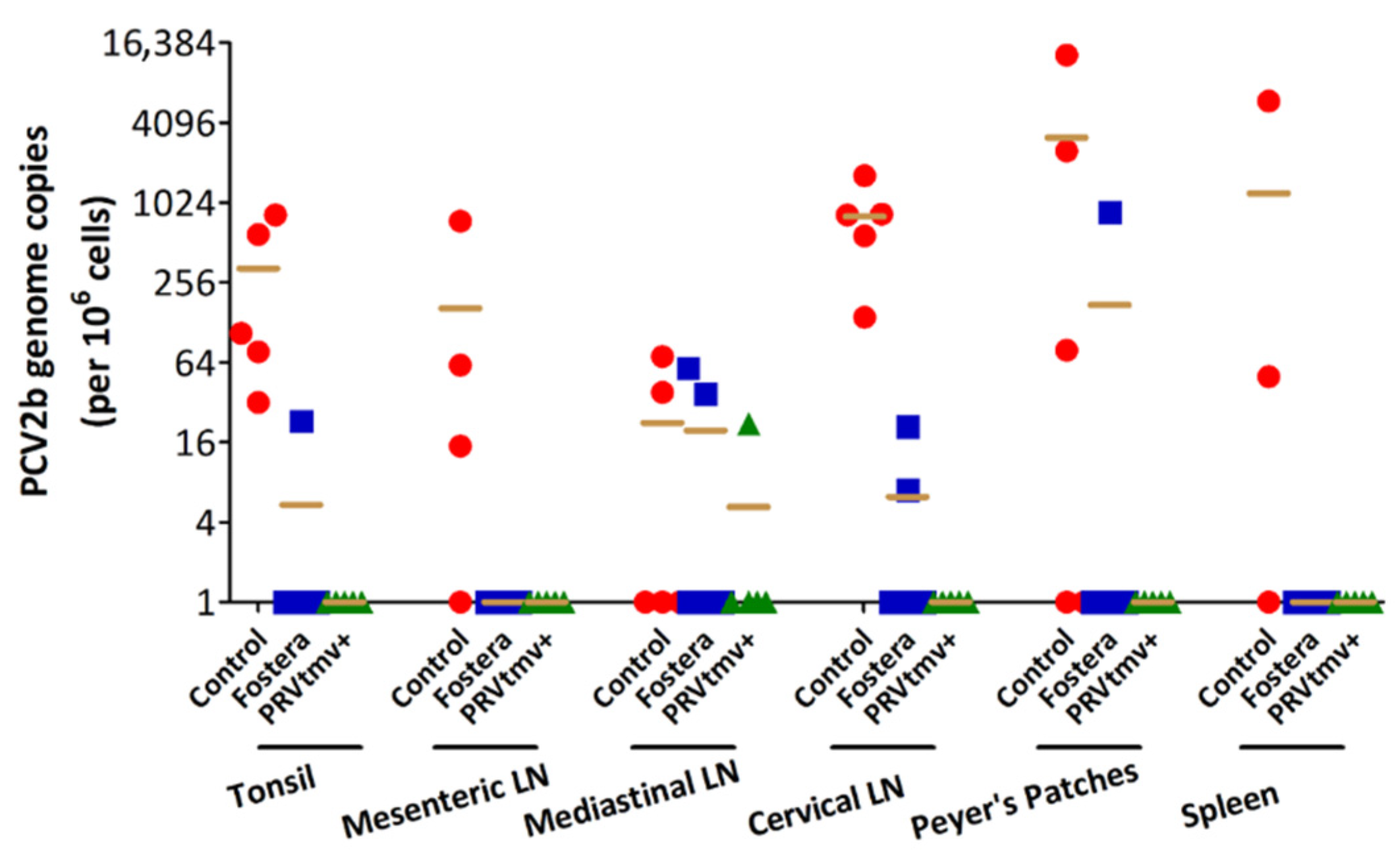
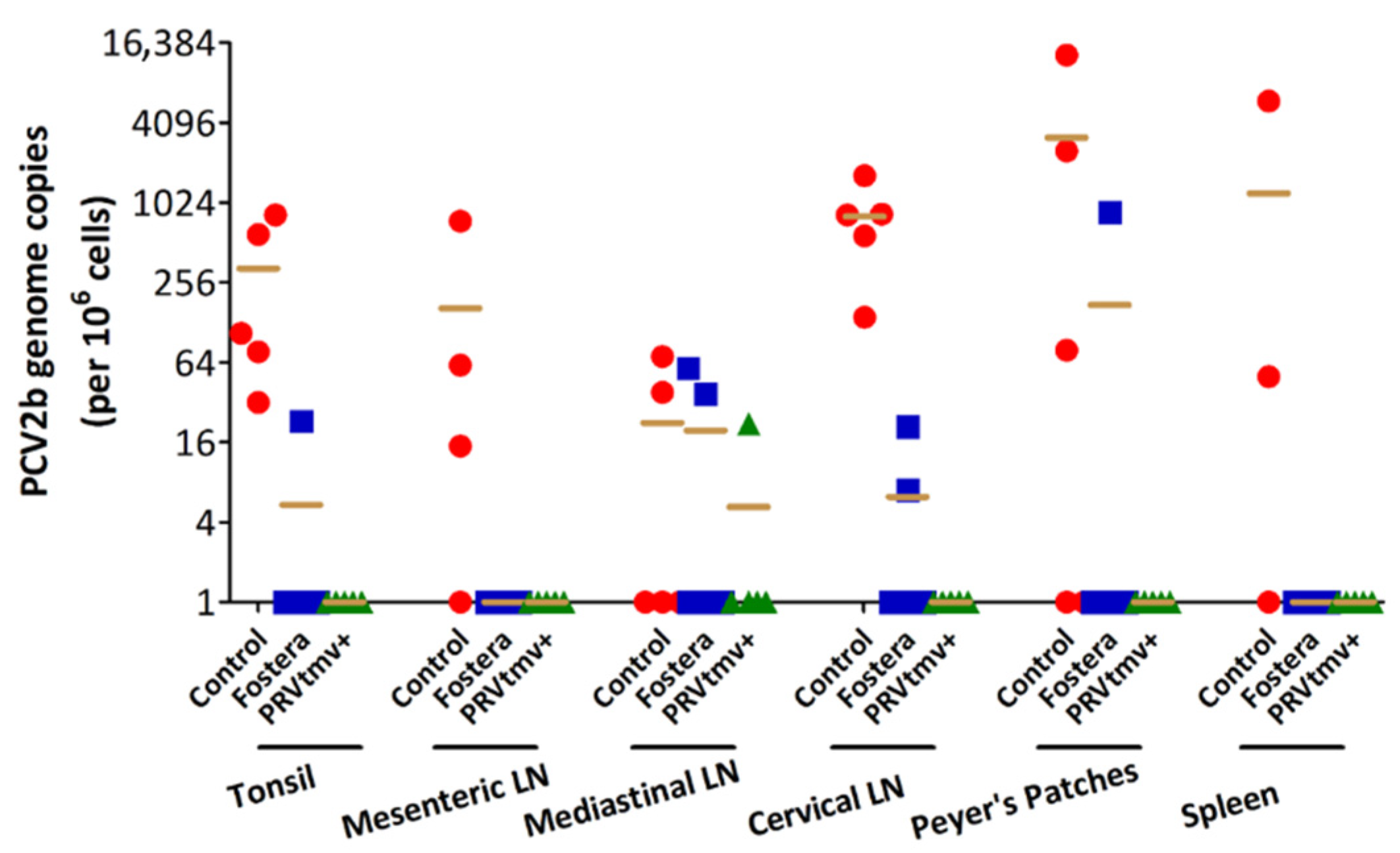
3.15. PRVtmv+ Vaccine Protected Pigs against PCV2b Challenge Better than the Inactivated Commercial Vaccine Fostera Based on Viral Load in Lymph Nodes
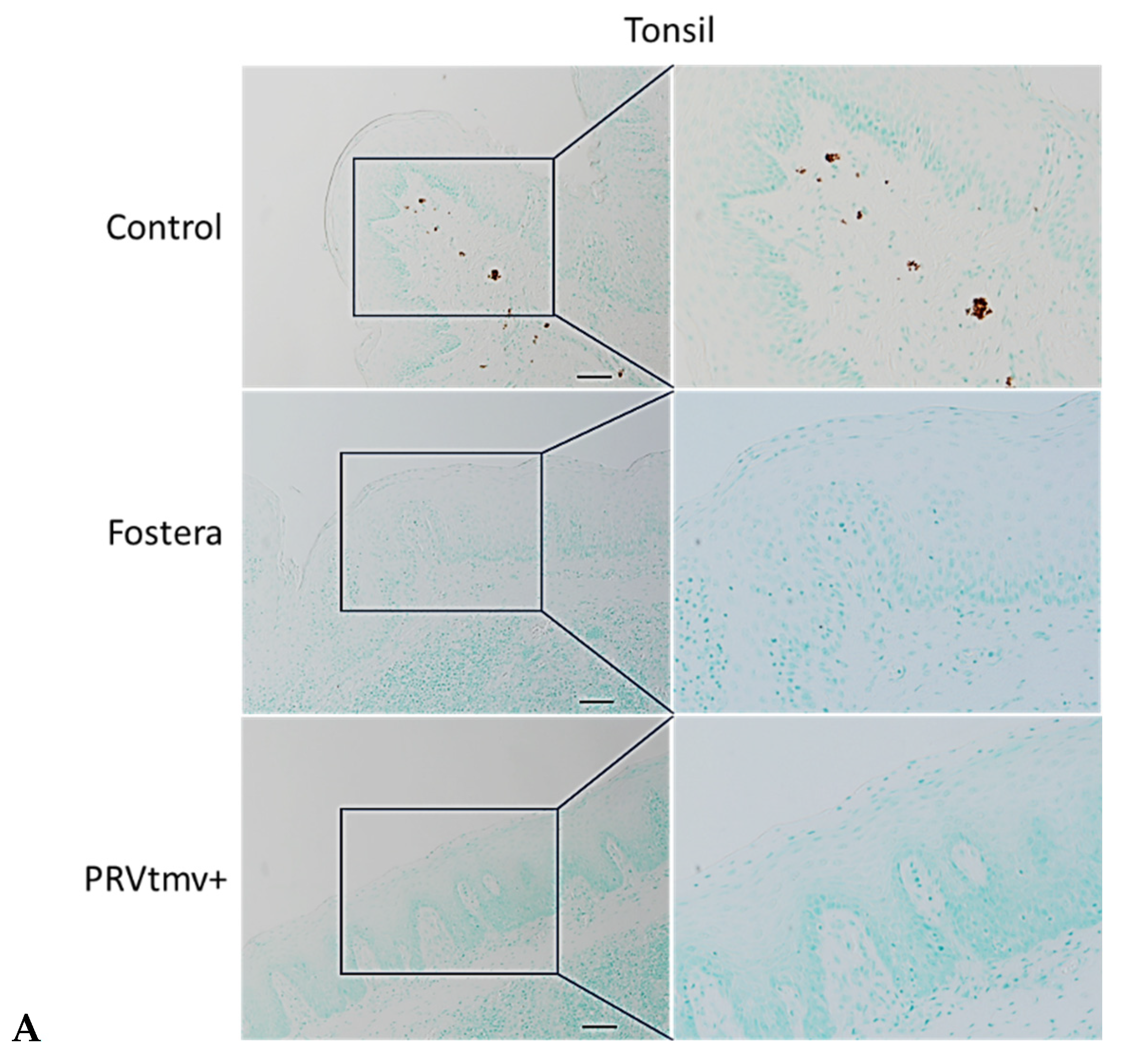
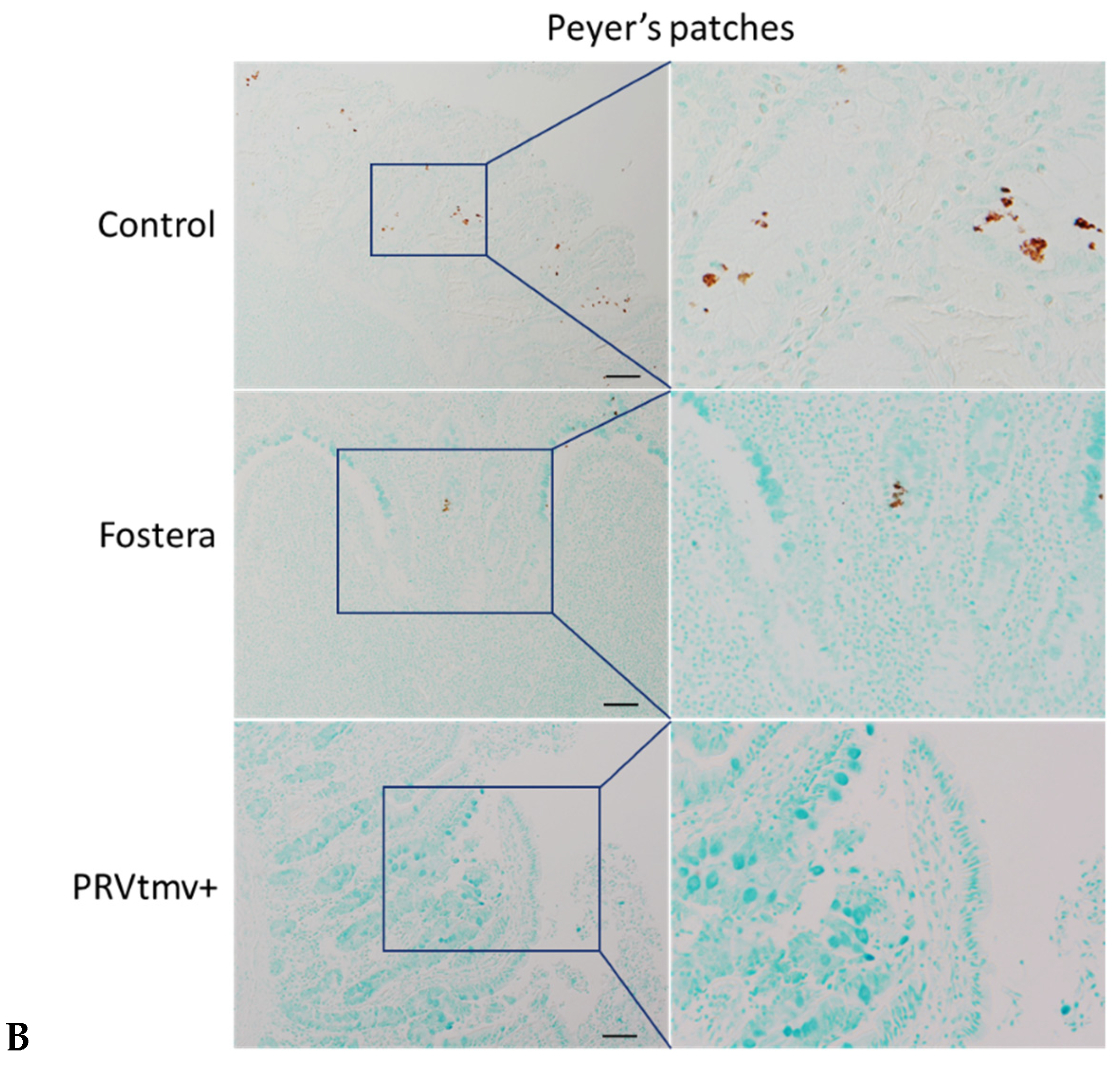
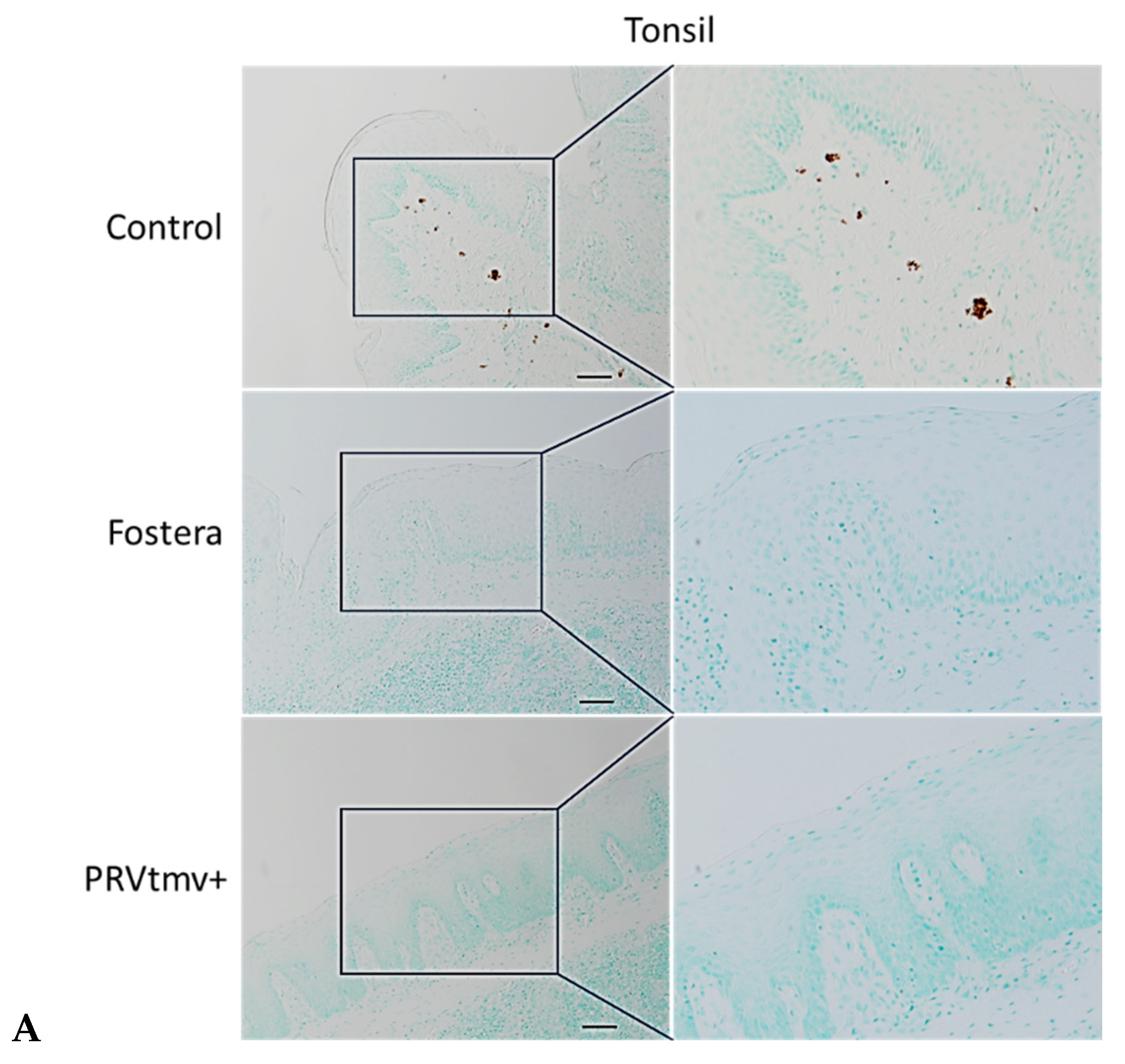
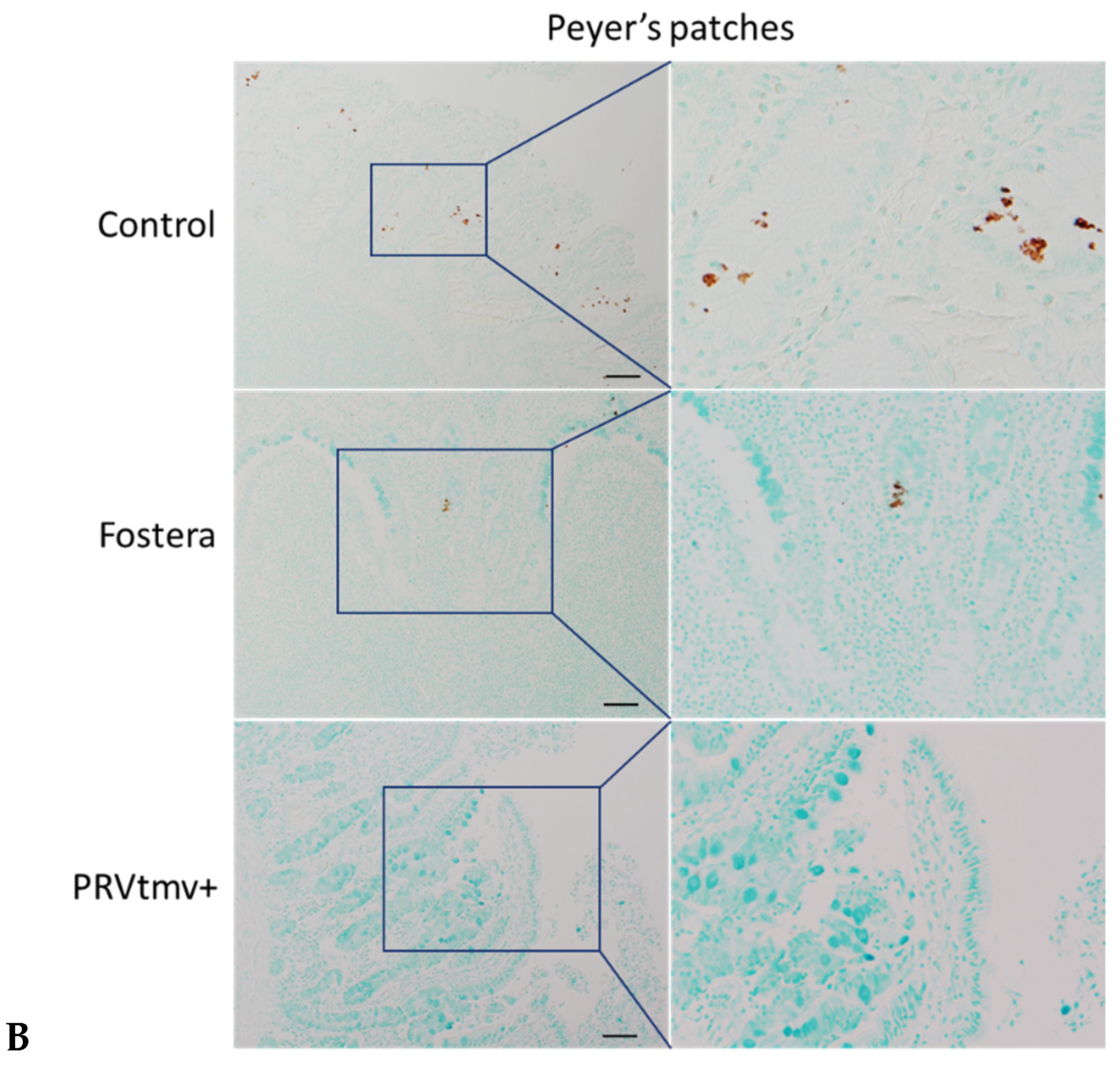
3.16. With the Exception of One Pig in the Fostera Group, Both PRVtmv+ and Fostera Vaccinated Pigs Were Negative for PCV2b Cap-Specific Antigens in the Lymphoid Tissues
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Baekbo, P.; Kristensen, C.S.; Larsen, L.E. Porcine circovirus diseases: A review of PMWS. Transbound. Emerg. Dis. 2012, 59 (Suppl. S1), 60–67. [Google Scholar] [CrossRef] [PubMed]
- Allan, G.M.; McNeilly, F.; Kennedy, S.; Daft, B.; Clarke, E.G.; Ellis, J.A.; Haines, D.M.; Meehan, B.M.; Adair, B.M. Isolation of porcine circovirus-like viruses from pigs with a wasting disease in the USA and Europe. J. Vet. Diagn. Investig. 1998, 10, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.J. Porcine circovirus type 2 (PCV2): Pathogenesis and interaction with the immune system. Annu. Rev. Anim. Biosci. 2013, 1, 43–64. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.A.; Bratanich, A.; Clark, E.G.; Allan, G.; Meehan, B.; Haines, D.M.; Harding, J.; West, K.H.; Krakowka, S.; Konoby, C.; et al. Coinfection by porcine circoviruses and porcine parvovirus in pigs with naturally acquired postweaning multisystemic wasting syndrome. J. Vet. Diagn. Investig. 2000, 12, 21–27. [Google Scholar] [CrossRef]
- Huang, Y.L.; Pang, V.F.; Lin, C.M.; Tsai, Y.C.; Chia, M.Y.; Deng, M.C.; Chang, C.Y.; Jeng, C.R. Porcine circovirus type 2 (PCV2) infection decreases the efficacy of an attenuated classical swine fever virus (CSFV) vaccine. Vet. Res. 2011, 42, 115. [Google Scholar] [CrossRef] [Green Version]
- Allan, G.M.; Ellis, J.A. Porcine circoviruses: A review. J. Vet. Diagn. Investig. 2000, 12, 3–14. [Google Scholar] [CrossRef]
- Edwards, S.; Fukusho, A.; Lefevre, P.C.; Lipowski, A.; Pejsak, Z.; Roehe, P.; Westergaard, J. Classical swine fever: The global situation. Vet. Microbiol. 2000, 73, 103–119. [Google Scholar] [CrossRef]
- OIE. WAHID Home Page. Available online: https://www.oie.int/en/disease/classical-swine-fever/#:~:text=Geographical%20distribution,currently%20free%20of%20the%20disease (accessed on 24 October 2021).
- Postel, A.; Austermann-Busch, S.; Petrov, A.; Moennig, V.; Becher, P. Epidemiology, diagnosis and control of classical swine fever: Recent developments and future challenges. Transbound. Emerg. Dis. 2018, 65 (Suppl. S1), 248–261. [Google Scholar] [CrossRef] [Green Version]
- Stegeman, A.; Elbers, A.; de Smit, H.; Moser, H.; Smak, J.; Pluimers, F. The 1997–1998 epidemic of classical swine fever in the Netherlands. Vet. Microbiol. 2000, 73, 183–196. [Google Scholar] [CrossRef]
- Coronado, L.; Perera, C.L.; Rios, L.; Frias, M.T.; Perez, L.J. A Critical Review about Different Vaccines against Classical Swine Fever Virus and Their Repercussions in Endemic Regions. Vaccines 2021, 9, 154. [Google Scholar] [CrossRef]
- Mettenleiter, T.C. Aujeszky’s disease (pseudorabies) virus: The virus and molecular pathogenesis—State of the art, June 1999. Vet. Res. 2000, 31, 99–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maresch, C.; Lange, E.; Teifke, J.P.; Fuchs, W.; Klupp, B.; Muller, T.; Mettenleiter, T.C.; Vahlenkamp, T.W. Oral immunization of wild boar and domestic pigs with attenuated live vaccine protects against Pseudorabies virus infection. Vet. Microbiol. 2012, 161, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.J.; Tian, Z.J.; Tong, G.Z.; Zhou, Y.J.; Ni, J.Q.; Luo, Y.Z.; Cai, X.H. Protective immunity induced by a recombinant pseudorabies virus expressing the GP5 of porcine reproductive and respiratory syndrome virus in piglets. Vet. Immunol. Immunopathol. 2005, 106, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Bo, Z.; Miao, Y.; Xi, R.; Gao, X.; Miao, D.; Chen, H.; Jung, Y.S.; Qian, Y.; Dai, J. Emergence of a novel pathogenic recombinant virus from Bartha vaccine and variant pseudorabies virus in China. Transbound. Emerg. Dis. 2021, 68, 1454–1464. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Bai, C.; Sun, J.; Chang, S.; Zhang, X. Emergence of virulent pseudorabies virus infection in northern China. J. Vet. Sci. 2013, 14, 363–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Pan, Y.; Liu, M.; Han, Z. Prevalence of Porcine Pseudorabies Virus and Its Coinfection Rate in Heilongjiang Province in China from 2013 to 2018. Viral Immunol. 2020, 33, 550–554. [Google Scholar] [CrossRef]
- Sun, Y.; Luo, Y.; Wang, C.H.; Yuan, J.; Li, N.; Song, K.; Qiu, H.J. Control of swine pseudorabies in China: Opportunities and limitations. Vet. Microbiol. 2016, 183, 119–124. [Google Scholar] [CrossRef]
- Blome, S.; Staubach, C.; Henke, J.; Carlson, J.; Beer, M. Classical Swine Fever-An Updated Review. Viruses 2017, 9, 86. [Google Scholar] [CrossRef] [Green Version]
- USDA-APHIS. Procedure Manual for Classical Swine Fever (CSF) Surveillance. Available online: https://www.aphis.usda.gov/vs/nahss/swine/csf/CSF_procedure_manual_2007.pdf (accessed on 24 October 2021).
- Hamel, A.L.; Lin, L.L.; Nayar, G.P. Nucleotide sequence of porcine circovirus associated with postweaning multisystemic wasting syndrome in pigs. J. Virol. 1998, 72, 5262–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firth, C.; Charleston, M.A.; Duffy, S.; Shapiro, B.; Holmes, E.C. Insights into the evolutionary history of an emerging livestock pathogen: Porcine circovirus 2. J. Virol. 2009, 83, 12813–12821. [Google Scholar] [CrossRef] [Green Version]
- Karuppannan, A.K.; Opriessnig, T. Porcine Circovirus Type 2 (PCV2) Vaccines in the Context of Current Molecular Epidemiology. Viruses 2017, 9, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahe, D.; Blanchard, P.; Truong, C.; Arnauld, C.; Le Cann, P.; Cariolet, R.; Madec, F.; Albina, E.; Jestin, A. Differential recognition of ORF2 protein from type 1 and type 2 porcine circoviruses and identification of immunorelevant epitopes. J. Gen. Virol. 2000, 81, 1815–1824. [Google Scholar] [CrossRef] [PubMed]
- Pogranichnyy, R.M.; Yoon, K.J.; Harms, P.A.; Swenson, S.L.; Zimmerman, J.J.; Sorden, S.D. Characterization of immune response of young pigs to porcine circovirus type 2 infection. Viral Immunol. 2000, 13, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Becher, P.; Avalos Ramirez, R.; Orlich, M.; Cedillo Rosales, S.; Konig, M.; Schweizer, M.; Stalder, H.; Schirrmeier, H.; Thiel, H.J. Genetic and antigenic characterization of novel pestivirus genotypes: Implications for classification. Virology 2003, 311, 96–104. [Google Scholar] [CrossRef] [Green Version]
- Rice, C.M. Flaviviridae: The Viruses and Their Replication, 3rd ed.; Fields, B.N., Knipe, D.M., Howley, P.M., Chanock, R.M., Melnick, J.L., Monath, T.P., Roizman, B., Straus, S.E., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 1996. [Google Scholar]
- Weiland, F.; Weiland, E.; Unger, G.; Saalm masculineller, A.; Thiel, H.J. Localization of pestiviral envelope proteins E(rns) and E2 at the cell surface and on isolated particles. J. Gen. Virol. 1999, 80, 1157–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Nie, Y.; Wang, P.; Ding, M.; Deng, H. Characterization of classical swine fever virus entry by using pseudotyped viruses: E1 and E2 are sufficient to mediate viral entry. Virology 2004, 330, 332–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Gennip, H.G.; Bouma, A.; van Rijn, P.A.; Widjojoatmodjo, M.N.; Moormann, R.J. Experimental non-transmissible marker vaccines for classical swine fever (CSF) by trans-complementation of E(rns) or E2 of CSFV. Vaccine 2002, 20, 1544–1556. [Google Scholar] [CrossRef]
- Konig, M.; Lengsfeld, T.; Pauly, T.; Stark, R.; Thiel, H.J. Classical swine fever virus: Independent induction of protective immunity by two structural glycoproteins. J. Virol. 1995, 69, 6479–6486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Oirschot, J.T. Vaccinology of classical swine fever: From lab to field. Vet. Microbiol. 2003, 96, 367–384. [Google Scholar] [CrossRef]
- Je, S.H.; Kwon, T.; Yoo, S.J.; Lee, D.U.; Lee, S.; Richt, J.A.; Lyoo, Y.S. Classical Swine Fever Outbreak after Modified Live LOM Strain Vaccination in Naive Pigs, South Korea. Emerg. Infect. Dis. 2018, 24, 798–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mettenleiter, T.C.; Schreurs, C.; Zuckermann, F.; Ben-Porat, T. Role of pseudorabies virus glycoprotein gI in virus release from infected cells. J. Virol. 1987, 61, 2764–2769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mettenleiter, T.C.; Zsak, L.; Kaplan, A.S.; Ben-Porat, T.; Lomniczi, B. Role of a structural glycoprotein of pseudorabies in virus virulence. J. Virol. 1987, 61, 4030–4032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kit, S. Genetically engineered vaccines for control of Aujeszky’s disease (pseudorabies). Vaccine 1990, 8, 420–424. [Google Scholar] [CrossRef]
- Viejo-Borbolla, A.; Munoz, A.; Tabares, E.; Alcami, A. Glycoprotein G from pseudorabies virus binds to chemokines with high affinity and inhibits their function. J. Gen. Virol. 2010, 91, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Firkins, L.D.; Weigel, R.M.; Biehl, L.G.; Hahn, E.C. Field trial to evaluate the immunogenicity of pseudorabies virus vaccines with deletions for glycoproteins G and E. Am. J. Vet. Res. 1997, 58, 976–984. [Google Scholar] [PubMed]
- Platt, K.B.; Mare, C.J.; Hinz, P.N. Differentiation of vaccine strains and field isolates of pseudorabies (Aujeszky’s disease) virus: Thermal sensitivity and rabbit virulence markers. Arch. Virol. 1979, 60, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Fenaux, M.; Halbur, P.G.; Haqshenas, G.; Royer, R.; Thomas, P.; Nawagitgul, P.; Gill, M.; Toth, T.E.; Meng, X.J. Cloned genomic DNA of type 2 porcine circovirus is infectious when injected directly into the liver and lymph nodes of pigs: Characterization of clinical disease, virus distribution, and pathologic lesions. J. Virol. 2002, 76, 541–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, S.I.; Pannhorst, K.; Sangewar, N.; Pavulraj, S.; Wen, X.; Stout, R.W.; Mwangi, W.; Paulsen, D.B. BoHV-1-Vectored BVDV-2 Subunit Vaccine Induces BVDV Cross-Reactive Cellular Immune Responses and Protects against BVDV-2 Challenge. Vaccines 2021, 9, 46. [Google Scholar] [CrossRef]
- Hulst, M.M.; van Gennip, H.G.; Moormann, R.J. Passage of classical swine fever virus in cultured swine kidney cells selects virus variants that bind to heparan sulfate due to a single amino acid change in envelope protein E(rns). J. Virol. 2000, 74, 9553–9561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pomeranz, L.E.; Reynolds, A.E.; Hengartner, C.J. Molecular biology of pseudorabies virus: Impact on neurovirology and veterinary medicine. Microbiol. Mol. Biol. Rev. 2005, 69, 462–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.Y.; Yang, Y.L.; Feng, C.; Sun, M.X.; Peng, J.M.; Tian, Z.J.; Tang, Y.D.; Cai, X.H. Pseudorabies Virus UL24 Abrogates Tumor Necrosis Factor Alpha-Induced NF-kappaB Activation by Degrading P65. Viruses 2020, 12, 51. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, S.I.; Lee, B.J.; Ozkul, A.; Weiss, M.L. Bovine herpesvirus 5 glycoprotein E is important for neuroinvasiveness and neurovirulence in the olfactory pathway of the rabbit. J. Virol. 2000, 74, 2094–2106. [Google Scholar] [CrossRef] [Green Version]
- Pannhorst, K.; Wei, H.; Yezid, H.; He, J.; Chowdhury, S.I. Bovine Herpesvirus 1 UL49.5 Interacts with gM and VP22 To Ensure Virus Cell-to-Cell Spread and Virion Incorporation: Novel Role for VP22 in gM-Independent UL49.5 Virion Incorporation. J. Virol. 2018, 92, e00240-18. [Google Scholar] [CrossRef] [Green Version]
- Pavulraj, S.; Kamel, M.; Stephanowitz, H.; Liu, F.; Plendl, J.; Osterrieder, N.; Azab, W. Equine Herpesvirus Type 1 Modulates Cytokine and Chemokine Profiles of Mononuclear Cells for Efficient Dissemination to Target Organs. Viruses 2020, 12, 999. [Google Scholar] [CrossRef] [PubMed]
- Anonymous. Manual of Diagnostic Tests for Detection of CSF. Available online: https://www.tiho-hannover.de/kliniken-institute/institute/institut-fuer-virologie/eu-and-oie-reference-laboratory/diagnostic-methods (accessed on 22 October 2021).
- Grummer, B.; Fischer, S.; Depner, K.; Riebe, R.; Blome, S.; Greiser-Wilke, I. Replication of classical swine fever virus strains and isolates in different porcine cell lines. Dtsch. Tierarztl. Wochenschr. 2006, 113, 138–142. [Google Scholar] [PubMed]
- Nauwynck, H.J. Functional aspects of Aujeszky’s disease (pseudorabies) viral proteins with relation to invasion, virulence and immunogenicity. Vet. Microbiol. 1997, 55, 3–11. [Google Scholar] [CrossRef]
- Huang, Y.L.; Meyer, D.; Postel, A.; Tsai, K.J.; Liu, H.M.; Yang, C.H.; Huang, Y.C.; Berkley, N.; Deng, M.C.; Wang, F.I.; et al. Identification of a Common Conformational Epitope on the Glycoprotein E2 of Classical Swine Fever Virus and Border Disease Virus. Viruses 2021, 13, 1655. [Google Scholar] [CrossRef]
- Nielsen, J.; Vincent, I.E.; Botner, A.; Ladekaer-Mikkelsen, A.S.; Allan, G.; Summerfield, A.; McCullough, K.C. Association of lymphopenia with porcine circovirus type 2 induced postweaning multisystemic wasting syndrome (PMWS). Vet. Immunol. Immunopathol. 2003, 92, 97–111. [Google Scholar] [CrossRef]
- Darwich, L.; Segales, J.; Mateu, E. Pathogenesis of postweaning multisystemic wasting syndrome caused by Porcine circovirus 2: An immune riddle. Arch. Virol. 2004, 149, 857–874. [Google Scholar] [CrossRef]
- Patterson, A.R.; Ramamoorthy, S.; Madson, D.M.; Meng, X.J.; Halbur, P.G.; Opriessnig, T. Shedding and infection dynamics of porcine circovirus type 2 (PCV2) after experimental infection. Vet. Microbiol. 2011, 149, 91–98. [Google Scholar] [CrossRef]
- Sanchez, R.E., Jr.; Meerts, P.; Nauwynck, H.J.; Ellis, J.A.; Pensaert, M.B. Characteristics of porcine circovirus-2 replication in lymphoid organs of pigs inoculated in late gestation or postnatally and possible relation to clinical and pathological outcome of infection. J. Vet. Diagn. Investig. 2004, 16, 175–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tariq, H.; Batool, S.; Asif, S.; Ali, M.; Abbasi, B.H. Virus-Like Particles: Revolutionary Platforms for Developing Vaccines Against Emerging Infectious Diseases. Front. Microbiol. 2021, 12, 790121. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.W.; Jeng, C.R.; Lin, C.M.; Liu, J.J.; Chang, C.C.; Tsai, Y.C.; Chia, M.Y.; Pang, V.F. The involvement of Fas/FasL interaction in porcine circovirus type 2 and porcine reproductive and respiratory syndrome virus co-inoculation-associated lymphocyte apoptosis in vitro. Vet. Microbiol. 2007, 122, 72–82. [Google Scholar] [CrossRef]
- Seeliger, F.A.; Brugmann, M.L.; Kruger, L.; Greiser-Wilke, I.; Verspohl, J.; Segales, J.; Baumgartner, W. Porcine circovirus type 2-associated cerebellar vasculitis in postweaning multisystemic wasting syndrome (PMWS)-affected pigs. Vet. Pathol. 2007, 44, 621–634. [Google Scholar] [CrossRef] [Green Version]
- Galindo-Cardiel, I.; Grau-Roma, L.; Perez-Maillo, M.; Segales, J. Characterization of necrotizing lymphadenitis associated with porcine circovirus type 2 infection. J. Comp. Pathol. 2011, 144, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Resendes, A.R.; Majo, N.; van den Ingh, T.S.; Mateu, E.; Domingo, M.; Calsamiglia, M.; Segales, J. Apoptosis in postweaning multisystemic wasting syndrome (PMWS) hepatitis in pigs naturally infected with porcine circovirus type 2 (PCV2). Vet. J. 2011, 189, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Sinha, A.; Schalk, S.; Lager, K.M.; Wang, C.; Opriessnig, T. Singular PCV2a or PCV2b infection results in apoptosis of hepatocytes in clinically affected gnotobiotic pigs. Res. Vet. Sci. 2012, 92, 151–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Resendes, A.R.; Majo, N.; Segales, J.; Mateu, E.; Calsamiglia, M.; Domingo, M. Apoptosis in lymphoid organs of pigs naturally infected by porcine circovirus type 2. J. Gen. Virol. 2004, 85, 2837–2844. [Google Scholar] [CrossRef] [PubMed]
- Shibahara, T.; Sato, K.; Ishikawa, Y.; Kadota, K. Porcine circovirus induces B lymphocyte depletion in pigs with wasting disease syndrome. J. Vet. Med. Sci. 2000, 62, 1125–1131. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Liu, C.H.; Roberts, A.I.; Das, J.; Xu, G.; Ren, G.; Zhang, Y.; Zhang, L.; Yuan, Z.R.; Tan, H.S.; et al. Granulocyte-macrophage colony-stimulating factor (GM-CSF) and T-cell responses: What we do and don’t know. Cell Res. 2006, 16, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Delva, J.L.; Nauwynck, H.J.; Mettenleiter, T.C.; Favoreel, H.W. The Attenuated Pseudorabies Virus Vaccine Strain Bartha K61: A Brief Review on the Knowledge Gathered During 60 Years of Research. Pathogens 2020, 9, 897. [Google Scholar] [CrossRef]
- Lin, Y.; Tian, L.; Wang, C.; He, S.; Fang, L.; Wang, Z.; Zhong, Y.; Zhang, K.; Liu, D.; Yang, Q.; et al. Serological Investigation and Genetic Characteristics of Pseudorabies Virus in Hunan Province of China From 2016 to 2020. Front. Vet. Sci. 2021, 8, 1503. [Google Scholar] [CrossRef] [PubMed]
- Fritzemeier, J.; Teuffert, J.; Greiser-Wilke, I.; Staubach, C.; Schluter, H.; Moennig, V. Epidemiology of classical swine fever in Germany in the 1990s. Vet. Microbiol. 2000, 77, 29–41. [Google Scholar] [CrossRef]
- Leifer, I.; Hoffmann, B.; Hoper, D.; Bruun Rasmussen, T.; Blome, S.; Strebelow, G.; Horeth-Bontgen, D.; Staubach, C.; Beer, M. Molecular epidemiology of current classical swine fever virus isolates of wild boar in Germany. J. Gen. Virol. 2010, 91, 2687–2697. [Google Scholar] [CrossRef]
- Ganges, L.; Crooke, H.R.; Bohorquez, J.A.; Postel, A.; Sakoda, Y.; Becher, P.; Ruggli, N. Classical swine fever virus: The past, present and future. Virus Res. 2020, 289, 198151. [Google Scholar] [CrossRef]
- Meyers, G.; Tautz, N.; Becher, P.; Thiel, H.J.; Kummerer, B.M. Recovery of cytopathogenic and noncytopathogenic bovine viral diarrhea viruses from cDNA constructs. J. Virol. 1997, 71, 1735. [Google Scholar] [CrossRef] [Green Version]
- Reimann, I.; Depner, K.; Trapp, S.; Beer, M. An avirulent chimeric Pestivirus with altered cell tropism protects pigs against lethal infection with classical swine fever virus. Virology 2004, 322, 143–157. [Google Scholar] [CrossRef] [Green Version]
- Konig, P.; Blome, S.; Gabriel, C.; Reimann, I.; Beer, M. Innocuousness and safety of classical swine fever marker vaccine candidate CP7_E2alf in non-target and target species. Vaccine 2011, 30, 5–8. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence | |
|---|---|---|
| PRV TK left flanking | Forward (F3) | 5′-gcgaattcccgccttatcaccgcgcccacccgctcc-3′ |
| Reverse (R3) | 5′-cggggtaccttactcacttagcccacgaaggccgccgcgctgatgtcc-3′ | |
| PRV TK right flanking | Forward (F4) | 5′-cgcgttaagctttcgctggacccggacgagcacctgcg -3′ |
| Reverse (R4) | 5′-ccaatgcatggccgagcaggtgcccgcggatgc-3′ | |
| Primer Name | Primer Sequence | |
|---|---|---|
| PRV TK flanking 5′ sequencing | Forward (F5) | 5′-cttgctgggcgtgttgaggttcc-3′ |
| Forward (F6) | 5′-accggcaagagcaccactgc-3′ | |
| Reverse (R5) | 5′-agaaggcgtccttgaccctgg-3′ | |
| Reverse (R6) | 5′-tcctcctcgctcaggctgc-3′ | |
| PRV TK flanking 3′ sequencing | Forward (F7) | 5′-aacacgtcgcgctacctgagc-3′ |
| Forward (F8) | 5′-cgccttcacgtcggagatggg-3′ | |
| Reverse (R7) | 5′-gccttgtacgcgccaaagaggg-3′ | |
| Reverse (R8) | 5′-ggcatggtgacgggcacg-3′ | |
| TK deletion confirmation | Forward (F9) | 5′-gcgcactctgttcgacacggacacggtgg-3′ |
| Reverse (R9) | 5′-ggccaccaccaggttgccgcc-3′ | |
| PCV Cap cloning PRV TK left/PCV2 Cap | Forward (F14) | 5′-acgccgtacctgctgctgcacacg-3′ |
| Reverse (R14) | 5′-gcatcatgtccacggcccaggaaggtgtgg-3′ | |
| Primer/Probe/ds-Gblock Name | Sequence | |
|---|---|---|
| PRV (Major capsid protein) | Forward | 5′-ccatccagtttgaggtgcag-3′ |
| Reverse | 5′-cgaggcgcttgatcatgtag-3′ | |
| Probe | 5′Fam-cccgtcgcgcgcgatcatcg-3′ Tamra | |
| ds-gblock | 5′-ctcagctacgtggccgagggcaccatccagtttgaggtgcagcagccgatgatc gcgcgcgacgggccgcacccggccgaccagcccgtgcacaactacatgatcaagc gcctcgatcgccgctccctcaacgccgc-3′ | |
| Swine Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) | Forward | 5′-atgacaacttcggcatcgtg-3′ |
| Reverse | 5′-ccatccacagtcttctgggt-3′ | |
| Probe | 5′Fam-accacagtccatgccatcactgcc-3′ Tamra | |
| ds-gblock | 5′-gcacccctggccaaggtcatccatgacaacttcggcatcgtggaaggactcatgaccacagtccatgccatcactgccacccagaagactgtggatggcccctctgggaaacgtggcgt-3′ | |
| Porcine circovirus type 2b-capsid protein (ORF2) | Forward | 5′-cacagccctcacctatgacc-3′ |
| Reverse | 5′-aaagtagcgggagtggtagg-3′ | |
| Probe | 5′Fam-cccgccataccataacccagccc-3′ Tamra | |
| ds-gblock | 5′-atgataactttgtaacaaaggccacagccctcacctatgacccctatgtaaactacTcctcccgccataccataacccagcccttctcctaccactcccgctactttacccccaaacctgtcctagatt-3′ | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavulraj, S.; Pannhorst, K.; Stout, R.W.; Paulsen, D.B.; Carossino, M.; Meyer, D.; Becher, P.; Chowdhury, S.I. A Triple Gene-Deleted Pseudorabies Virus-Vectored Subunit PCV2b and CSFV Vaccine Protects Pigs against PCV2b Challenge and Induces Serum Neutralizing Antibody Response against CSFV. Vaccines 2022, 10, 305. https://doi.org/10.3390/vaccines10020305
Pavulraj S, Pannhorst K, Stout RW, Paulsen DB, Carossino M, Meyer D, Becher P, Chowdhury SI. A Triple Gene-Deleted Pseudorabies Virus-Vectored Subunit PCV2b and CSFV Vaccine Protects Pigs against PCV2b Challenge and Induces Serum Neutralizing Antibody Response against CSFV. Vaccines. 2022; 10(2):305. https://doi.org/10.3390/vaccines10020305
Chicago/Turabian StylePavulraj, Selvaraj, Katrin Pannhorst, Rhett W. Stout, Daniel B. Paulsen, Mariano Carossino, Denise Meyer, Paul Becher, and Shafiqul I. Chowdhury. 2022. "A Triple Gene-Deleted Pseudorabies Virus-Vectored Subunit PCV2b and CSFV Vaccine Protects Pigs against PCV2b Challenge and Induces Serum Neutralizing Antibody Response against CSFV" Vaccines 10, no. 2: 305. https://doi.org/10.3390/vaccines10020305
APA StylePavulraj, S., Pannhorst, K., Stout, R. W., Paulsen, D. B., Carossino, M., Meyer, D., Becher, P., & Chowdhury, S. I. (2022). A Triple Gene-Deleted Pseudorabies Virus-Vectored Subunit PCV2b and CSFV Vaccine Protects Pigs against PCV2b Challenge and Induces Serum Neutralizing Antibody Response against CSFV. Vaccines, 10(2), 305. https://doi.org/10.3390/vaccines10020305