
Humoral and Cellular Responses to COVID-19 Vaccination Indicate the Need for Post-Vaccination Testing in Frail Population


 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Population
2.2. Sample Collection
2.3. Antibody Detection
2.4. Pseudovirus Neutralization Assay
2.5. T-Cell Response to SARS-CoV-2 Spike Peptide Pool
2.6. Statistical Analysis
3. Results
3.1. Nursing Home Residents Showed Impaired Antibody Response and Neutralization Following BNT162b2 Vaccination
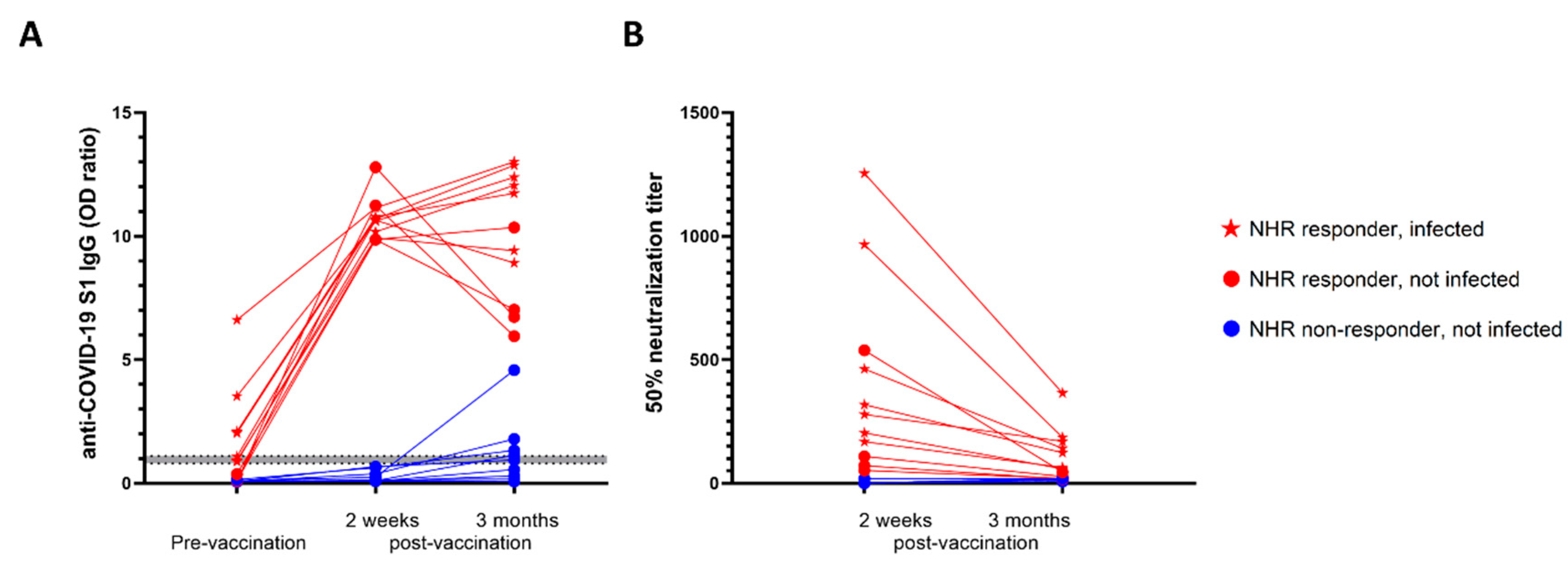
3.2. Pre-Vaccination Infection Status Secured High Antibody Levels 3 Months Post-Vaccination in NHRs
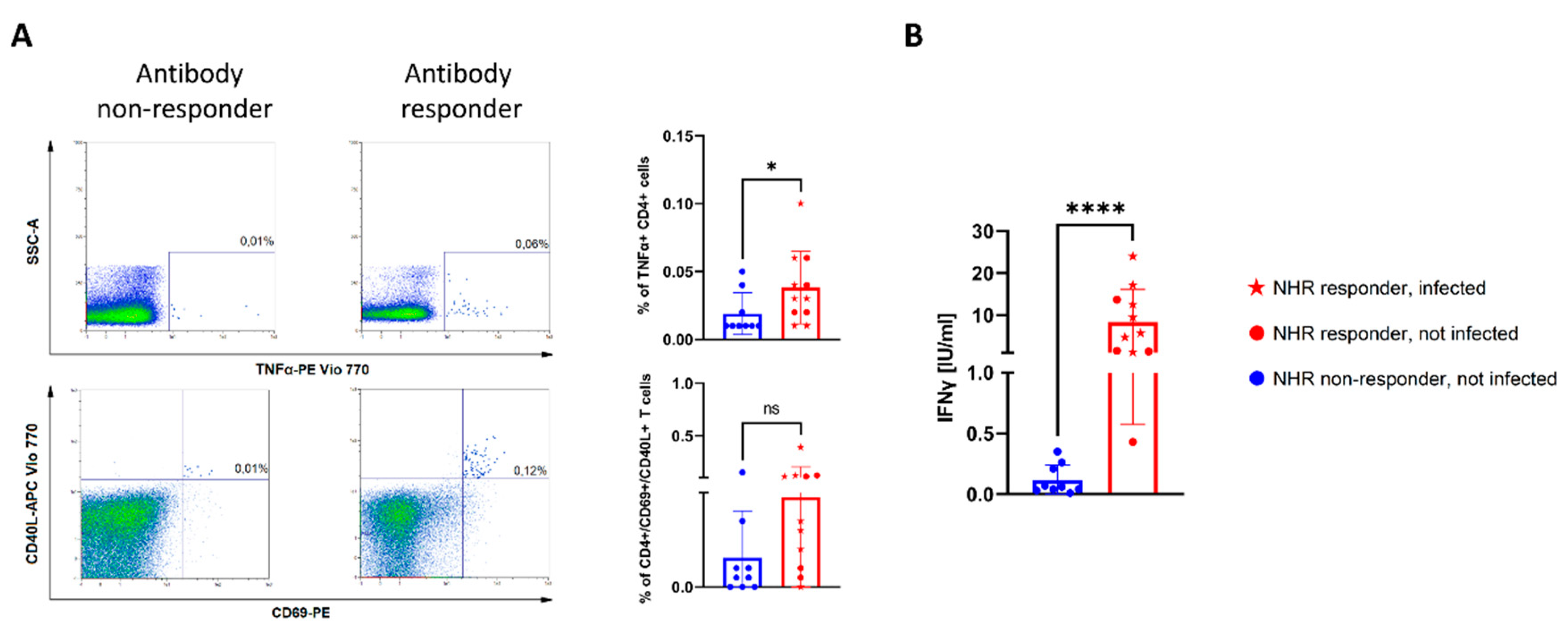
3.3. Lack of COVID-19-Specific T-Cell Responses among NHR Antibody Non-Responders
4. Discussion
5. Conclusions
6. Limitations of the Study
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- O’Driscoll, M.; Dos Santos, G.R.; Wang, L.; Cummings, D.A.T.; Azman, A.S.; Paireau, J.; Fontanet, A.; Cauchemez, S.; Salje, H. Age-specific mortality and immunity patterns of SARS-CoV-2. Nature 2021, 590, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Marc, G.P.; Moreira, E.D.; Zerbini, C.; et al. Safety and efficacy of the BNT162b2 mRNA COVID-19 vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Voysey, M.; Clemens, C.S.A.; Madhi, S.A.; Weckx, L.Y.; Folegatti, P.M.; Aley, P.K.; Angus, B.; Baillie, L.V.; Barnabas, S.L.; Bhorat, Q.E.; et al. Safety and efficacy of the ChAdOx1 nCoV-19 vaccine (AZD1222) against SARS-CoV-2: An interim analysis of four randomised controlled trials in Brazil, South Africa, and the UK. Lancet 2021, 397, 99–111. [Google Scholar] [CrossRef]
- Dyer, O. COVID-19: US reports low rate of new infections in people already vaccinated. BMJ 2021, 373, 1000. [Google Scholar] [CrossRef]
- Dagan, N.; Barda, N.; Kepten, E.; Miron, O.; Perchik, S.; Katz, M.A.; Hernán, M.A.; Lipsitch, M.; Reis, B.; Balicer, R.D. BNT162b2 mRNA Covid-19 Vaccine in a Nationwide Mass Vaccination Setting. N. Engl. J. Med. 2021, 384, 1412–1423. [Google Scholar] [CrossRef]
- Albert, E.; Burgos, J.S.; Salvador, P.; Salas, D.; Vanaclocha, E.G.; Limon, R.; Alcaraz, M.J.; Sanchez-Paya, J.; Dominigo, J.D.; Navarro, D.; et al. Immunological response against SARS-CoV-2 following full-dose administration of Comirnaty(R) COVID-19 vaccine in nursing home residents. Clin. Microbiol. Infect. 2021, 28, 279–284. [Google Scholar] [CrossRef]
- Ríos, S.S.; Romero, M.M.; Zamora, E.B.C.; Sahuquillo, M.T.T.; Rizos, L.R.; Sánchez-Jurado, P.M.; Sánchez-Nievas, G.; Señalada, J.J.B.; Nogueras, I.G.; Cazalla, J.D.D.E.; et al. Immunogenicity of the BNT162b2 vaccine in frail or disabled nursing home residents: COVID-A study. J. Am. Geriatr. Soc. 2021, 69, 1441–1447. [Google Scholar] [CrossRef]
- Urlaub, D.; Wolfsdroff, N.; Durak, D.; Renken, F.; Watzl, C. SARS-CoV-2 infection shortly after BNT162b2 vaccination results in high anti-spike antibody levels in nursing home residents and staff. Immun. Inflamm. Dis. 2021, 9, 1702–1706. [Google Scholar] [CrossRef]
- Canaday, D.H.; Carais, L.; Oyebanji, O.A.; Keresztesy, D.; Wilk, D.; Payne, M.; Aung, H.; Kerri, D.; Lam, E.C.; Rowley, C.F.; et al. Reduced BNT162b2 messenger RNA vaccine response in severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2)-naive nursing home residents. Clin. Infect. Dis. 2021, 73, 2112–2115. [Google Scholar] [CrossRef]
- Bergwerk, M.; Gonen, T.; Lustig, Y.; Amit, S.; Lipsitch, M.; Cohen, C.; Mandelboim, M.; Levin, E.G.; Rubin, C.; Indenbaum, V.; et al. COVID-19 breakthrough infections in vaccinated health care workers. N. Engl. J. Med. 2021, 385, 1474–1484. [Google Scholar] [CrossRef] [PubMed]
- Bernal, J.L.; Andrews, N.; Gower, C.; Gallagher, E.; Simmons, R.; Thelwall, S.; Stowe, J.; Tessier, E.; Groves, N.; Dabrera, G.; et al. Effectiveness of Covid-19 Vaccines against the B.1.617.2 (Delta) Variant. N. Engl. J. Med. 2021, 385, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, A.; Wiedra, M.; Grikscheit, K.; Toptan, T.; Schen, B.; Pallas, C.; Metzler, M.; Khomer, N.; Hoehl, S.; Helfritz, F.A.; et al. Reduced neutralization of SARS-CoV-2 omicron variant by vaccine sera and monoclonal antibodies. medRxiv 2021, 21267432. [Google Scholar]
- Stephenson, J. COVID-19 vaccinations in nursing home residents and staff give robust protection, though breakthrough infections still possible. JAMA Health Forum 2021, 2, e211195. [Google Scholar] [CrossRef]
- Manisty, C.; Otter, A.D.; Treibel, T.A.; McKnight, A.; Altmann, D.M.; Brooks, T. Antibody response to first BNT162b2 dose in previously SARS-CoV-2-infected individuals. Lancet 2021, 397, 1057–1058. [Google Scholar] [CrossRef]
- Ebinger, J.E.; Fert-Bober, J.; Printsev, I.; Wu, M.; Sun, N.; Prostko, J.C.; Frias, E.C.; Stewart, J.L.; Eyk, J.E.V.; Braun, J.G.; et al. Antibody responses to the BNT162b2 mRNA vaccine in individuals previously infected with SARS-CoV-2. Nat. Med. 2021, 27, 981–984. [Google Scholar] [CrossRef]
- Blain, H.; Tuaillon, E.; Gamon, L.; Pisoni, A.; Miot, S.; Rolland, Y.; Picot, M.; Bousquet, J. Antibody response after one and two jabs of the BNT162b2 vaccine in nursing home residents: The CONsort-19 study. Allergy 2021, 77, 271–281. [Google Scholar] [CrossRef]
- Monge, S.; Olmedo, C.; Alejos, B.; Lapen, M.; Sierr, M.J.; Limia, A. Direct and indirect effectiveness of mRNA vaccination against SARS-CoV-2 infection in long-term care facilities in Spain. medRxiv 2021, 27, 21255055. [Google Scholar]
- Criscuolo, E.; Diotti, R.A.; Strollo, M.; Rolla, S.; Ambrosi, A.; Locatelli, M.; Burioni, R.; Mancini, N.; Clementi, M.; Clementi, N.; et al. Weak correlation between antibody titers and neutralizing activity in sera from SARS-CoV-2 infected subjects. J. Med. Virol. 2021, 93, 2160–2167. [Google Scholar] [CrossRef]
- Collier, D.A.; Fierra, I.A.T.; Kotagiri, P.; Datir, R.P.; Lim, E.Y.; Touzier, E.; Abudulahi, A.; Elmer, A.; Kingston, N.; Graves, B.; et al. Age-related immune response heterogeneity to SARS-CoV-2 vaccine BNT162b2. Nature 2021, 596, 417–422. [Google Scholar] [CrossRef]
- Muller, L.; Andree, M.; Moskorz, W.; Drelxler, I.; Walotka, L.; Grothman, R.; Ptok, J.; Hillebrandt, J.; Ritchie, A.; Babl, D.; et al. Age-dependent immune response to the Biontech/Pfizer BNT162b2 COVID-19 vaccination. Clin. Infect. Dis. 2021, 73, 2065–2072. [Google Scholar] [PubMed]
- Gilbert, P.B.; Montefiori, D.C.; McDermott, A.B.; Fong, Y.; Benkeser, D.; Deng, W.; Zhou, H.; Houchens, C.R.; Martins, K.; Jayashankar, L.; et al. Immune correlates analysis of the mRNA-1273 COVID-19 vaccine efficacy clinical trial. Science 2022, 375, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Aw, D.; Woodrow, L.; Ogliari, G.; Harwood, R. Association of frailty with mortality in older inpatients with Covid-19: A cohort study. Age Ageing 2020, 49, 915–922. [Google Scholar] [CrossRef] [PubMed]
- Bar-On, Y.M.; Goldberg, Y.; Mandel, M.; Bodenheimer, O.; Freedman, L.; Alroy-Preis, S.; Ash, N.; Huppert, A.; Milo, R. Protection against Covid-19 by BNT162b2 Booster across Age Groups. N. Engl. J. Med. 2021, 385, 2421–2430. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Witkowski, W.; Gerlo, S.; De Smet, E.; Wejda, M.; Acar, D.; Callens, S.; Heytens, S.; Padalko, E.; Vercruysse, H.; Cools, P.; et al. Humoral and Cellular Responses to COVID-19 Vaccination Indicate the Need for Post-Vaccination Testing in Frail Population. Vaccines 2022, 10, 260. https://doi.org/10.3390/vaccines10020260
Witkowski W, Gerlo S, De Smet E, Wejda M, Acar D, Callens S, Heytens S, Padalko E, Vercruysse H, Cools P, et al. Humoral and Cellular Responses to COVID-19 Vaccination Indicate the Need for Post-Vaccination Testing in Frail Population. Vaccines. 2022; 10(2):260. https://doi.org/10.3390/vaccines10020260
Chicago/Turabian StyleWitkowski, Wojciech, Sarah Gerlo, Evelien De Smet, Magdalena Wejda, Delphine Acar, Steven Callens, Stefan Heytens, Elizaveta Padalko, Hanne Vercruysse, Piet Cools, and et al. 2022. "Humoral and Cellular Responses to COVID-19 Vaccination Indicate the Need for Post-Vaccination Testing in Frail Population" Vaccines 10, no. 2: 260. https://doi.org/10.3390/vaccines10020260
APA StyleWitkowski, W., Gerlo, S., De Smet, E., Wejda, M., Acar, D., Callens, S., Heytens, S., Padalko, E., Vercruysse, H., Cools, P., & Vandekerckhove, L. (2022). Humoral and Cellular Responses to COVID-19 Vaccination Indicate the Need for Post-Vaccination Testing in Frail Population. Vaccines, 10(2), 260. https://doi.org/10.3390/vaccines10020260