Aerobic Training Down-Regulates Pentraxin 3 and Pentraxin 3/Toll-Like Receptor 4 Ratio, Irrespective of Oxidative Stress Response, in Elderly Subjects

 ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Experimental Protocol
2.3. Blood Sampling
2.4. Plasma Assays
2.5. Isolation of PBMCs
2.6. Western Blot Analysis
2.7. Statistical Analysis
3. Results
3.1. Measurements of Inflammatory and Oxidative Biomarkers at Baseline
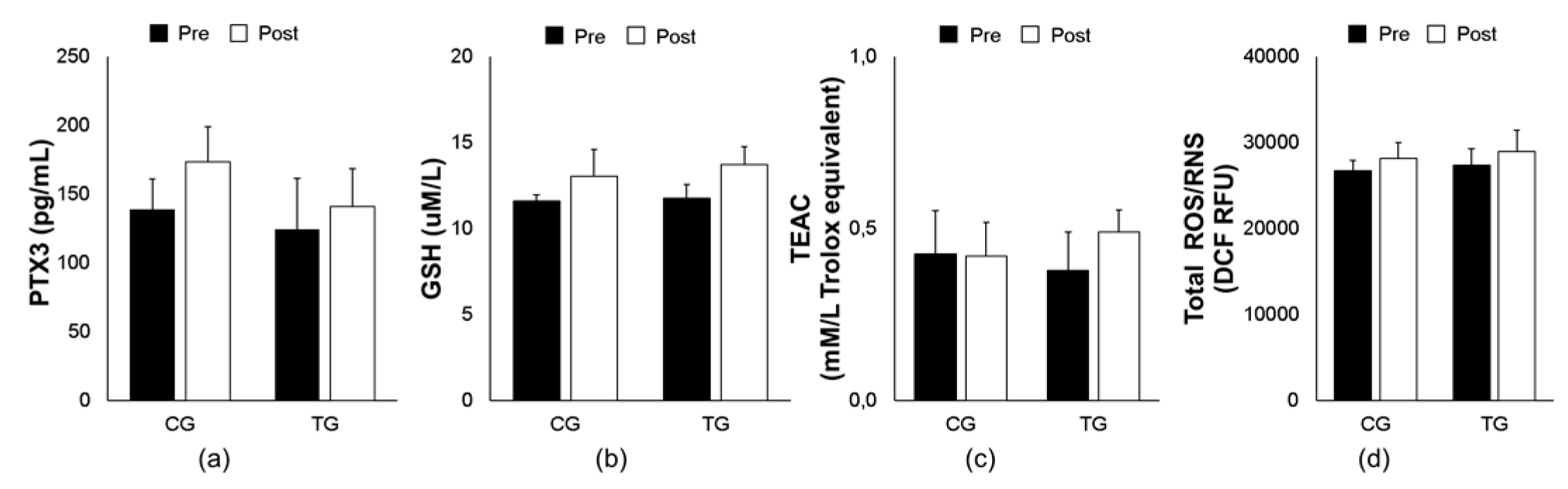
3.2. Effects of Aerobic Training on Inflammatory and Oxidative Stress Responses
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chen, F.; Liu, Y.; Wong, N.-K.; Xiao, J.; So, K.-F. Oxidative stress in stem cell aging. Cell Transplant. 2017, 26, 1483–1495. [Google Scholar] [CrossRef]
- Han, E.-S.; Muller, F.L.; Pérez, V.I.; Qi, W.; Liang, H.; Xi, L.; Fu, C.; Doyle, E.; Hickey, M.; Cornell, J.; et al. The in vivo gene expression signature of oxidative stress. Physiol. Genom. 2008, 34, 112–126. [Google Scholar] [CrossRef]
- Lelkes, P.I.; Hahn, K.A.; Karmiol, S.; Schmidt, D.H. Hypoxia/reoxygenation enhances tube formation of cultured human microvascular endothelial cells: The role of reactive oxygen species. In Angiogenesis; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 1998; pp. 321–336. [Google Scholar]
- Ivanov, A.V.; Valuev-Elliston, V.T.; Ivanova, O.N.; Kochetkov, S.N.; Starodubova, E.S.; Bartosch, B.; Isaguliants, M.G. Oxidative stress during HIV infection: Mechanisms and consequences. Oxidative Med. Cell. Longev. 2016, 2016, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Bullone, M.; Lavoie, J.-P. The contribution of oxidative stress and inflamm-aging in human and equine asthma. Int. J. Mol. Sci. 2017, 18, 2612. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Zhang, J.; Li, H. Selenium, aging and aging-related diseases. Aging Clin. Exp. Res. 2019, 31, 1035–1047. [Google Scholar] [CrossRef] [PubMed]
- Slusher, A.L.; Whitehurst, M.; Maharaj, A.; Dodge, K.M.; Fico, B.G.; Mock, J.T.; Huang, C.-J. Plasma pentraxin 3 and glucose kinetics following acute high-intensity interval exercise versus continuous moderate-intensity exercise in healthy men. Appl. Physiol. Nutr. Metab. 2018, 43, 1233–1238. [Google Scholar] [CrossRef]
- Alles, V.V.; Bottazzi, B.; Peri, G.; Golay, J.; Introna, M.; Mantovani, A. Inducible expression of PTX3, a new member of the pentraxin family, in human mononuclear phagocytes. Blood 1994, 84, 3483–3493. [Google Scholar] [CrossRef]
- Bozza, S.; Campo, S.; Arseni, B.; Inforzato, A.; Ragnar, L.; Bottazzi, B.; Mantovani, A.; Moretti, S.; Oikonomous, V.; De Santis, R.; et al. PTX3 binds MD-2 and promotes TRIF-dependent immune protection in aspergillosis. J. Immunol. 2014, 193, 2340–2348. [Google Scholar] [CrossRef]
- Wang, L.; Cano, M.; Datta, S.; Wei, H.; Ebrahimi, K.B.; Gorashi, Y.; Garlanda, C.; Handa, J.T. Pentraxin 3 recruits complement factor H to protect against oxidative stress-induced complement and inflammasome overactivation. J. Pathol. 2016, 240, 495–506. [Google Scholar] [CrossRef]
- Balci, Y.I.; Nuray, E.; Polat, A.; Enli, Y.; Ozgurler, F.; Akin, M.; Balcı, Y.I.; Akın, M. Pentraxin-3 levels in beta thalassemia major and minor patients, and its relationship with antioxidant capacity and total oxidant Stress. J. Pediatr. Hematol. 2016, 38, 1–16. [Google Scholar]
- Lee, R.; Shin, M.-H.; Kim, H.-N.; Lee, Y.-H.; Choi, S.-W.; Ahn, H.-R.; Kweon, S.-S. Relationship between plasma pentraxin 3 level and risk of chronic kidney disease in the Korean elderly: The Dong-gu study. Int. Urol. Nephrol. 2017, 49, 2027–2033. [Google Scholar] [CrossRef]
- Rodriguez-Miguelez, P.; Fernandez-Gonzalo, R.; Almar, M.; Mejías, Y.; Rivas, A.; De Paz, J.A.; Cuevas, M.J.; González-Gallego, J. Role of Toll-like receptor 2 and 4 signaling pathways on the inflammatory response to resistance training in elderly subjects. AGE 2014, 36, 9734. [Google Scholar] [CrossRef]
- Fujie, S.; Hasegawa, N.; Sanada, K.; Hamaoka, T.; Maeda, S.; Padilla, J.; Martinez-Lemus, L.A.; Iemitsu, M. Increased serum salusin-α by aerobic exercise training correlates with improvements in arterial stiffness in middle-aged and older adults. Aging 2020. [Google Scholar] [CrossRef] [PubMed]
- Naseeb, M.A.; Volpe, S.L. Protein and exercise in the prevention of sarcopenia and aging. Nutr. Res. 2017, 40, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Northey, J.M.; Cherbuin, N.; Pumpa, K.L.; Smee, D.J.; Rattray, B. Exercise interventions for cognitive function in adults older than 50: A systematic review with meta-analysis. Br. J. Sports Med. 2018, 52, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Ross, M.D. Aging—How Lifestyle changes could prove to be an effective medicine for the aging cardiovascular system. Curr. Cardiol. Rev. 2018, 14, 225–226. [Google Scholar] [CrossRef]
- Paillard, T.; Rolland, Y.; Barreto, P.D.S. Protective effects of physical exercise in Alzheimer’s disease and Parkinson’s disease: A narrative review. J. Clin. Neurol. 2015, 11, 212–219. [Google Scholar] [CrossRef]
- Radak, Z.; Torma, F.; Berkes, I.; Goto, S.; Mimura, T.; Posa, A.; Balogh, L.; Boldogh, I.; Suzuki, K.; Higuchi, M.; et al. Exercise effects on physiological function during aging. Free. Radic. Boil. Med. 2019, 132, 33–41. [Google Scholar] [CrossRef]
- Mejías-Peña, Y.; Rodriguez-Miguelez, P.; Fernandez-Gonzalo, R.; Martínez-Flórez, S.; Almar, M.; De Paz, J.A.; Cuevas, M.J.; González-Gallego, J. Effects of aerobic training on markers of autophagy in the elderly. AGE 2016, 38, 33. [Google Scholar] [CrossRef]
- Mejías-Peña, Y.; Estébanez, B.; Rodriguez-Miguelez, P.; Fernandez-Gonzalo, R.; Almar, M.; De Paz, J.A.; González-Gallego, J.; Cuevas, M.J. Impact of resistance training on the autophagy-inflammation-apoptosis crosstalk in elderly subjects. Aging 2017, 9, 408–418. [Google Scholar] [CrossRef]
- Estébanez, B.; Moreira, O.C.; Almar, M.; De Paz, J.A.; Gonzalez-Gallego, J.; Cuevas, M.J. Effects of a resistance-training programme on endoplasmic reticulum unfolded protein response and mitochondrial functions in PBMCs from elderly subjects. Eur. J. Sport Sci. 2019, 19, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Moreira, O.C.; Estébanez, B.; Martínez-Florez, S.; De Paz, J.A.; Cuevas, M.J.; González-Gallego, J. Mitochondrial function and mitophagy in the elderly: Effects of exercise. Oxidative Med. Cell. Longev. 2017, 2017, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Estébanez, B.; De Paz, J.A.; Cuevas, M.J.; González-Gallego, J. Endoplasmic reticulum unfolded protein response, aging and exercise: An update. Front. Physiol. 2018, 9, 1744. [Google Scholar] [CrossRef] [PubMed]
- Fatouros, I.G.; Jamurtas, A.Z.; Villiotou, V.; Pouliopoulou, S.; Fotinakis, P.; Taxildaris, K.; Deliconstantinos, G. Oxidative stress responses in older men during endurance training and detraining. Med. Sci. Sports Exerc. 2004, 36, 2065–2072. [Google Scholar] [CrossRef]
- Park, S.-Y.; Kwak, Y.-S. Impact of aerobic and anaerobic exercise training on oxidative stress and antioxidant defense in athletes. J. Exerc. Rehabilitation 2016, 12, 113–118. [Google Scholar] [CrossRef]
- Bouzid, M.A.; Filaire, E.; Matran, R.; Robin, S.; Fabre, C. Lifelong voluntary exercise modulates age-related changes in oxidative stress. Int. J. Sports Med. 2018, 39, 21–28. [Google Scholar] [CrossRef]
- Huang, C.-J.; Webb, H.E.; Beasley, K.N.; McAlpine, D.A.; Tangsilsat, S.E.; Acevedo, E.O. Cardiorespiratory fitness does not alter plasma pentraxin 3 and cortisol reactivity to acute psychological stress and exercise. Appl. Physiol. Nutr. Metab. 2014, 39, 375–380. [Google Scholar] [CrossRef]
- Slusher, A.L.; Zúñiga, T.M.; Acevedo, E.O. Aerobic fitness alters the capacity of mononuclear cells to produce pentraxin 3 following maximal exercise. Graefe’s Arch. Clin. Exp. Ophthalmol. 2018, 118, 1515–1526. [Google Scholar] [CrossRef]
- Slusher, A.L.; Mock, J.T.; Whitehurst, M.; Maharaj, A.; Huang, C.-J. The impact of obesity on pentraxin 3 and inflammatory milieu to acute aerobic exercise. Metab. 2015, 64, 323–329. [Google Scholar] [CrossRef]
- Slusher, A.L.; Huang, C.-J. Association of pentraxin 3 with insulin resistance and glucose response following maximal aerobic exercise in obese and normal-mass individuals. Can. J. Physiol. Pharmacol. 2016, 94, 734–738. [Google Scholar] [CrossRef]
- Slusher, A.L.; Shibata, Y.; Whitehurst, M.; Maharaj, A.; Quiles, J.M.; Huang, C.-J. Exercise reduced pentraxin 3 levels produced by endotoxin-stimulated human peripheral blood mononuclear cells in obese individuals. Exp. Boil. Med. 2017, 242, 1279–1286. [Google Scholar] [CrossRef] [PubMed]
- Miyaki, A.; Maeda, S.; Choi, Y.; Akazawa, N.; Tanabe, Y.; Ajisaka, R. Habitual aerobic exercise increases plasma pentraxin 3 levels in middle-aged and elderly women. Appl. Physiol. Nutr. Metab. 2012, 37, 907–911. [Google Scholar] [CrossRef] [PubMed]
- Zempo-Miyaki, A.; Fujie, S.; Sato, K.; Hasegawa, N.; Sanada, K.; Maeda, S.; Hamaoka, T.; Iemitsu, M. Elevated pentraxin 3 level at the early stage of exercise training is associated with reduction of arterial stiffness in middle-aged and older adults. J. Hum. Hypertens. 2016, 30, 521–526. [Google Scholar] [CrossRef] [PubMed]
- McFarlin, B.K.; Flynn, M.G.; Campbell, W.W.; Craig, B.A.; Robinson, J.P.; Stewart, L.K.; Timmerman, K.L.; Coen, P.M. Physical activity status, but not age, influences inflammatory biomarkers and toll-like receptor 4. J. Gerontol. Ser. A Boil. Sci. Med Sci. 2006, 61, 388–393. [Google Scholar] [CrossRef]
- Stewart, L.K.; Flynn, M.G.; Campbell, W.W.; Craig, B.A.; Robinson, J.P.; McFarlin, B.K.; Timmerman, K.L.; Coen, P.M.; Felker, J.; Talbert, E. Influence of exercise training and age on CD14+ cell-surface expression of toll-like receptor 2 and 4. Brain Behav. Immun. 2005, 19, 389–397. [Google Scholar] [CrossRef]
- Rodriguez-Miguelez, P.; Fernandez-Gonzalo, R.; Collado, P.S.; Almar, M.; Martínez-Flórez, S.; De Paz, J.A.; González-Gallego, J.; Cuevas, M.J. Whole-body vibration improves the anti-inflammatory status in elderly subjects through toll-like receptor 2 and 4 signaling pathways. Mech. Ageing Dev. 2015, 150, 12–19. [Google Scholar] [CrossRef]
- Farinha, J.B.; Steckling, F.M.; Stefanello, S.T.; Cardoso, M.S.; Nunes, L.S.; Barcelos, R.P.; Duarte, T.; Kretzmann, N.A.; Mota, C.B.; Bresciani, G.; et al. Response of oxidative stress and inflammatory biomarkers to a 12-week aerobic exercise training in women with metabolic syndrome. Sports Med. Open 2015, 1, 1640. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Markers of oxidative stress in erythrocytes and plasma during aging in humans. Oxidative Med. Cell. Longev. 2010, 3, 2–12. [Google Scholar] [CrossRef]
- Fisher-Wellman, K.; Bloomer, R.J. Acute exercise and oxidative stress: A 30 year history. Dyn. Med. 2009, 8, 1. [Google Scholar] [CrossRef]
- Krause, M.; Rodrigues-Krause, J.; O’Hagan, C.; Medlow, P.; Davison, G.; Susta, D.; Boreham, C.; Newsholme, P.; O’Donnell, M.; Murphy, C.; et al. The effects of aerobic exercise training at two different intensities in obesity and type 2 diabetes: Implications for oxidative stress, low-grade inflammation and nitric oxide production. Eur. J. Appl. Physiol. 2014, 114, 251–260. [Google Scholar] [CrossRef]
- Samjoo, I.A.; Safdar, A.; Hamadeh, M.J.; Raha, S.; Tarnopolsky, M.A. The effect of endurance exercise on both skeletal muscle and systemic oxidative stress in previously sedentary obese men. Nutr. Diabetes 2013, 3, e88. [Google Scholar] [CrossRef] [PubMed]
- Campbell, P.T.; Gross, M.D.; Potter, J.D.; Schmitz, K.H.; Duggan, C.; McTiernan, A.; Ulrich, C.M. Effect of exercise on oxidative stress: A 12-month randomized, controlled trial. Med. Sci. Sports Exerc. 2010, 42, 1448–1453. [Google Scholar] [CrossRef] [PubMed]
- Simar, D.; Malatesta, D.; Mas, E.; Delage, M.; Caillaud, C. Effect of an 8-weeks aerobic training program in elderly on oxidative stress and HSP72 expression in leukocytes during antioxidant supplementation. J. Nutr. Heal. Aging 2012, 16, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Done, A.J.; Traustadóttir, T. Aerobic exercise increases resistance to oxidative stress in sedentary older middle-aged adults. A pilot study. AGE 2016, 38, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Krzanowski, M.; Krzanowska, K.; Gajda, M.; Dumnicka, P.; Dziewierz, A.; Woziwodzka, K.; Litwin, J.A.; Sułowicz, W. Pentraxin 3 as a new indicator of cardiovascular-related death in patients with advanced chronic kidney disease. Pol. Arch. Intern. Med. 2017, 127, 170–177. [Google Scholar] [PubMed]
- Fernandez-Gonzalo, R.; De Paz, J.A.; Rodriguez-Miguelez, P.; Cuevas, M.J.; González-Gallego, J. TLR4-mediated blunting of inflammatory responses to eccentric exercise in young women. Mediat. Inflamm. 2014, 2014, 1–11. [Google Scholar] [CrossRef]
- Fernandez-Gonzalo, R.; De Paz, J.A.; Rodriguez-Miguelez, P.; Cuevas, M.J.; González-Gallego, J. Effects of eccentric exercise on toll-like receptor 4 signaling pathway in peripheral blood mononuclear cells. J. Appl. Physiol. 2012, 112, 2011–2018. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Variable | CG (n = 5) | TG (n = 9) | p Value |
|---|---|---|---|
| Age (years) | 70.79 ± 1.66 | 68.67 ± 1.25 | 0.330 |
| Height (cm) | 160.60 ± 0.03 | 159.50 ± 0.034 | 0.825 |
| Weight (kg) | 67.00 ± 4.36 | 65.63 ± 3.87 | 0.828 |
| BMI (kg/m2) | 25.87 ± 1.07 | 25.71 ± 0.90 | 0.914 |
| VO2peak (mL/kg/min) | 30.37 ± 1.44 | 31.02 ± 1.58 | 0.789 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Estébanez, B.; Rodriguez, A.L.; Visavadiya, N.P.; Whitehurst, M.; Cuevas, M.J.; González-Gallego, J.; Huang, C.-J. Aerobic Training Down-Regulates Pentraxin 3 and Pentraxin 3/Toll-Like Receptor 4 Ratio, Irrespective of Oxidative Stress Response, in Elderly Subjects. Antioxidants 2020, 9, 110. https://doi.org/10.3390/antiox9020110
Estébanez B, Rodriguez AL, Visavadiya NP, Whitehurst M, Cuevas MJ, González-Gallego J, Huang C-J. Aerobic Training Down-Regulates Pentraxin 3 and Pentraxin 3/Toll-Like Receptor 4 Ratio, Irrespective of Oxidative Stress Response, in Elderly Subjects. Antioxidants. 2020; 9(2):110. https://doi.org/10.3390/antiox9020110
Chicago/Turabian StyleEstébanez, Brisamar, Alexandra L. Rodriguez, Nishant P. Visavadiya, Michael Whitehurst, María J. Cuevas, Javier González-Gallego, and Chun-Jung Huang. 2020. "Aerobic Training Down-Regulates Pentraxin 3 and Pentraxin 3/Toll-Like Receptor 4 Ratio, Irrespective of Oxidative Stress Response, in Elderly Subjects" Antioxidants 9, no. 2: 110. https://doi.org/10.3390/antiox9020110
APA StyleEstébanez, B., Rodriguez, A. L., Visavadiya, N. P., Whitehurst, M., Cuevas, M. J., González-Gallego, J., & Huang, C.-J. (2020). Aerobic Training Down-Regulates Pentraxin 3 and Pentraxin 3/Toll-Like Receptor 4 Ratio, Irrespective of Oxidative Stress Response, in Elderly Subjects. Antioxidants, 9(2), 110. https://doi.org/10.3390/antiox9020110