Effects of Two-Week Betaine Supplementation on Apoptosis, Oxidative Stress, and Aerobic Capacity after Exhaustive Endurance Exercise

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experimental Design
2.3. Supplementation Protocol
2.4. Graded Exercise Test
2.5. Blood Sampling and Analysis
2.5.1. Lymphocyte Apoptosis
2.5.2. MTP Decline in Lymphocyte
2.5.3. Concentrations of Plasma Betaine and Choline
2.5.4. TBARS Concentration
2.5.5. PC Concentration
2.6. Statistical Analysis
3. Results
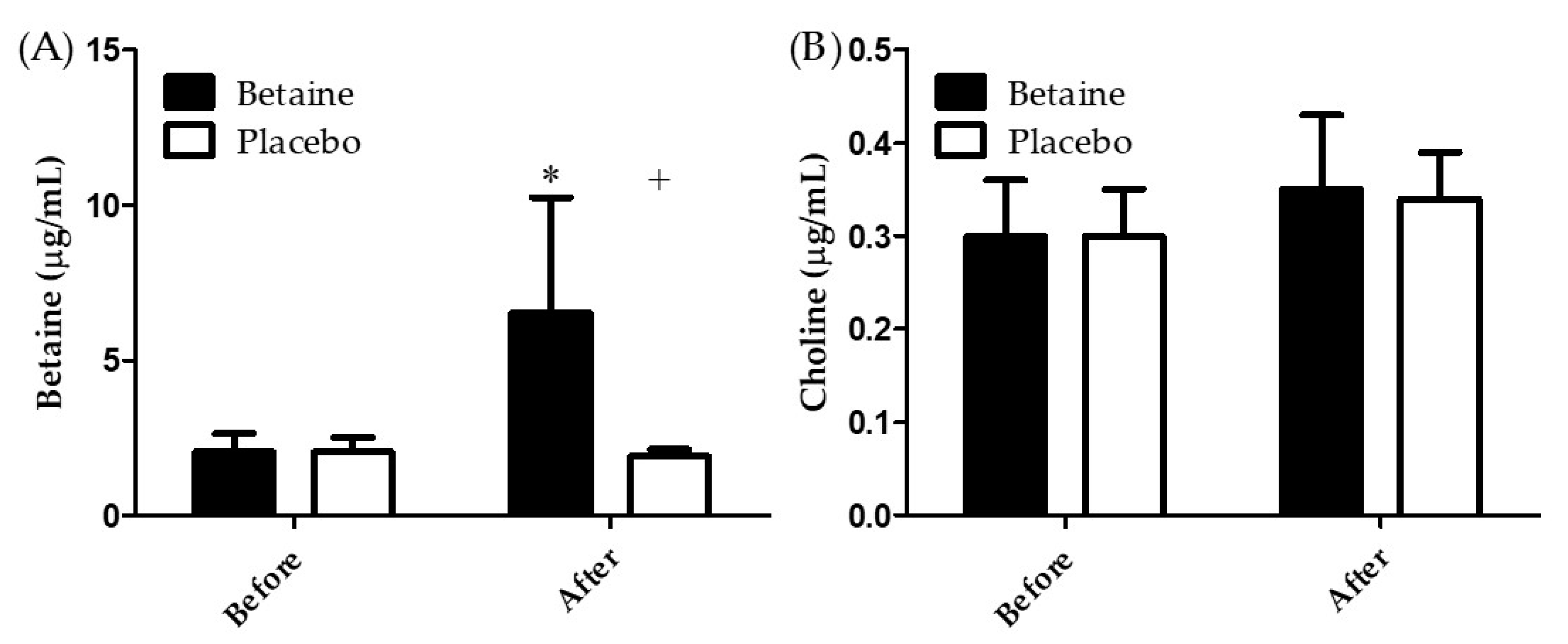
3.1. Blood Concentrations of Betaine and Coline
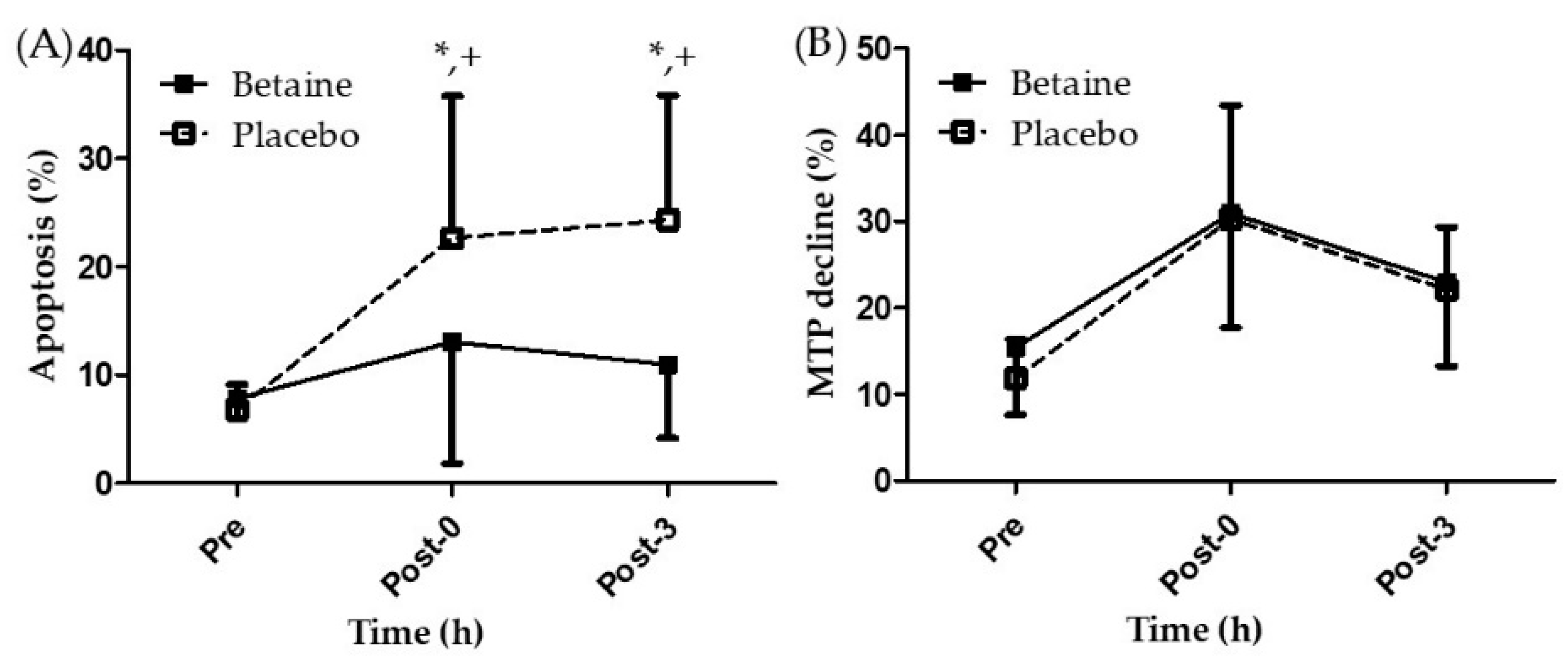
3.2. Effects of Betaine Supplementation on Lymphocyte Apoptosis and MTP Decline
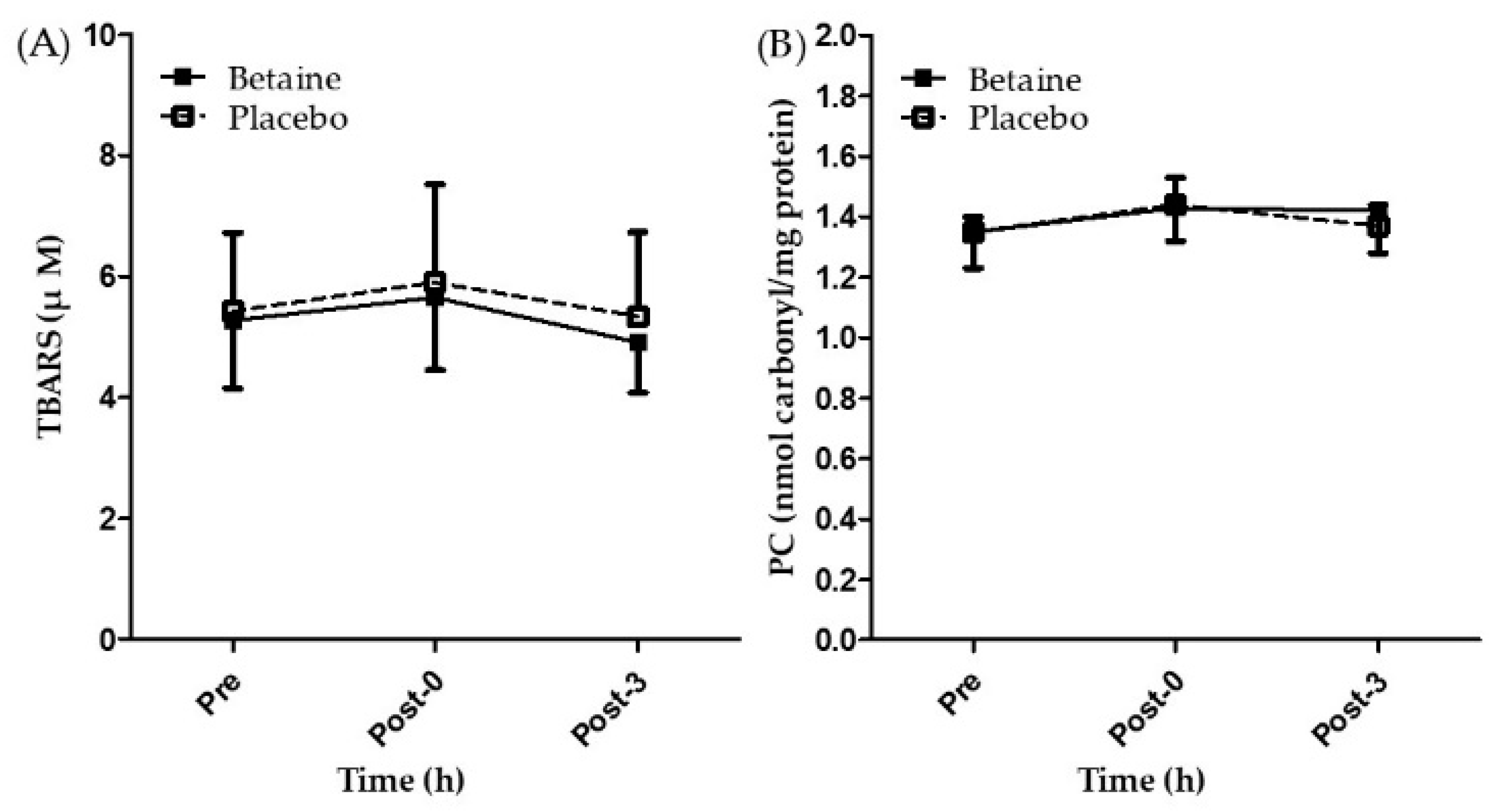
3.3. Effect of Betaine Supplementation on Oxidative Stress
3.4. Effect of Betaine Supplementation on Aerobic Capacity
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Aslani, B.A.; Ghobadi, S. Studies on oxidants and antioxidants with a brief glance at their relevance to the immune system. Life Sci. 2016, 146, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Jornot, L.; Petersen, H.; Junod, A.F. Hydrogen peroxide-induced DNA damage is independent of nuclear calcium but dependent on redox-active ions. Biochem. J. 1998, 335, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Kono, Y.; Fridovich, I. Superoxide radical inhibits catalase. J. Biol. Chem. 1982, 257, 5751–5754. [Google Scholar] [PubMed]
- Clarkson, P.M.; Thompson, H.S. Antioxidants: What role do they play in physical activity and health? Am. J. Clin. Nutr. 2000, 72, 637S–646S. [Google Scholar] [CrossRef] [PubMed]
- Finaud, J.; Lac, G.; Filaire, E. Oxidative stress: Relationship with exercise and training. Sports Med. 2006, 36, 327–358. [Google Scholar] [CrossRef]
- Phaneuf, S.; Leeuwenburgh, C. Apoptosis and exercise. Med. Sci. Sports Exerc. 2001, 33, 393–396. [Google Scholar] [CrossRef]
- Mooren, F.C.; Blöming, D.; Lechtermann, A.; Lerch, M.M.; Völker, K. Lymphocyte apoptosis after exhaustive and moderate exercise. J. Appl. Physiol. 2002, 93, 147–153. [Google Scholar] [CrossRef]
- Ådén, J.; Mushtaq, A.U.; Dingeldein, A.; Wallgren, M.; Gröbner, G. A novel recombinant expression and purification approach for the full-length anti-apoptotic membrane protein Bcl-2. Protein Expr. Purif. 2020, 172, 105628. [Google Scholar] [CrossRef]
- Tuan, T.C.; Hsu, T.G.; Fong, M.C.; Hsu, C.F.; Tsai, K.K.; Lee, C.Y.; Kong, C.W. Deleterious effects of short-term, high-intensity exercise on immune function: Evidence from leucocyte mitochondrial alterations and apoptosis. Br. J. Sports Med. 2008, 42, 11–15. [Google Scholar] [CrossRef]
- Townsend, J.R.; Stout, J.R.; Jajtner, A.R.; Church, D.D.; Beyer, K.S.; Riffe, J.J.; Muddle, T.W.D.; Herrlinger, K.L.; Fukuda, D.H.; Hoffman, J.R. Polyphenol supplementation alters intramuscular apoptotic signaling following acute resistance exercise. Physiol. Rep. 2018, 6, e13552. [Google Scholar] [CrossRef]
- Shakeri, M.; Cottrell, J.J.; Wilkinson, S.; Le, H.H.; Suleria, H.A.R.; Warner, R.D.; Dunshea, F.R. Growth performance and characterization of meat quality of broiler chickens supplemented with betaine and antioxidants under cyclic heat stress. Antioxidants 2019, 8, 336. [Google Scholar] [CrossRef] [PubMed]
- Craig, S.A. Betaine in human nutrition. Am. J. Clin. Nutr. 2004, 80, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; He, F.; Wu, C.; Li, P.; Li, N.; Deng, J.; Zhu, G.; Ren, W.; Peng, Y. Betaine in inflammation: Mechanistic aspects and applications. Front. Immunol. 2018, 9, 1070. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Chen, X.; Jiang, X.; Niu, J.; Cui, C.; Chen, Z.; Sun, J. Betaine ameliorates prenatal valproic-acid-induced autism-like behavioral abnormalities in mice by promoting homocysteine metabolism. Psychiatry Clin. Neurosci. 2019, 73, 317–322. [Google Scholar] [CrossRef]
- Saeed, M.; Babazadeh, D.; Naveed, M.; Arain, M.A.; Hassan, F.U.; Chao, S. Reconsidering betaine as a natural anti-heat stress agent in poultry industry: A review. Trop. Anim. Health Prod. 2017, 49, 1329–1338. [Google Scholar] [CrossRef]
- Dragolovich, J. Dealing with salt stress in animal cells: The role and regulation of glycine betaine concentrations. J. Exp. Zool. 1994, 268, 139–144. [Google Scholar] [CrossRef]
- Zeisel, S.H.; Growdon, J.H.; Wurtman, R.J.; Magil, S.G.; Logue, M. Normal plasma choline responses to ingested lecithin. Neurology 1980, 30, 1226–1229. [Google Scholar] [CrossRef]
- Flower, R.J.; Pollitt, R.J.; Sanford, P.A.; Smyth, D.H. Metabolism and transfer of choline in hamster small intestine. J. Physiol. 1972, 226, 473–489. [Google Scholar] [CrossRef]
- Fernández-Fígares, I.; Wray-Cahen, D.; Steele, N.C.; Campbell, R.G.; Hall, D.D.; Virtanen, E.; Caperna, T.J. Effect of dietary betaine on nutrient utilization and partitioning in the young growing feed-restricted pig. J. Anim. Sci. 2002, 80, 421–428. [Google Scholar] [CrossRef]
- Matthews, J.O.; Southern, L.L.; Pontif, J.E.; Higbie, A.D.; Bidner, T.D. Interactive effects of betaine, crude protein, and net energy in finishing pigs. J. Anim. Sci. 1998, 76, 2444–2455. [Google Scholar] [CrossRef]
- Waldenstedt, L.; Elwinger, K.; Thebo, P.; Uggla, A. Effect of betaine supplement on broiler performance during an experimental coccidial infection. Poult. Sci. 1999, 78, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, L.E.; Casa, D.J.; Roti, M.W.; Lee, E.C.; Craig, S.A.; Sutherland, J.W.; Fiala, K.A.; Maresh, C.M. Influence of betaine consumption on strenuous running and sprinting in a hot environment. J. Strength Cond. Res. 2008, 22, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Apicella, J.M.; Lee, E.C.; Bailey, B.L.; Saenz, C.; Anderson, J.M.; Craig, S.A.; Kraemer, W.J.; Volek, J.S.; Maresh, C.M. Betaine supplementation enhances anabolic endocrine and Akt signaling in response to acute bouts of exercise. Eur. J. Appl. Physiol. 2013, 113, 793–802. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.C.; Maresh, C.M.; Kraemer, W.J.; Yamamoto, L.M.; Hatfield, D.L.; Bailey, B.L.; Armstrong, L.E.; Volek, J.S.; McDermott, B.P.; Craig, S.A. Ergogenic effects of betaine supplementation on strength and power performance. J. Int. Soc. Sports Nutr. 2010, 7, 27. [Google Scholar] [CrossRef]
- Hoffman, J.R.; Ratamess, N.A.; Kang, J.; Gonzalez, A.M.; Beller, N.A.; Craig, S.A. Effect of 15 days of betaine ingestion on concentric and eccentric force outputs during isokinetic exercise. J. Strength Cond. Res. 2011, 25, 2235–2241. [Google Scholar] [CrossRef]
- Veskovic, M.; Mladenovic, D.; Milenkovic, M.; Tosic, J.; Borozan, S.; Gopcevic, K.; Labudovic-Borovic, M.; Dragutinovic, V.; Vucevic, D.; Jorgacevic, B.; et al. Betaine modulates oxidative stress, inflammation, apoptosis, autophagy, and Akt/mTOR signaling in methionine-choline deficiency-induced fatty liver disease. Eur. J. Pharmacol. 2019, 848, 39–48. [Google Scholar] [CrossRef]
- Garrett, Q.; Khandekar, N.; Shih, S.; Flanagan, J.L.; Simmons, P.; Vehige, J.; Willcox, M.D. Betaine stabilizes cell volume and protects against apoptosis in human corneal epithelial cells under hyperosmotic stress. Exp. Eye Res. 2013, 108, 33–41. [Google Scholar] [CrossRef]
- Giris, M.; Dogru-Abbasoglu, S.; Soluk-Tekkesin, M.; Olgac, V.; Uysal, M. Effect of betaine treatment on the regression of existing hepatic triglyceride accumulation and oxidative stress in rats fed on high fructose diet. Gen. Physiol. Biophys. 2018, 37, 563–570. [Google Scholar] [CrossRef]
- Heidari, R.; Niknahad, H.; Sadeghi, A.; Mohammadi, H.; Ghanbarinejad, V.; Ommati, M.M.; Hosseini, A.; Azarpira, N.; Khodaei, F.; Farshad, O.; et al. Betaine treatment protects liver through regulating mitochondrial function and counteracting oxidative stress in acute and chronic animal models of hepatic injury. Biomed. Pharmacother. 2018, 103, 75–86. [Google Scholar] [CrossRef]
- Yang, M.T.; Lin, S.C.; Chien, L.H.; Chan, K.H. Effects of acute betaine supplementation on apoptosis, oxidative stress and aerobic capacity after exhaustive endurance exercise. Sports Exerc. Res. 2018, 20, 373–383. [Google Scholar] [CrossRef]
- Ho, C.F.; Shih, C.Y.; Chan, K.H.; Wang, T.Y. Effect of uphill high-intensity interval training on aerobic capacity and power of lower extremity in basketball players. Sports Exerc. Res. 2012, 14, 476–482. [Google Scholar] [CrossRef]
- Bruce, S.J.; Guy, P.A.; Rezzi, S.; Ross, A.B. Quantitative measurement of betaine and free choline in plasma, cereals and cereal products by isotope dilution LC-MS/MS. J. Agric. Food Chem. 2010, 58, 2055–2061. [Google Scholar] [CrossRef] [PubMed]
- Keles, M.S.; Taysi, S.; Sen, N.; Aksoy, H.; Akcay, F. Effect of corticosteroid therapy on serum and CSF malondialdehyde and antioxidant proteins in multiple sclerosis. Can. J. Neurol. Sci. 2001, 28, 141–143. [Google Scholar] [CrossRef] [PubMed]
- Boadi, W.Y.; Iyere, P.A.; Adunyah, S.E. Effect of Quercetin and Genistein on Copper- And Iron-Induced Lipid Peroxidation in Methyl Linolenate. J. Appl. Toxicol. 2003, 23, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Margaritelis, N.V.; Theodorou, A.A.; Paschalis, V.; Veskoukis, A.S.; Dipla, K.; Zafeiridis, A.; Panayiotou, G.; Vrabas, I.S.; Kyparos, A.; Nikolaidis, M.G. Adaptations to endurance training depend on exercise-induced oxidative stress: Exploiting redox interindividual variability. Acta Physiol. 2018, 222. [Google Scholar] [CrossRef] [PubMed]
- Moser, C.C.; Farid, T.A.; Chobot, S.E.; Dutton, P.L. Electron tunneling chains of mitochondria. Biochim. Biophys. Acta 2006, 1757, 1096–1109. [Google Scholar] [CrossRef]
- Nilsson, M.I.; Tarnopolsky, M.A. Mitochondria and aging-the role of exercise as a countermeasure. Biology 2019, 8, 40. [Google Scholar] [CrossRef]
- Andreotti, D.Z.; Silva, J.D.N.; Matumoto, A.M.; Orellana, A.M.; de Mello, P.S.; Kawamoto, E.M. Effects of physical exercise on autophagy and apoptosis in aged brain: Human and animal studies. Front. Nutr. 2020, 7, 94. [Google Scholar] [CrossRef]
- Bratton, S.B.; MacFarlane, M.; Cain, K.; Cohen, G.M. Protein complexes activate distinct caspase cascades in death receptor and stress-induced apoptosis. Exp. Cell Res. 2000, 256, 27–33. [Google Scholar] [CrossRef]
- van Loo, G.; Saelens, X.; van Gurp, M.; MacFarlane, M.; Martin, S.J.; Vandenabeele, P. The role of mitochondrial factors in apoptosis: A Russian roulette with more than one bullet. Cell Death Differ. 2002, 9, 1031–1042. [Google Scholar] [CrossRef]
- del Favero, S.; Roschel, H.; Artioli, G.; Ugrinowitsch, C.; Tricoli, V.; Costa, A.; Barroso, R.; Negrelli, A.L.; Otaduy, M.C.; da Costa Leite, C.; et al. Creatine but not betaine supplementation increases muscle phosphorylcreatine content and strength performance. Amino Acids 2012, 42, 2299–2305. [Google Scholar] [CrossRef] [PubMed]
- Pryor, J.L.; Craig, S.A.; Swensen, T. Effect of betaine supplementation on cycling sprint performance. J. Int. Soc. Sports Nutr. 2012, 9, 12. [Google Scholar] [CrossRef] [PubMed]
- Trepanowski, J.F.; Farney, T.M.; McCarthy, C.G.; Schilling, B.K.; Craig, S.A.; Bloomer, R.J. The effects of chronic betaine supplementation on exercise performance, skeletal muscle oxygen saturation and associated biochemical parameters in resistance trained men. J. Strength Cond. Res. 2011, 25, 3461–3471. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Jani, K.K.; Gupta, N. Hypertension: Salt restriction, sodium homeostasis, and other ions. Indian J. Med. Sci. 2011, 65, 121–132. [Google Scholar] [CrossRef]
- Alfieri, R.R.; Cavazzoni, A.; Petronini, P.G.; Bonelli, M.A.; Caccamo, A.E.; Borghetti, A.F.; Wheeler, K.P. Compatible osmolytes modulate the response of porcine endothelial cells to hypertonicity and protect them from apoptosis. J. Physiol. 2002, 540, 499–508. [Google Scholar] [CrossRef]
- Graf, D.; Kurz, A.K.; Reinehr, R.; Fischer, R.; Kircheis, G.; Häussinger, D. Prevention of bile acid-induced apoptosis by betaine in rat liver. Hepatology 2002, 36, 829–839. [Google Scholar] [CrossRef]
- Teocchi, M.A.; D’Souza-Li, L. Apoptosis through death receptors in temporal lobe epilepsy-associated hippocampal sclerosis. Mediators Inflamm. 2016, 2016, 8290562. [Google Scholar] [CrossRef]
- Bingül, İ.; Başaran-Küçükgergin, C.; Aydın, A.F.; Çoban, J.; Doğan-Ekici, I.; Doğru-Abbasoğlu, S.; Uysal, M. Betaine treatment decreased oxidative stress, inflammation, and stellate cell activation in rats with alcoholic liver fibrosis. Environ. Toxicol. Pharmacol. 2016, 45, 170–178. [Google Scholar] [CrossRef]
- Hagar, H.; Medany, A.E.; Salam, R.; Medany, G.E.; Nayal, O.A. Betaine supplementation mitigates cisplatin-induced nephrotoxicity by abrogation of oxidative/nitrosative stress and suppression of inflammation and apoptosis in rats. Exp. Toxicol. Pathol. 2015, 67, 133–141. [Google Scholar] [CrossRef]
- Alirezaei, M. Betaine protects cerebellum from oxidative stress following levodopa and benserazide administration in rats. Iran. J. Basic Med. Sci. 2015, 18, 950–957. [Google Scholar]
- Guo, Y.; Xu, L.S.; Zhang, D.; Liao, Y.P.; Wang, H.P.; Lan, Z.H.; Guan, W.J.; Liu, C.Q. Betaine Effects on Morphology, Proliferation, and p53-induced Apoptosis of HeLa Cervical Carcinoma Cells in Vitro. Asian Pac. J. Cancer Prev. 2015, 16, 3195–3201. [Google Scholar] [CrossRef] [PubMed]
- Harisa, G.I. Oxidative stress and paraoxonase activity in experimental selenosis: Effects of betaine administration. Biol. Trace Elem. Res. 2013, 152, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.Y.; Jung, Y.S.; Kim, S.J.; Park, H.K.; Park, J.H.; Kim, Y.C. Impaired sulfur-amino acid metabolism and oxidative stress in nonalcoholic fatty liver are alleviated by betaine supplementation in rats. J. Nutr. 2009, 139, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. The antioxidant paradox: Less paradoxical now? Br. J. Clin. Pharmacol. 2013, 75, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Whiteman, M. Measuring reactive species and oxidative damage in vivo and in cell culture: How should you do it and what do the results mean? Br. J. Pharmacol. 2004, 142, 231–255. [Google Scholar] [CrossRef]
- Ganesan, B.; Anandan, R.; Lakshmanan, P.T. Studies on the protective effects of betaine against oxidative damage during experimentally induced restraint stress in Wistar albino rats. Cell Stress Chaperones 2011, 16, 641–652. [Google Scholar] [CrossRef]
- Barstow, T.J. Characterization of VO2 kinetics during heavy exercise. Med. Sci. Sports Exerc. 1994, 26, 1327–1334. [Google Scholar] [CrossRef]
- Schwab, U.; Törrönen, A.; Toppinen, L.; Alfthan, G.; Saarinen, M.; Aro, A.; Uusitupa, M. Betaine supplementation decreases plasma homocysteine concentrations but does not affect body weight, body composition, or resting energy expenditure in human subjects. Am. J. Clin. Nutr. 2002, 76, 961–967. [Google Scholar] [CrossRef]
- Olthof, M.R.; Verhoef, P. Effects of betaine intake on plasma homocysteine concentrations and consequences for health. Curr. Drug Metab. 2005, 6, 15–22. [Google Scholar] [CrossRef]







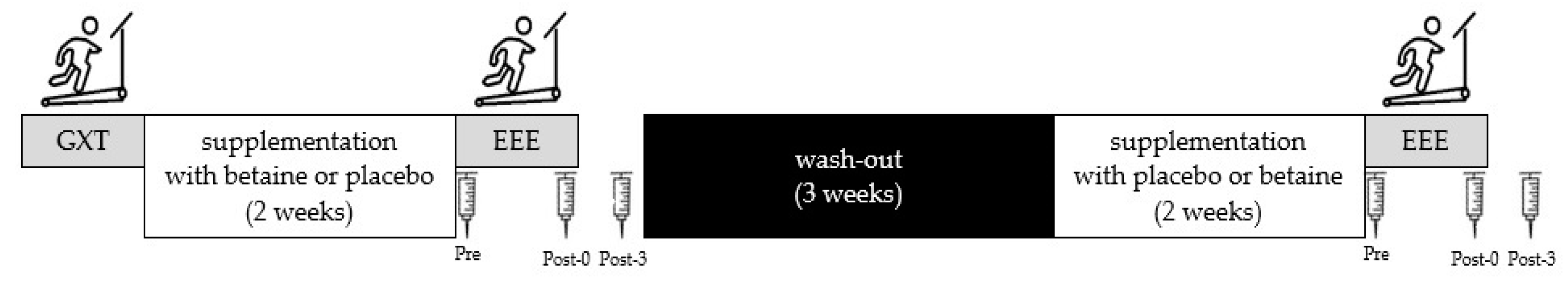{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Parent Ion (m/z) | Molecular Ion (m/z) | DP (V) | FP (V) | EP (V) | CE (V) | CXP (V) |
|---|---|---|---|---|---|---|---|
| choline | 104 | 60 | 28 | 320 | 8 | 26 | 7 |
| d9-choline | 113 | 69 | 35 | 367 | 8.8 | 26 | 8 |
| betaine | 118 | 58 | 35 | 350 | 10 | 38 | 6 |
| d11-betaine | 129 | 66 | 38 | 350 | 8 | 43 | 7 |
| Trial | TTE (min) | HRmax (beats/min) | AHR (beats/min) | VO2peak (mL/kg/min) |
|---|---|---|---|---|
| Betaine | 33.61 ± 2.33 | 195.70 ± 7.06 | 173.33 ± 10.05 | 50.58 ± 6.50 |
| Placebo | 33.32 ± 1.62 | 195.70 ± 7.32 | 173.26 ± 8.99 | 47.40 ± 4.10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, M.-T.; Lee, X.-X.; Huang, B.-H.; Chien, L.-H.; Wang, C.-C.; Chan, K.-H. Effects of Two-Week Betaine Supplementation on Apoptosis, Oxidative Stress, and Aerobic Capacity after Exhaustive Endurance Exercise. Antioxidants 2020, 9, 1189. https://doi.org/10.3390/antiox9121189
Yang M-T, Lee X-X, Huang B-H, Chien L-H, Wang C-C, Chan K-H. Effects of Two-Week Betaine Supplementation on Apoptosis, Oxidative Stress, and Aerobic Capacity after Exhaustive Endurance Exercise. Antioxidants. 2020; 9(12):1189. https://doi.org/10.3390/antiox9121189
Chicago/Turabian StyleYang, Ming-Ta, Xiu-Xin Lee, Bo-Huei Huang, Li-Hui Chien, Chia-Chi Wang, and Kuei-Hui Chan. 2020. "Effects of Two-Week Betaine Supplementation on Apoptosis, Oxidative Stress, and Aerobic Capacity after Exhaustive Endurance Exercise" Antioxidants 9, no. 12: 1189. https://doi.org/10.3390/antiox9121189
APA StyleYang, M.-T., Lee, X.-X., Huang, B.-H., Chien, L.-H., Wang, C.-C., & Chan, K.-H. (2020). Effects of Two-Week Betaine Supplementation on Apoptosis, Oxidative Stress, and Aerobic Capacity after Exhaustive Endurance Exercise. Antioxidants, 9(12), 1189. https://doi.org/10.3390/antiox9121189