Microvesicle Formation Induced by Oxidative Stress in Human Erythrocytes

 ,
,  , ,
, , 

Abstract
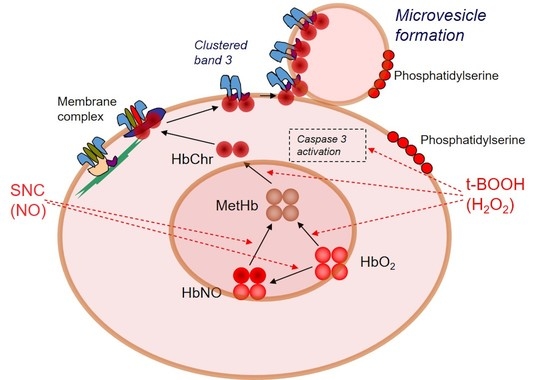
1. Introduction
2. Materials and Methods
2.1. Reagents and Chemicals
2.2. Methods
2.2.1. RBC Preparation
2.2.2. Stress Models
2.2.3. Microparticle Isolation and Analysis
2.2.4. Spectral Analysis of Hemoglobin Species
Hemoglobin Species Calculation
Induction of Hypoxia
2.2.5. Characterization of RBC Deformability by Laser Diffraction Method
Osmotic Fragility Test (OFT)
Ammonium Stress Test (AST)
2.2.6. Flow Cytometry Analysis
Size and Structure Analysis
Esterase Activity Analysis
Phosphatidylserine (PS) Externalization at the RBC Surface
Analysis of Band 3 Clustering
Microparticle Detection
Caspase-3 Activation
2.2.7. Confocal Microscopy
2.2.8. Data Analysis
3. Results
3.1. RBC Viability Strongly Depended on the Ratio of Oxidant to Cell Count
3.2. t-BOOH Induced RBC Vesiculation
3.3. Oxidative Stress Induced Hemoglobin Oxidation to Ferryl (Hemichrome) Forms
3.4. t-BOOH Dose-Dependently Decreased RBC Deformability
3.5. OS Induced RBC Transformation and Microvesicle Formation Was Independent on Extracellular Calcium Concentration
3.6. t-BOOH-Induced OS Triggered Band 3 Clustering
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rosa-Fernandes, L.; Rocha, V.B.; Carregari, V.C.; Urbani, A.; Palmisano, G.A. Perspective on Extracellular Vesicles Proteomics. Front. Chem. 2017, 5, 102. [Google Scholar] [CrossRef] [PubMed]
- Alexandru, N.; Costa, A.; Constantin, A.; Cochior, D.; Georgescu, A. Microparticles: From Biogenesis to Biomarkers and Diagnostic Tools in Cardiovascular Disease. Curr. Stem Cell Res. 2017, 12, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Said, A.S.; Rogers, S.C.; Doctor, A. Physiologic Impact of Circulating RBC Microparticles upon Blood-Vascular Interactions. Front. Physiol. 2018, 8, 1120. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.D.; Bergeron, A.L.; Dong, J.F.; López, J.A. Flow cytometric measurement of microparticles: Pitfalls and protocol modifications. Platelets 2008, 19, 365–372. [Google Scholar] [CrossRef]
- Hattangadi, S.M.; Lodish, H.F. Regulation of erythrocyte lifespan: Do reactive oxygen species set the clock? J. Clin. Investig. 2007, 117, 2075–2077. [Google Scholar] [CrossRef]
- Bosman, G.J.; Lasonder, E.; Luten, M.; Roerdinkholder-Stoelwinder, B.; Novotný, V.M.; Bos, H.; De Grip, W.J. The proteome of red cell membranes and vesicles during storage in blood bank conditions. Transfusion 2008, 48, 827–835. [Google Scholar] [CrossRef]
- Mohanty, J.G.; Nagababu, E.; Rifkind, J.M. Red blood cell oxidative stress impairs oxygen delivery and induces red blood cell aging. Front. Physiol. 2014, 5, 84. [Google Scholar] [CrossRef]
- Bratosin, D.; Estaquier, J.; Petit, F.; Arnoult, D.; Quatannens, B.; Tissier, J.P.; Slomianny, C.; Sartiaux, C.; Alonso, C.; Huart, J.J.; et al. Programmed cell death in mature erythrocytes: A model for investigating death effector pathways operating in the absence of mitochondria. Cell Death Differ. 2001, 8, 1143–1156. [Google Scholar] [CrossRef]
- Nguyen, D.B.; Ly, T.B.; Wesseling, M.C.; Hittinger, M.; Torge, A.; Devitt, A.; Perrie, I.; Bernhardt, I. Characterization of Microvesicles Released from Human Red Blood Cells. Cell Physiol. Biochem. 2016, 38, 1085–1099. [Google Scholar] [CrossRef]
- Lutz, H.U.; Liu, S.C.; Palek, J. Release of spectrin-free vesicles from human erythrocytes during ATP depletion. I. Characterization of spectrin-free vesicles. J. Cell Biol. 1977, 73, 548–560. [Google Scholar] [CrossRef]
- D’Alessandro, A.; Kriebardis, A.G.; Rinalducci, S.; Antonelou, M.H.; Hansen, K.C.; Papassideri, I.S.; Zolla, L. An update on red blood cell storage lesions, as gleaned through biochemistry and omics technologies. Transfusion 2015, 55, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Greenwalt, T.J. The how and why of exocytic vesicles. Transfusion 2006, 46, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Kriebardis, A.G.; Antonelou, M.H.; Stamoulis, K.E.; Economou-Petersen, E.; Margaritis, L.H.; Papassideri, I.S. RBC-derived vesicles during storage: Ultrastructure, protein composition, oxidation, and signaling components. Transfusion 2008, 48, 1943–1953. [Google Scholar] [CrossRef] [PubMed]
- Willekens, F.L.A.; Roerdinkholder-Stoelwinder, B.; Groenen-Döpp, Y.A.M.; Bos, H.J.; Bosman, G.J.C.G.M.; van den Bos, A.G.; Verkleij, A.J.; Were, J.M. Hemoglobin loss from erythrocytes in vivo results from spleen-facilitated vesiculation. Blood 2003, 101, 747–751. [Google Scholar] [CrossRef] [PubMed]
- Willekens, F.L.; Were, J.M.; Groenen-Döpp, Y.A.; Roerdinkholder-Stoelwinder, B.; de Pauw, B.; Bosman, G.J. Erythrocyte vesiculation: A self-protective mechanism? Br. J. Haematol. 2008, 141, 549–556. [Google Scholar] [CrossRef]
- Pollet, H.; Conrard, L.; Cloos, A.S.; Tyteca, D. Plasma Membrane Lipid Domains as Platforms for Vesicle Biogenesis and Shedding? Biomolecules 2018, 8, 94. [Google Scholar] [CrossRef]
- Leal, J.K.F.; Adjobo-Hermans, M.J.W.; Bosman, G.J.C.G.M. Red Blood Cell Homeostasis: Mechanisms and Effects of Microvesicle Generation in Health and Disease. Front. Physiol. 2018, 9, 703. [Google Scholar] [CrossRef]
- Kostova, E.B.; Beuger, B.M.; Klei, T.R.; Halonen, P.; Lieftink, C.; Beijersbergen, R.L.; van den Berg, T.K.; van Bruggen, R. Identification of signalling cascades involved in red blood cell shrinkage and vesiculation. Biosci. Rep. 2015, 35, e00187. [Google Scholar] [CrossRef]
- Cloos, A.-S.; Ghodsi, M.; Stommen, A.; Vanderroost, J.; Dauguet, N.; Pollet, H.; D’Auria, L.; Mignolet, E.; Larondelle, Y.; Terrasi, R.; et al. Interplay Between Plasma Membrane Lipid Alteration, Oxidative Stress and Calcium-Based Mechanism for Extracellular Vesicle Biogenesis from Erythrocytes During Blood Storage. Front. Physiol. 2020, 11, 712. [Google Scholar] [CrossRef]
- Rifkind, J.M.; Nagababu, E. Hemoglobin redox reactions and red blood cell aging. Antioxid. Redox. Signal 2013, 18, 2274–2283. [Google Scholar] [CrossRef]
- Arashiki, N.; Kimata, N.; Manno, S.; Mohandas, N.; Takakuwa, Y. Membrane Peroxidation and Methemoglobin Formation Are Both Necessary for Band 3 Clustering: Mechanistic Insights into Human Erythrocyte Senescence. Biochemistry 2013, 52, 5760–5769. [Google Scholar] [CrossRef] [PubMed]
- Ferru, E.; Pantaleo, A.; Carta, F.; Mannu, F.; Khadjavi, A.; Gallo, V.; Ronzoni, L.; Graziadei, G.; Cappellini, M.D.; Turrini, F. Thalassemic erythrocytes release microparticles loaded with hemichromes by redox activation of p72Syk kinase. Haematologica 2014, 99, 570–578. [Google Scholar] [CrossRef] [PubMed]
- Jana, S.; Strader, M.B.; Meng, F.; Hicks, W.; Kassa, T.; Tarandovskiy, I.; de Paoli, S.; Simak, J.; Heaven, M.R.; Belcher, J.D.; et al. Hemoglobin oxidation-dependent reactions promote interactions with band 3 and oxidative changes in sickle cell-derived microparticles. JCI Insight 2018, 3, e120451. [Google Scholar] [CrossRef] [PubMed]
- Dinkla, S.; Brock, R.; Joosten, I.; Bosman, G.J. Gateway to understanding microparticles: Standardized isolation and identification of plasma membrane-derived vesicles. Nanomedicine (Lond). 2013, 8, 1657–1668. [Google Scholar] [CrossRef]
- Kanias, T.; Acker, J.P. Mechanism of hemoglobin-induced cellular injury in desiccated red blood cells. Free Radic. Biol. Med. 2010, 49, 539–547. [Google Scholar] [CrossRef]
- Mindukshev, I.; Gambaryan, S.; Kehrer, L.; Schuetz, C.; Kobsar, A.; Rukoyatkina, N.; Nikolaev, V.O.; Krivchenko, A.; Watson, S.P.; Walter, U.; et al. Low angle light scattering analysis: A novel quantitative method for functional characterization of human and murine platelet receptors. Clin. Chem. Lab. Med. 2012, 50, 1253–1262. [Google Scholar] [CrossRef]
- Mindukshev, I.; Kudryavtsev, I.; Serebriakova, M.; Trulioff, A.; Gambaryan, S.; Sudnitsyna, J.; Khmelevskoy, D.; Voitenko, N.; Avdonin, P.; Jenkins, R.; et al. Flow cytometry and light scattering technique in evaluation of nutraceuticals. In Nutraceuticals: Efficacy, Safety and Toxicity; Gupta, R.C., Ed.; Elsevier: Oxford, UK, 2016; pp. 319–332. [Google Scholar] [CrossRef]
- Mindukshev, I.V.; Sudnitsyna, J.S.; Skverchinskaya, E.A.; Andreyeva, A.Y.; Dobrylko, I.A.; Senchenkova, E.Y.; Krivchenko, A.I.; Gambaryan, S.P. Erythrocytes’ Reactions to Osmotic, Ammonium, and Oxidative Stress Are Inhibited under Hypoxia. Biochem (Moscow). Suppl. Ser. A Membr. Cell Biol. 2019, 13, 352–364. [Google Scholar] [CrossRef]
- Sudnitsyna, J.S.; Skverchinskaya, E.A.; Dobrylko, I.A.; Nikitina, E.R.; Krivchenko, A.I.; Gambaryan, S.P.; Mindukshev, I.V. Human erythrocyte ammonium transport is mediated by functional interaction of ammonium (RhAG) and anion (AE1) transporters. Biochem (Moscow). Suppl. Ser. A Membr. Cell Biol. 2016, 10, 301–310. [Google Scholar] [CrossRef]
- Darzynkiewicz, Z.; Juan, G.; Li, X.; Gorczyca, W.; Murakami, T.; Traganos, F. Cytometry in cell necrobiology: Analysis of apoptosis and accidental cell death (necrosis). Cytometry 1997, 27, 1–20. [Google Scholar] [CrossRef]
- Luchetti, C.G.; Solano, M.E.; Sander, V.; Arcos, M.L.; Gonzalez, C.D.; Di Girolamo, G.; Chiocchio, S.; Cremaschi, G.A.; Motta, A.B. Effects of dehydroepiandrosterone on ovarian cystogenesis and immune function. J. Reprod. Immunol. 2004, 64, 59–74. [Google Scholar] [CrossRef]
- Tzur, A.; Moore, J.K.; Jorgensen, P.; Shapiro, H.M.; Kirschner, M.W. Optimizing optical flow cytometry for cell volume-based sorting and analysis. PLoS ONE 2011, 6, e16053. [Google Scholar] [CrossRef] [PubMed]
- Bratosin, D.; Mitrofan, L.; Palii, C.; Estaquier, J.; Montreuil, J. Novel fluorescence assay using calcein-AM for the determination of human erythrocyte viability and aging. Cytometry A 2005, 66, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Kassa, T.; Jana, S.; Meng, F.; Alayash, A.I. Differential heme release from various hemoglobin redox states and the upregulation of cellular heme oxygenase-1. FEBS Open Bio 2016, 6, 876–884. [Google Scholar] [CrossRef]
- Chintagari, N.R.; Jana, S.; Alayas, A.I. Oxidized Ferric and Ferryl Forms of Hemoglobin Trigger Mitochondrial Dysfunction and Injury in Alveolar Type I Cells. Am. J. Respir. Cell Mol. Biol. 2016, 55, 288–298. [Google Scholar] [CrossRef]
- King, M.J.; Zanella, A. Hereditary red cell membrane disorders and laboratory diagnostic testing. Int. J. Lab. Hematol. 2013, 35, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Walski, T.; Chludzińska, L.; Komorowska, M.; Witkiewicz, W. Individual osmotic fragility distribution: A new parameter for determination of the osmotic properties of human red blood cells. Biomed. Res. Int. 2014, 2014, 162102. [Google Scholar] [CrossRef]
- Huisjes, R.; Bogdanova, A.; van Solinge, W.W.; Schiffelers, R.M.; Kaestner, L.; van Wijk, R. Squeezing for Life—Properties of Red Blood Cell Deformability. Front. Physiol. 2018, 9, 656. [Google Scholar] [CrossRef]
- Cueff, A.; Seear, R.; Dyrda, A.; Bouyer, G.; Egée, S.; Esposito, A.; Skepper, J.; Tiffert, T. Effects of elevated intracellular calcium on the osmotic fragility of human red blood cells. Cell Calcium. 2010, 47, 29–36. [Google Scholar] [CrossRef]
- Bogdanova, A.; Makhro, A.; Wang, J.; Lipp, P.; Kaestner, L. Calcium in red blood cells—A perilous balance. Int. J. Mol. Sci. 2013, 14, 9848–9872. [Google Scholar] [CrossRef]
- Kuck, L.; Peart, J.N.; Simmonds, M.J. Calcium dynamically alters erythrocyte mechanical response to shear [published online ahead of print, 2020]. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118802. [Google Scholar] [CrossRef]
- Kuck, L.; Peart, J.N.; Simmonds, M.J. Active modulation of human erythrocyte mechanics. Am. J. Physiol. Cell Physiol. 2020, 319, C250–C257. [Google Scholar] [CrossRef] [PubMed]
- Lang, P.A.; Kaiser, S.; Myssina, S.; Wieder, T.; Lang, F.; Huber, S.M. Role of Ca2+-activated K+ channels in human erythrocyte apoptosis. Am. J. Physiol Cell Physiol. 2003, 285, C1553–C1560. [Google Scholar] [CrossRef] [PubMed]
- Hertz, L.; Huisjes, R.; Llaudet-Planas, E.; Petkova-Kirova, P.; Makhro, A.; Danielczok, J.G.; Egee, S.; Del Mar Mañú-Pereira, M.; van Wijk, R.; Vives Corrons, J.L.; et al. Is Increased Intracellular Calcium in Red Blood Cells a Common Component in the Molecular Mechanism Causing Anemia? Front. Physiol. 2017, 8, 673. [Google Scholar] [CrossRef] [PubMed]
- Dodson, R.A.; Hinds, T.R.; Vincenzi, F.F. Effects of calcium and A23187 on deformability and volume of human red blood cells. Blood Cells 1987, 12, 555–564. [Google Scholar] [PubMed]
- Romero, P.J.; Hernández-Chinea, C. The Action of Red Cell Calcium Ions on Human Erythrophagocytosis in Vitro. Front. Physiol. 2017, 8, 1008. [Google Scholar] [CrossRef]
- Hierso, R.; Waltz, X.; Mora, P.; Romana, M.; Lemonne, N.; Connes, P.; Hardy-Dessources, M.-D. Effects of oxidative stress on red blood cell rheology in sickle cell patients. Br. J. Haematol. 2014, 166, 601–606. [Google Scholar] [CrossRef]
- Raducka-Jaszul, O.; Bogusławska, D.M.; Jędruchniewicz, N.; Sikorski, A.F. Role of Extrinsic Apoptotic Signaling Pathway during Definitive Erythropoiesis in Normal Patients and in Patients with β-Thalassemia. Int. J. Mol. Sci. 2020, 21, 3325. [Google Scholar] [CrossRef]
- Shimo, H.; Arjunan, S.N.; Machiyama, H.; Nishino, T.; Suematsu, M.; Fujita, H.; Tomita, M.; Takahashi, K. Particle Simulation of Oxidation Induced Band 3 Clustering in Human Erythrocytes. PLoS Comput. Biol. 2015, 11, e1004210. [Google Scholar] [CrossRef]
- Gallagher, P.G. Red blood cell membrane disorders. In Hematology: Basic Principles and Practice; Hoffman, R., Benz, E.J., Silberstein, L.E., Heslop, H.E., Weitz, J.I., Anastasi, J., Salama, M.E., Abutalib, S., Eds.; Elsevier: Oxford, UK, 2018; pp. 626–647. [Google Scholar] [CrossRef]
- Bevers, E.M.; Wiedmer, T.; Comfurius, P.; Shattil, S.J.; Weiss, H.J.; Zwaal, R.F.; Sims, P.J. Defective Ca(2+)-induced microvesiculation and deficient expression of procoagulant activity in erythrocytes from a patient with a bleeding disorder: A study of the red blood cells of Scott syndrome. Blood 1992, 79, 380–388. [Google Scholar] [CrossRef]
- Hafiane, A.; Daskalopoulou, S.S. Extracellular vesicles characteristics and emerging roles in atherosclerotic cardiovascular disease. Metabolism 2018, 85, 213–222. [Google Scholar] [CrossRef]
- Westerman, M.; Porter, J.B. Red blood cell-derived microparticles: An overview. Blood Cells Mol. Dis. 2016, 59, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Bosman, G.J.; Lasonder, E.; Groenen-Döpp, Y.A.; Willekens, F.L.; Were, J.M. The proteome of erythrocyte-derived microparticles from plasma: New clues for erythrocyte aging and vesiculation. J. Proteom. 2012, 76, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Li, K.Y.; Zheng, L.; Wang, Q.; Hu, Y.W. Characteristics of erythrocyte-derived microvesicles and its relation with atherosclerosis. Atherosclerosis 2016, 255, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Noubouossie, D.F.; Henderson, M.W.; Mooberry, M.; Ilich, A.; Ellsworth, P.; Piegore, M.; Skinner, S.C.; Pawlinski, R.; Welsby, I.; Renné, T.; et al. Red blood cell microvesicles activate the contact system, leading to factor IX activation via 2 independent pathways. Blood 2020, 135, 755–765. [Google Scholar] [CrossRef]
- Danesh, A.; Inglis, H.C.; Jackman, R.P.; Wu, S.; Deng, X.; Muench, M.O.; Heitman, J.W.; Norris, P.J. Exosomes from red blood cell units bind to monocytes and induce proinflammatory cytokines, boosting T-cell responses in vitro. Blood 2014, 123, 687–696. [Google Scholar] [CrossRef]
- Tissot, J.D.; Rubin, O.; Canellini, G. Analysis and clinical relevance of microparticles from red blood cells. Curr. Opin. Hematol. 2010, 17, 571–577. [Google Scholar] [CrossRef]
- Lang, E.; Qadri, S.M.; Lang, F. Killing me softly-suicidal erythrocyte death. Int. J. Biochem. Cell Biol. 2012, 44, 1236–1243. [Google Scholar] [CrossRef]
- Föller, M.; Lang, F. Ion Transport in Eryptosis, the Suicidal Death of Erythrocytes. Front. Cell Dev. Biol. 2020, 8, 597. [Google Scholar] [CrossRef]
- Carelli-Alinovi, C.; Misiti, F. Erythrocytes as Potential Link between Diabetes and Alzheimer’s Disease. Front. Aging Neurosci. 2017, 9, 276. [Google Scholar] [CrossRef]
- Turpin, C.; Catan, A.; Guerin-Dubourg, A.; Debussche, X.; Bravo, S.B.; Álvarez, E.; van den Elsen, J.; Meilhac, O.; Rondeau, P.; Bourdon, E. Enhanced oxidative stress and damage in glycated erythrocytes. PLoS ONE 2020, 15, e0235335. [Google Scholar] [CrossRef]
- Föller, M.; Huber, S.M.; Lang, F. Erythrocyte programmed cell death. IUBMB Life 2008, 60, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Bateman, R.M.; Sharpe, M.D.; Singer, M.; Ellis, C.G. The Effect of Sepsis on the Erythrocyte. Int. J. Mol. Sci. 2017, 18, 1932. [Google Scholar] [CrossRef] [PubMed]
- Lang, F.; Bissinger, R.; Abed, M.; Artunc, F. Eryptosis—The neglected cause of anemia in end stage renal disease. Kidney Blood Press. Res. 2017, 42, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Bogdanova, A.; Kaestner, L.; Simionato, G.; Wickrema, A.; Makhro, A. Heterogeneity of Red Blood Cells: Causes and Consequences. Front. Physiol. 2020, 11, 392. [Google Scholar] [CrossRef]
- Petrou, T.; Olsen, H.L.; Thrasivoulou, C.; Masters, J.R.; Ashmore, J.F.; Ahmed, A. Intracellular Calcium Mobilization in Response to Ion Channel Regulators via a Calcium-Induced Calcium Release Mechanism. J. Pharmacol. Exp. Ther. 2017, 360, 378–387. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hb Species | Control | t-BOOH | A23187 | SNC | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.5 mM | 1 mM | 2 mM | 1 µM | 500 µM | ||||||||
| 1 h | 3 h | 1 h | 3 h | 1 h | 3 h | 1 h | 3 h | 1 h | 3 h | 1 h | 3 h | |
| % OxyHb | 98.0 ± 2.0 | 95.6 ± 3.0 | 66.3 ± 4.2 * | 45.3 ± 3.9 * | 30.7 ± 9.0 ** | 27.0 ± 13.6 ** | 20.9 ± 16.4 ** | 20.7 ± 15.1 ** | 97.4 ± 3.1 | 96.6 ± 3.0 | 47.2 ± 5.9 ** | 33.1 ± 3.4 ** |
| % MetHb | 1.8 ± 0.1 | 4.0 ± 2.3 | 30.6 ± 5.1 * | 49.1 ± 6.6 * | 50.5 ± 8.1 ** | 47.0 ± 11.8 ** | 58.8 ± 9.4 ** | 46.1 ± 6.6 ** | 2.3 ± 0.5 | 3.4 ± 0.6 | 50.8 ± 4.1 ** | 63.4 ± 4.6 ** |
| % HbChr | 0.2 ± 0.6 | 0.4 ± 0.2 | 4.9 ± 3.6 | 5.4 ± 2.1 | 18.8 ± 5.4 * | 25.9 ± 8.1 ** | 20.3 ± 12.8 * | 33.2 ± 9.1 ** | 0.3 ± 0.2 | 0.2 ± 0.1 | 2.5 ± 1.4 | 3.6 ± 2.5 |
| OFT Characteristics | Control | t-BOOH | A23187 | SNC | ||
|---|---|---|---|---|---|---|
| 0.5 mM | 1 mM | 2 mM | 1 µM | 500 µM | ||
| H50, mOsm/kg H2O | 122 ± 9 | 130 ± 7 | 117 ± 8 | 83 ± 6 ** | 68 ± 7 ** | 128 ± 6 |
| W, mOsm/kg H2O | 44 ± 7 | 52 ± 8 | 60 ± 9 * | 107 ± 10 ** | 41 ± 6 | 47 ± 7 |
| MCV300, fL | 84 ± 2 | 88 ± 6 | 87 ± 7 | 81 ± 6 | 73 ± 2 ** | 86 ± 5 |
| MCV120, fL | 124 ± 9 | 131 ± 8 | 115 ± 8 | 88 ± 6 * | 83 ± 2 ** | 127 ± 4 |
| AST Characteristics | Control | t-BOOH | A23187 | SNC | |||
|---|---|---|---|---|---|---|---|
| 0.5 mM | 1 mM | 1.5 mM | 2 mM | 1 µM | 500 µM | ||
| Vhem | 1.00 ± 0.02 | 1.01 ± 0.02 | 0.71 ± 0.21 | 0.57 ± 0.08 * | 0.32 ± 0.09 * | 0.06 ± 0.02 ** | 1.04 ± 0.02 |
| %Hem | 96.5 ± 0.2 | 92.1 ± 2.6 | 80.8 ± 11.4 | 69.3 ± 9.6 * | 47.8 ± 13.2 ** | 40.0 ± 11.3 ** | 95.6 ± 1.0 |
| MCV300 | 84.9 ± 0.9 | 86.7 ± 2.3 | 87.5 ± 5.9 | 87.8 ± 4.3 | 88.6 ± 10.4 | 70.5 ± 3.7 * | 86.87 ± 1.9 |
| MCVhem | 142.3 ± 3.2 | 143.8 ± 4.8 | 124.5 ± 9.8 | 116.0 ± 1.7 * | 105.1 ± 1.9 ** | 96.3 ± 6.5 ** | 141.0 ± 2.8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sudnitsyna, J.; Skverchinskaya, E.; Dobrylko, I.; Nikitina, E.; Gambaryan, S.; Mindukshev, I. Microvesicle Formation Induced by Oxidative Stress in Human Erythrocytes. Antioxidants 2020, 9, 929. https://doi.org/10.3390/antiox9100929
Sudnitsyna J, Skverchinskaya E, Dobrylko I, Nikitina E, Gambaryan S, Mindukshev I. Microvesicle Formation Induced by Oxidative Stress in Human Erythrocytes. Antioxidants. 2020; 9(10):929. https://doi.org/10.3390/antiox9100929
Chicago/Turabian StyleSudnitsyna, Julia, Elisaveta Skverchinskaya, Irina Dobrylko, Elena Nikitina, Stepan Gambaryan, and Igor Mindukshev. 2020. "Microvesicle Formation Induced by Oxidative Stress in Human Erythrocytes" Antioxidants 9, no. 10: 929. https://doi.org/10.3390/antiox9100929
APA StyleSudnitsyna, J., Skverchinskaya, E., Dobrylko, I., Nikitina, E., Gambaryan, S., & Mindukshev, I. (2020). Microvesicle Formation Induced by Oxidative Stress in Human Erythrocytes. Antioxidants, 9(10), 929. https://doi.org/10.3390/antiox9100929