p-Hydroxybenzoic Acid β-d-Glucosyl Ester and Cimidahurinine with Antimelanogenesis and Antioxidant Effects from Pyracantha angustifolia via Bioactivity-Guided Fractionation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
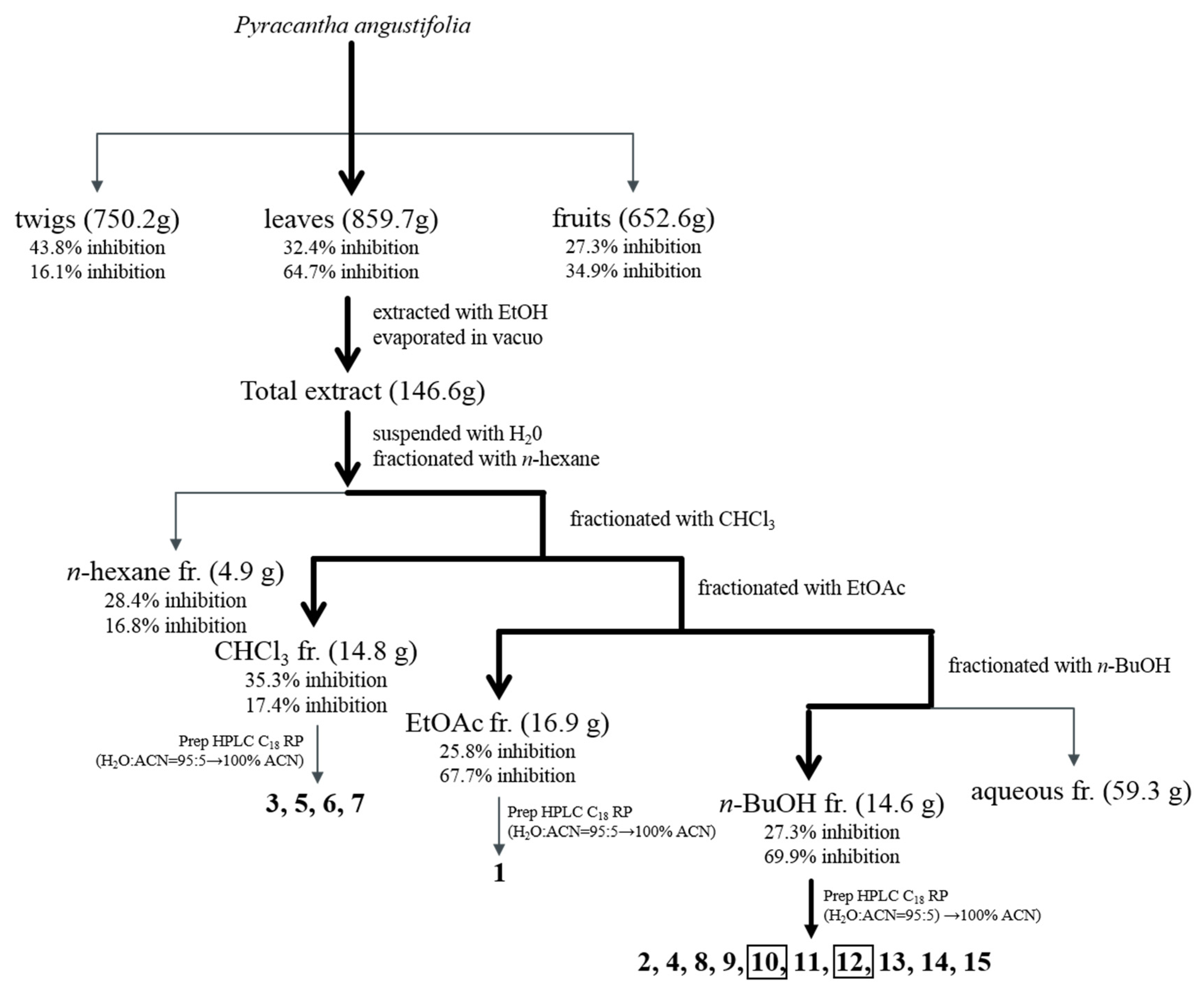
2.2. Extraction and Isolation
2.3. Anti-Melanogenesis Assay
2.3.1. Cell Culture and Viability Assay
2.3.2. Melanin Content Assay
2.3.3. Cellular TYR Activity Assay
2.3.4. Western Blot Analysis
2.3.5. ROS Assay
2.4. Antioxidant Assay
2.4.1. Preparation of Sample
2.4.2. DPPH Assay
2.4.3. ABTS Assay
2.5. Statistical Analysis
3. Results
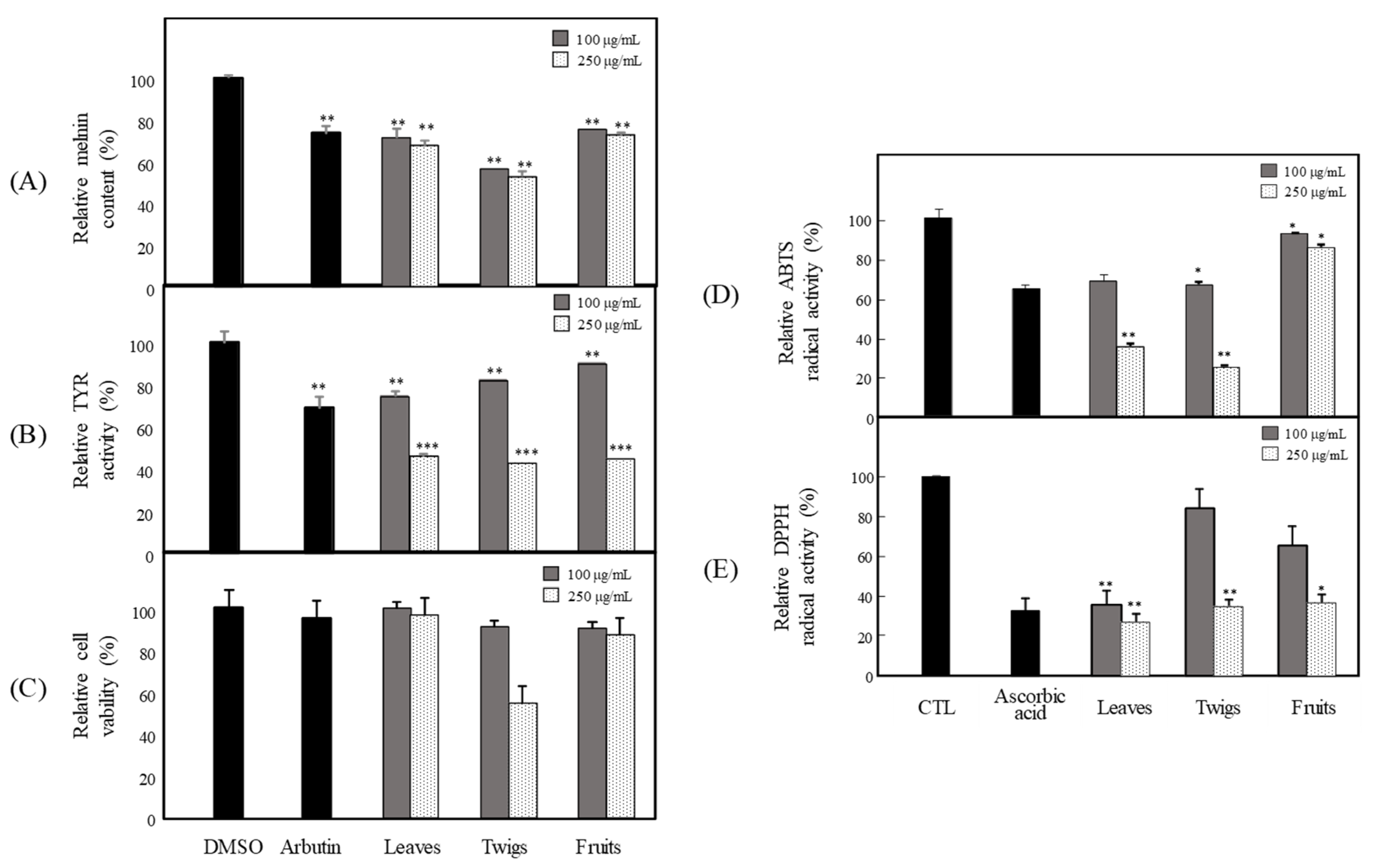
3.1. Anti-Melanogenesis and Antioxidant Effects of Extract
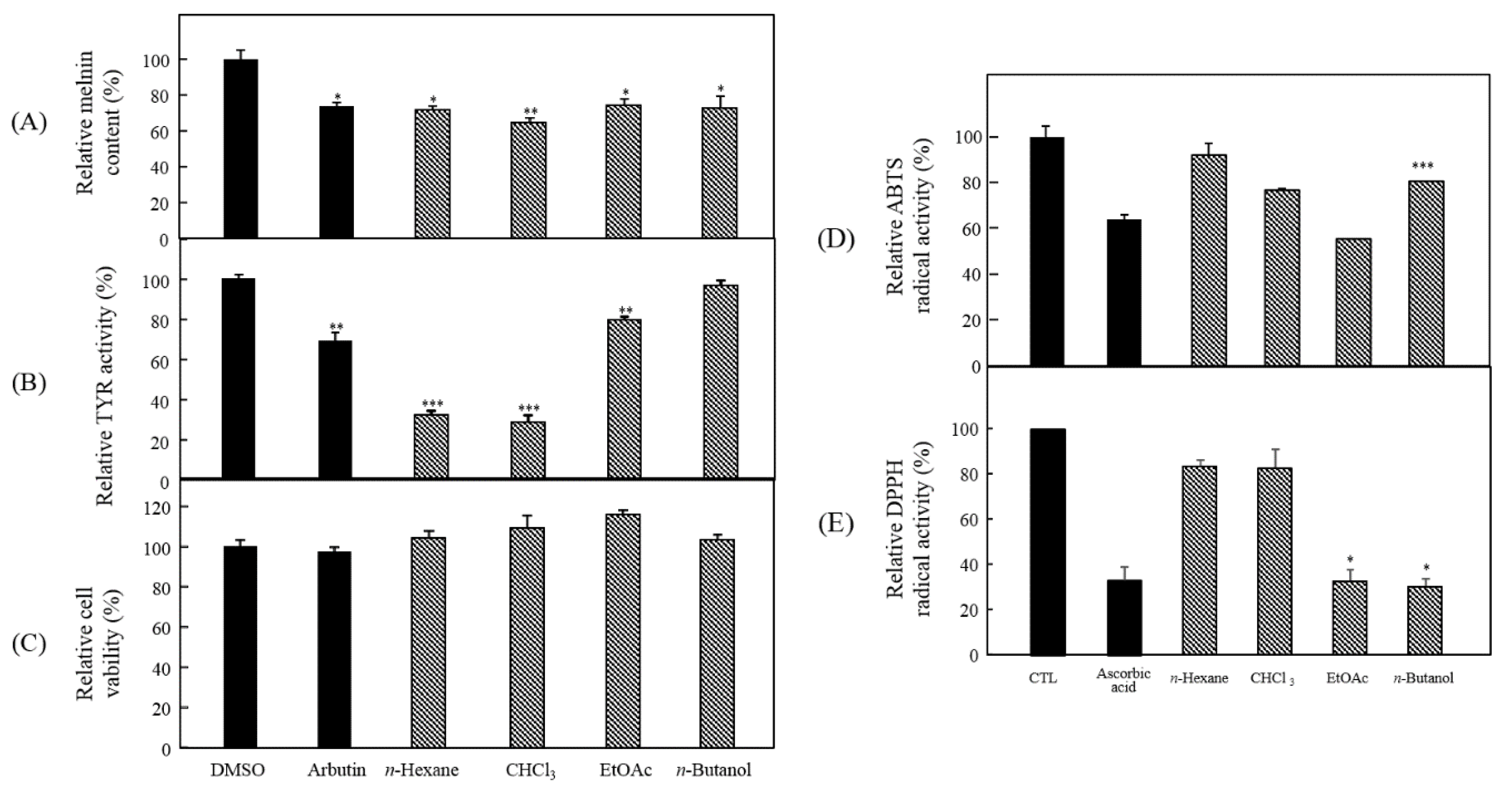
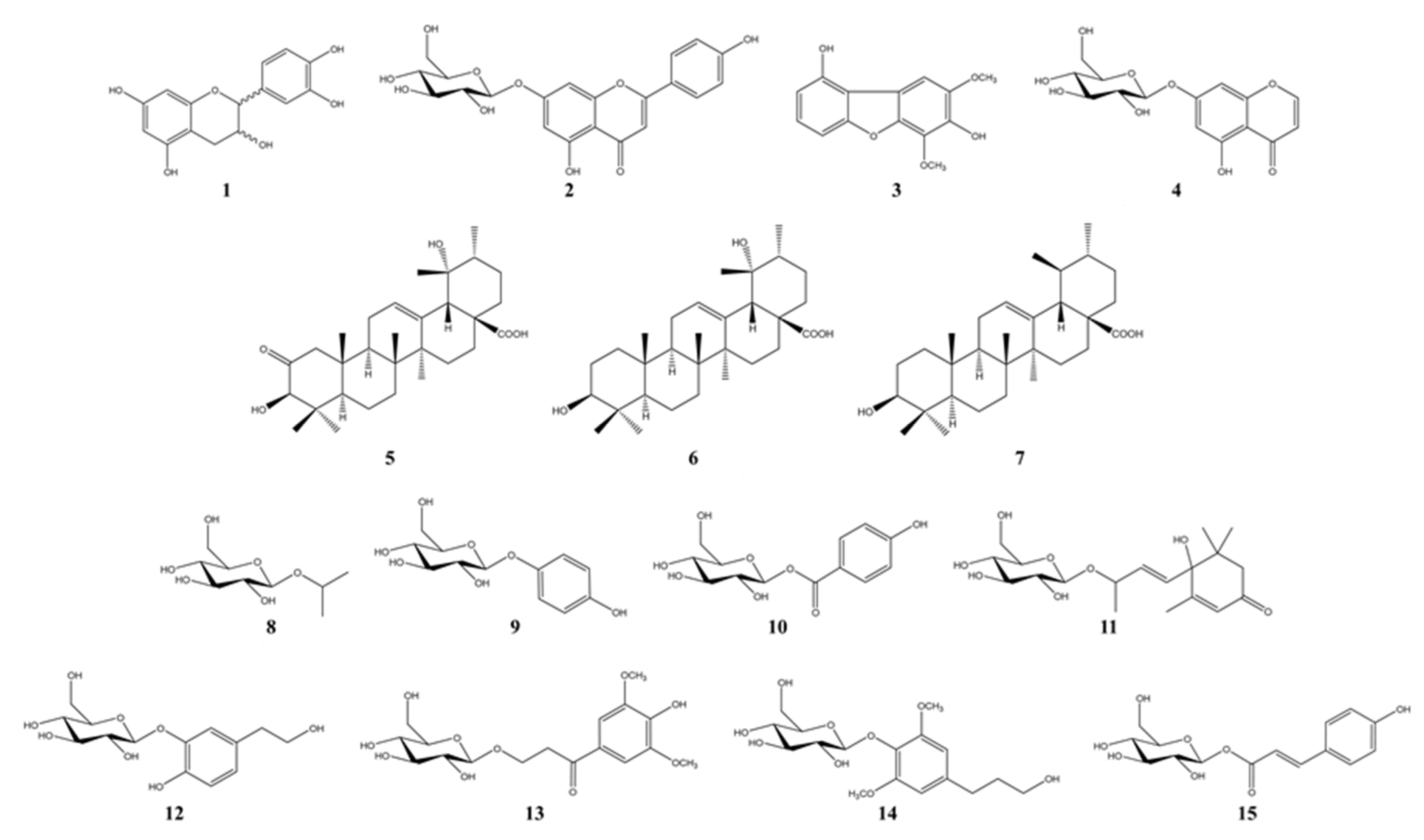
3.2. Bioactivity-Guided Isolation of Active Phytochemicals from P. angustifolia
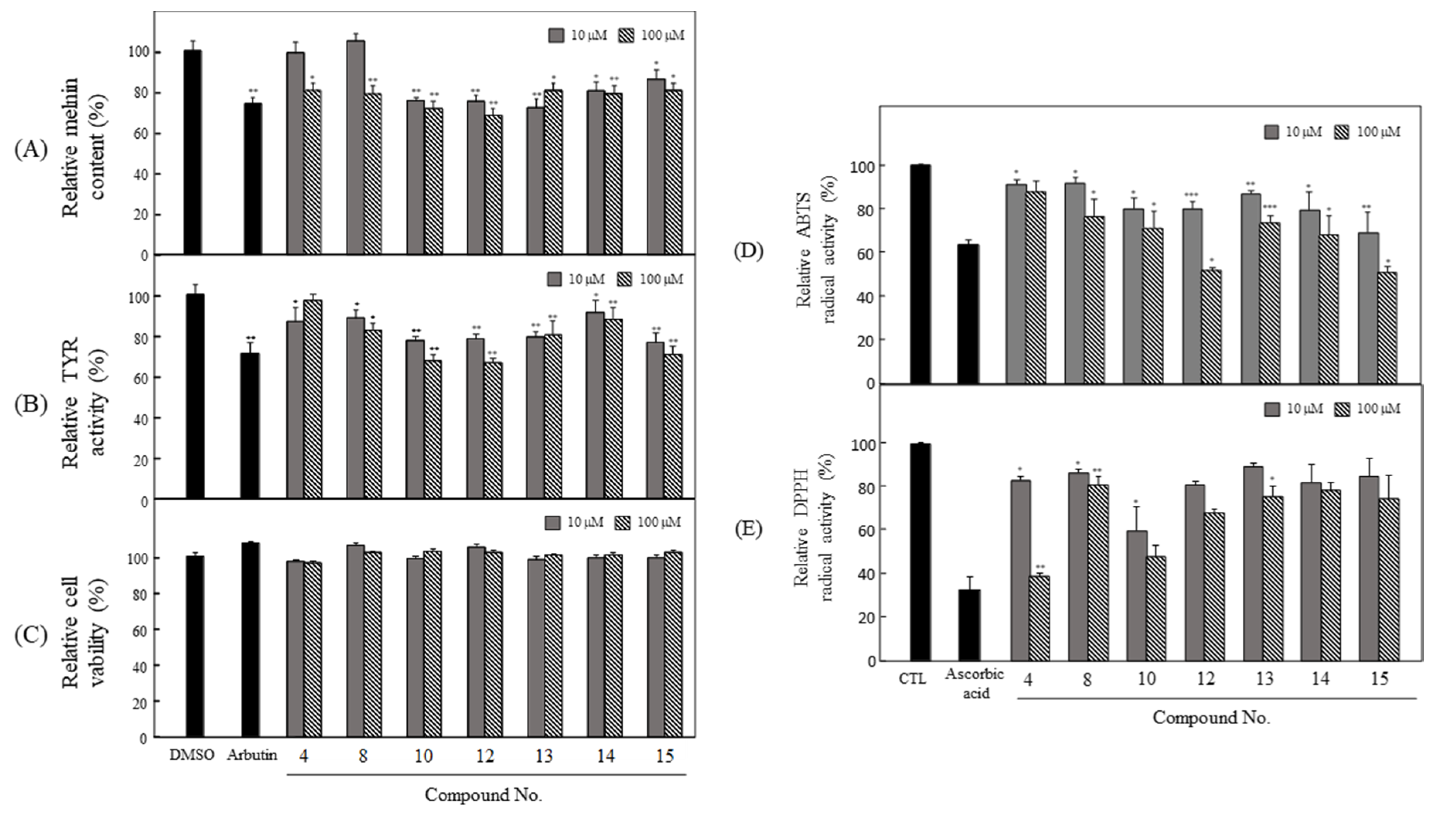
3.3. Anti-Melanogenesis and Antioxidant Effects of Seven Compounds Isolated from P. angustifolia Fraction
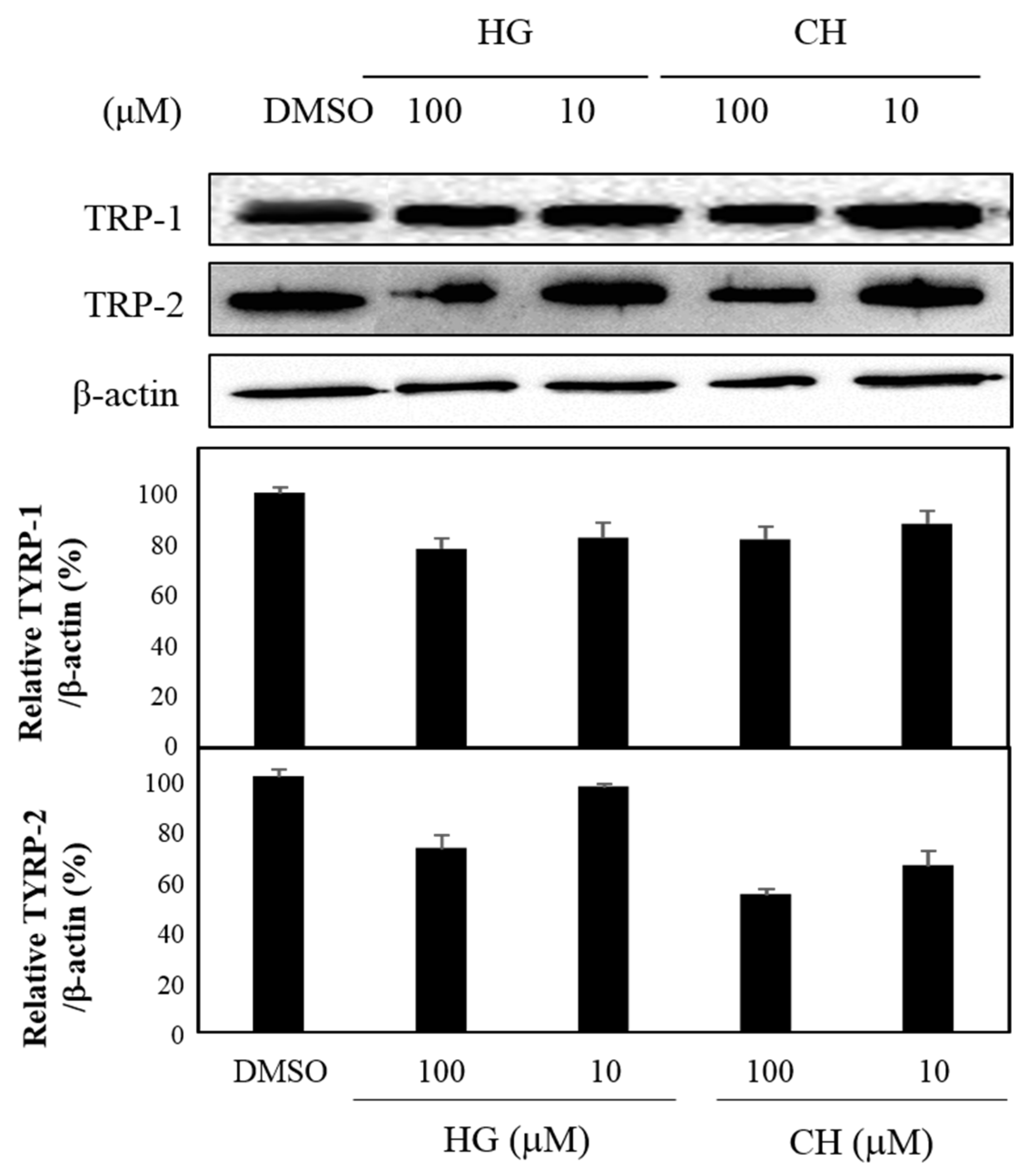
3.4. Effects of HG and CH on TYRP-1 and TYRP-2 Expression
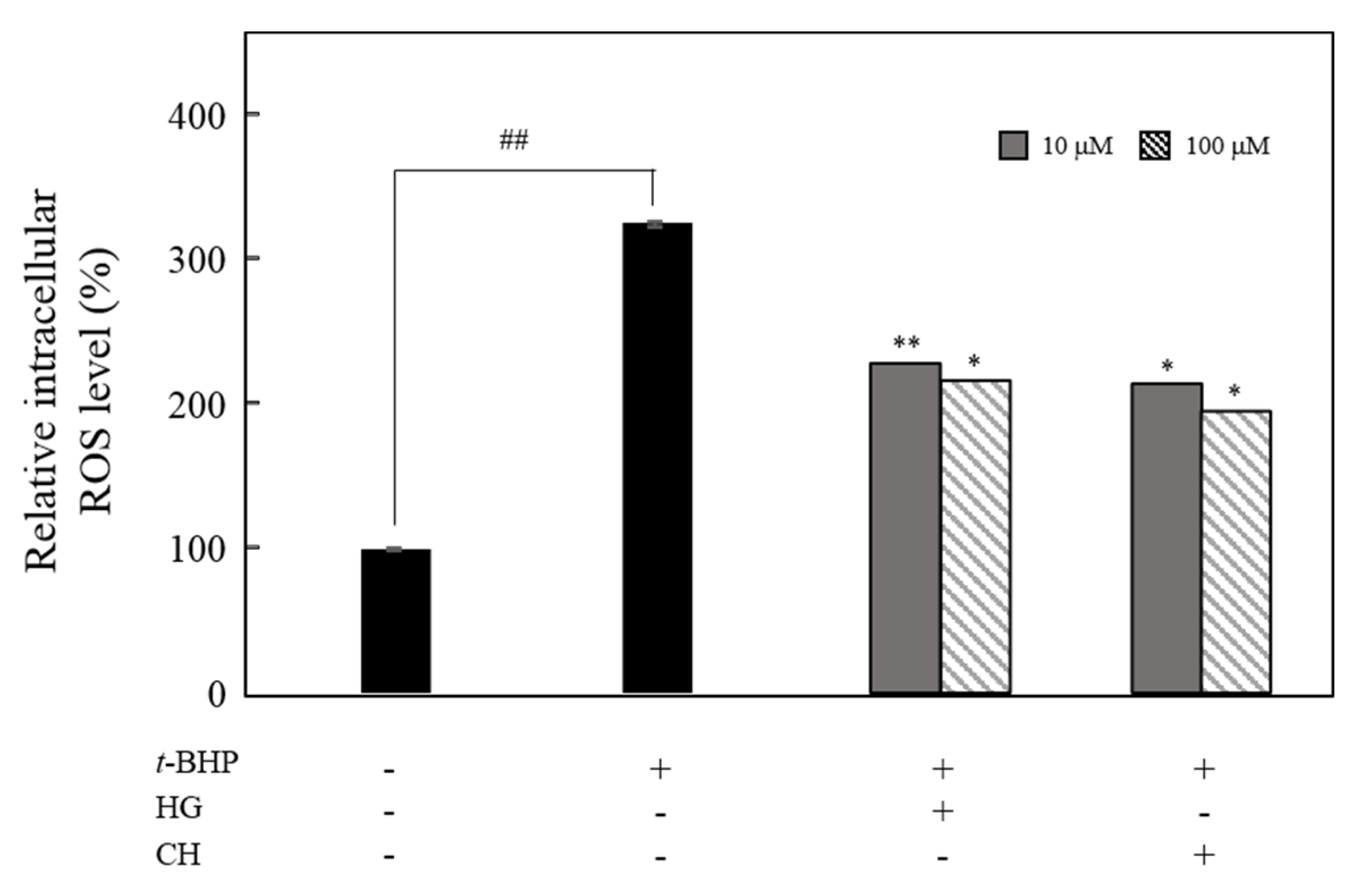
3.5. Effect of HG and CH on the Intracellular ROS Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- D’Mello, S.A.; Finlay, G.J.; Baguley, B.C.; Askarian-Amiri, M.E. Signaling pathways in melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar] [CrossRef] [PubMed]
- Bonaventure, J.; Domingues, M.J.; Larue, L. Cellular and molecular mechanisms controlling the migration of melanocytes and melanoma cells. Pigment Cell Melanoma Res. 2013, 26, 316–325. [Google Scholar] [CrossRef] [PubMed]
- Cichorek, M.; Wachulska, M.; Stasiewicz, A.; Tyminska, A. Skin melanocytes: Biology and development. Adv. Dermatol. Allergol. 2013, 30, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Denat, L.; Kadekaro, L.M.; Marrot, L.; Leachman, A.S.; Adbel-Malek, A.Z. Melanocytes as instigators and victims of oxidative stress. J. Investig. Dermatol. 2014, 134, 1512–1518. [Google Scholar] [CrossRef] [PubMed]
- Miyamura, Y.; Coelho, S.G.; Wolber, R.; Miller, S.A.; Wakamatsu, K.; Zmudzka, B.Z.; Ito, S.; Smuda, C.; Passeron, T.; Choi, W.; et al. Regulation of human skin pigmentation and responses to ultraviolet radiation. Pigment Cell Melanoma Res. 2007, 20, 2–13. [Google Scholar] [CrossRef]
- Brenner, M.; Hearing, V.J. The protective role of melanin against UV damage in human skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar] [CrossRef]
- De Gruijl, F.R.; van Kranen, H.J.; Mullenders, L.H. UV-induced DNA damage, repair, mutations and oncogenic pathways in skin cancer. J. Photochem. Photobiol. 2001, B63, 19–27. [Google Scholar] [CrossRef]
- Korner, A.; Pawelek, J. Mammalian tyrosinase catalases three reactions in the biocynthesis of melanin. Science 1982, 217, 1163–1165. [Google Scholar] [CrossRef]
- Tsukamoto, K.; Jackson, I.J.; Urabe, K.; Montague, P.M.; Hearing, V.J. A second tyrosinase-related protein, TRP-2, is a melanogenic enzyme termed DOPAchrome tautomerase. EMBO J. 1992, 11, 519–526. [Google Scholar] [CrossRef]
- Videira, I.F.; Moura, D.F.; Magina, S. Mechanisms regulating melanogenesis. An. Bras. Dermatol. 2013, 88, 76–83. [Google Scholar] [CrossRef]
- Brenner, M.; Berking, C. Principles of skin pigmentation. Biochemistry and regulation of melanogenesis. Hautarzt 2010, 61, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Brenner, M.; Hearing, V.J. The regulation of skin pigmentation. J. Biol. Chem. 2007, 282, 27557–27561. [Google Scholar] [CrossRef] [PubMed]
- Epstein, J.H. Photocarcinogenesis, skin cancer, and aging. J. Am. Acad. Dermatol. 1983, 9, 487–502. [Google Scholar] [CrossRef]
- Bjelakovic, G.; Nikolova, D.; Gluud, C. Antioxidant supplements and mortality. Curr. Opin. Clin. Nutr. Metab. Care 2014, 17, 40–44. [Google Scholar] [CrossRef]
- Yang, S.H.; Tsasakis, A.M.; Tzanakakis, G.; Kim, H.S.; Le, B.; Sifaki, M.; Spandidos, D.A.; Tsukamoto, C.; Chung, G. Soyasaponin Ag inhibits α-MSH-induced melanogenesis in B16F10 melanoma cells via the downregulation of TRP-2. Int. J. Mol. Med. 2017, 40, 631–636. [Google Scholar] [CrossRef]
- Kwon, M.; Lim, S.J.; Lee, B.; Shin, T.; Kim, H.R. Ethanolic extract of sargassum serratifolium inhibits adipogenesis in 3T3-L1 preadipocytes by cell cycle arrest. J. Appl. Phycol. 2018, 30, 559–568. [Google Scholar] [CrossRef]
- Lin, Y.S.; Chuang, M.T.; Chen, C.H.; Chien, M.Y.; Hou, W.C. Nicotinic acid hydroxamate downregulated the melanin synthesis and tyrosinase activity through activating the MEK/ERK and AKT/GSK3beta signaling pathways. J. Agric. Food Chem. 2012, 60, 4859–4864. [Google Scholar] [CrossRef]
- Blois, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Proestos, C.; Lytoudi, K.; Mavromelanidou, O.; Zoumpoulakis, P.; Sinanoglou, V. Antioxidant Capacity of Selected Plant Extracts and their Essential Oils. Antioxidants 2013, 2, 11–22. [Google Scholar] [CrossRef]
- Sarangarajan, R.; Boissy, R.E. Tyrp1 and oculocutaneous albinism type 3. Pigment Cell Res. 2001, 14, 437–444. [Google Scholar] [CrossRef]
- Zhu Yue, Z.; Yulong, W.; Zhang, J.; Zhang, N.; Lei, Y.; Gan, Z.; Sun, A. Isolation and identification of procyanidins in aronia melanocarpa using NMR, LC-IT-TOF/MS/MS and MALDI-TOF MS. J. Food Sci. Technol. 2019, 4, 614–628. [Google Scholar]
- Peng, Z.F.; Strack, D.; Baumert, A.; Subramaniam, R.; Goh, N.K.; Chia, T.F.; Tan, S.N.; Chia, L.S. Antioxidant Flavonoids from Leaves of Polygonum Hydropiper L. Phytochemistry 2003, 62, 219–228. [Google Scholar] [CrossRef]
- Lin, R.; Chen, M.; Liu, Y.; Lin, Y.; Lu, M.; Hsu, F.; Lee, M. New Whitening Constituents from Taiwan-Native Pyracantha Koidzumii: Structures and Tyrosinase Inhibitory Analysis in Human Epidermal Melanocytes. Int. J. Mol. Sci. 2015, 16, 28598–28613. [Google Scholar] [CrossRef]
- Backhouse, N.; Delporte, C.; Negrete, R.; Feliciano, S.S.; López-Pérez, J. Bioactive Phenolic Derivatives from Acaena Splendens Methanol Extract. Phytother. Res. 2002, 16, 562–566. [Google Scholar] [CrossRef]
- Xu, H.; Zeng, F.; Wan, M.; Sim, K. Anti-HIV Triterpene Acids from Geum Japonicum. Nat. Prod. 1996, 59, 643–645. [Google Scholar] [CrossRef]
- Cheng, J.; Zhang, L.; Cheng, H.; Chiou, C.; Lee, I.; Kuo, Y. Cytotoxic Hexacyclic Triterpene Acids from Euscaphis Japonica. J. Nat. Prod. 2010, 73, 1655–1658. [Google Scholar] [CrossRef]
- Lee, T.; Juang, S.; Hsu, F.; Wu, C. Triterpene Acids from the Leaves of Planchonella Duclitan (Blanco) Bakhuizan. J. Chin. Chem. Soc. 2005, 52, 1275–1280. [Google Scholar] [CrossRef]
- Du, X.; Yoshizawa, T.; Shoyama, Y. Butanoic Acid Glucoside Composition of Whole Body and in Vitro Plantlets of Anoectochilus Formosanus. Phytochemistry 1998, 49, 1925–1928. [Google Scholar] [CrossRef]
- Sugimoto, K.; Nishimura, T.; Nomura, K.; Sugimoto, K.; Kuriki, T. Syntheses of Arbutin-α-Glycosides and a Comparison of their Inhibitory Effects with those of α-Arbutin and Arbutin on Human Tyrosinase. Chem. Pharm. Bull. 2003, 51, 798–801. [Google Scholar] [CrossRef]
- Shen, Y.; Terazawa, M. Dihydroroseoside, a New Cyclohexanone Glucoside, from the Leaves of Shirakamba (Betula Platyphylla Sukatchev Var. Japonica Hara). J. Wood Sci. 2001, 47, 145–148. [Google Scholar] [CrossRef]
- Zhou, X.; Yan, L.; Yin, P.; Shi, L.; Zhang, J.; Liu, Y.; Ma, C. Structural Characterisation and Antioxidant Activity Evaluation of Phenolic Compounds from Cold-Pressed Perilla Frutescens Var. Arguta Seed Flour. Food Chem. 2014, 164, 150–157. [Google Scholar] [CrossRef]
- Jiang, Z.; Li, S.; Li, W.; Guo, J.; Tian, K.; Hu, Q.; Huang, X. Phenolic Glycosides from Ficus Tikoua and their Cytotoxic Activities. Carbohydr. Res. 2013, 382, 19–24. [Google Scholar] [CrossRef]
- Pan, H.; Chen, B.; Fu, L.; Wang, M. Chemical constituents from the stems of Dendrobium denneanum (II). Chin. J. Appl. Environ. Biol. 2013, 19, 952–955. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, J.; Liu, J.-H.; Yu, B.-Y. Biotransformation of p-, m-, and o-hydroxybenzoic acids by Panax ginseng hairy root cultures. J. Mol. Catal. B Enzymatic 2008, 54, 72–75. [Google Scholar] [CrossRef]
- He, X.; Liu, R.H. Phytochemicals of Apple Peels: Isolation, Structure Elucidation, and their Antiproliferative and Antioxidant Activities. J. Agric. Food Chem. 2008, 56, 9905–9910. [Google Scholar] [CrossRef]
- Kazuomi, S.; Masaru, T. Depigmenting Effect of Catechins. Molecules 2009, 14, 4425–4432. [Google Scholar]
- Seong, Z.-K.; Lee, Z.-K.; Poudel, A.; Oh, S.-R.; Lee, H.-K. Constituents of Cryptotaenia japonica Inhibit Melanogenesis via CREB- and MAPK-Associated Signaling Pathways in Murine B16 Melanoma Cells. Molecules 2016, 21, 1296. [Google Scholar] [CrossRef]
- Song, N.Y.; Cho, J.G.; Im, D.M.; Lee, D.Y.; Wu, Q.; Seo, W.D.; Kang, H.C.; Lee, Y.H.; Bae, N.I. Triterpenoids from Fragaria ananassa calyx and their inhibitory effects on melanogenesis in B16-F10 mouse melanoma cells. Nat. Prod. Res. 2013, 27, 2219–2223. [Google Scholar] [CrossRef]
- Jeon, H.J.; Kim, K.; Kim, Y.D.; Lee, S.E. Antimelanogenic activities of piperlongumine derived from Piper longum on murine B16F10 melanoma cells in vitro and zebrafish embryos in vivo: Its molecular mode of depigmenting action. Appl. Biol. Chem. 2019, 62. [Google Scholar] [CrossRef]
- Lim, Y.J.; Lee, E.H.; Kang, T.H.; Ha, S.K.; Oh, M.S.; Kim, S.M.; Yoon, T.J.; Gang, C.; Park, J.H.; Kim, S.Y. Inhibitory effects of arbutin on melanin biosynthesis of α-melanocyte stimulating hormone-induced hyperpigmentation in cultured brownish guinea pig skin tissues. Arch. Pharm. Res. 2009, 32, 367–373. [Google Scholar] [CrossRef]
- Han, E.; Chang, B.; Kim, D.; Cho, H.; Kim, S. Melanogenesis inhibitory effect of aerial part of Pueraria thunbergiana in vitro and in vivo. Arch. Dermatol. Res. 2015, 307, 57–72. [Google Scholar] [CrossRef]
- Manandhar, B.; Wagle, A.; Seong, S.H.; Paudel, P.; Kim, H.R.; Jung, H.A.; Choi, J.S. Phlorotannins with potential anti-tyrosinase and antioxidant activity isolated from the marine seaweed Ecklonia stolonifera. Antioxidants 2019, 8, 240. [Google Scholar] [CrossRef]
- Fang, D.; Tsuji, Y.; Setaluri, V. Selective down-regulation of tyrosinase family gene TYRP1 by inhibition of the activity of melanocyte transcription factor, MITF. Nucleic Acids Res. 2002, 30, 3096–3106. [Google Scholar] [CrossRef]
- Yasumoto, K.; Yokoyama, K.; Shibata, K.; Tomita, Y.; Shibahara, S. Microphthalmia-associated transcription factor as a regulator for melanocyte-specific transcription of the human tyrosinase gene. Mol. Cell. Biol. 1994, 14, 8058–8070. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shim, S.-Y.; Lee, Y.E.; Song, H.Y.; Lee, M. p-Hydroxybenzoic Acid β-d-Glucosyl Ester and Cimidahurinine with Antimelanogenesis and Antioxidant Effects from Pyracantha angustifolia via Bioactivity-Guided Fractionation. Antioxidants 2020, 9, 258. https://doi.org/10.3390/antiox9030258
Shim S-Y, Lee YE, Song HY, Lee M. p-Hydroxybenzoic Acid β-d-Glucosyl Ester and Cimidahurinine with Antimelanogenesis and Antioxidant Effects from Pyracantha angustifolia via Bioactivity-Guided Fractionation. Antioxidants. 2020; 9(3):258. https://doi.org/10.3390/antiox9030258
Chicago/Turabian StyleShim, Sun-Yup, Ye Eun Lee, Hwa Young Song, and Mina Lee. 2020. "p-Hydroxybenzoic Acid β-d-Glucosyl Ester and Cimidahurinine with Antimelanogenesis and Antioxidant Effects from Pyracantha angustifolia via Bioactivity-Guided Fractionation" Antioxidants 9, no. 3: 258. https://doi.org/10.3390/antiox9030258
APA StyleShim, S.-Y., Lee, Y. E., Song, H. Y., & Lee, M. (2020). p-Hydroxybenzoic Acid β-d-Glucosyl Ester and Cimidahurinine with Antimelanogenesis and Antioxidant Effects from Pyracantha angustifolia via Bioactivity-Guided Fractionation. Antioxidants, 9(3), 258. https://doi.org/10.3390/antiox9030258