Effects of Selenium- and Zinc-Enriched Lactobacillus plantarum SeZi on Antioxidant Capacities and Gut Microbiome in an ICR Mouse Model



Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation of Probiotic Strains from Human Feces
2.2. Screening of SeZn-Tolerant LAB Isolates from Human Feces
2.3. Assessing Bioaccumulation of Selenium and Zinc in LAB Strains During the Cultivation
2.4. Gene Analysis of Se/Zn Uptake and Resistance in L. plantarum SeZi
2.5. Effect of SeZn-Enriched L. plantarum SeZi in an ICR Mouse Model
2.5.1. Preparation of SeZn-Enriched L. plantarum SeZi for Mouse Study
2.5.2. Animals and Diets
2.5.3. Blood Analysis
2.5.4. Bacterial Community Analysis by 16S rRNA Gene Amplicon Sequencing
2.6. Statistic Analysis
3. Results
3.1. Screening and Selection of SeZn-Tolerant LAB Strains
3.2. Increased Concentrations of Selenium and Zinc in Blood after L. plantarum SeZi Administration
3.3. Increased Antioxidant Activities in Mice after L. plantarum SeZi Administration
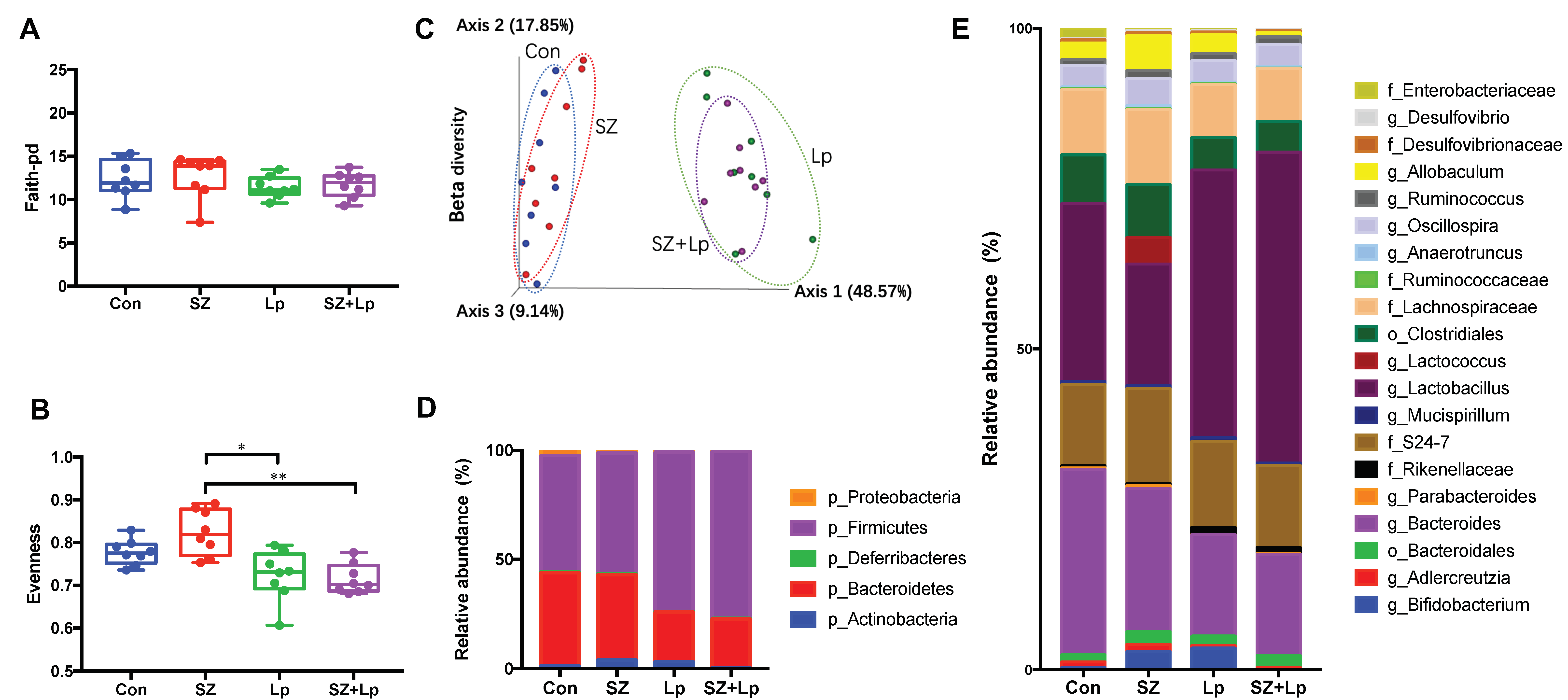
3.4. Changes in the Gut Microbiota after L. plantarum SeZi Administration
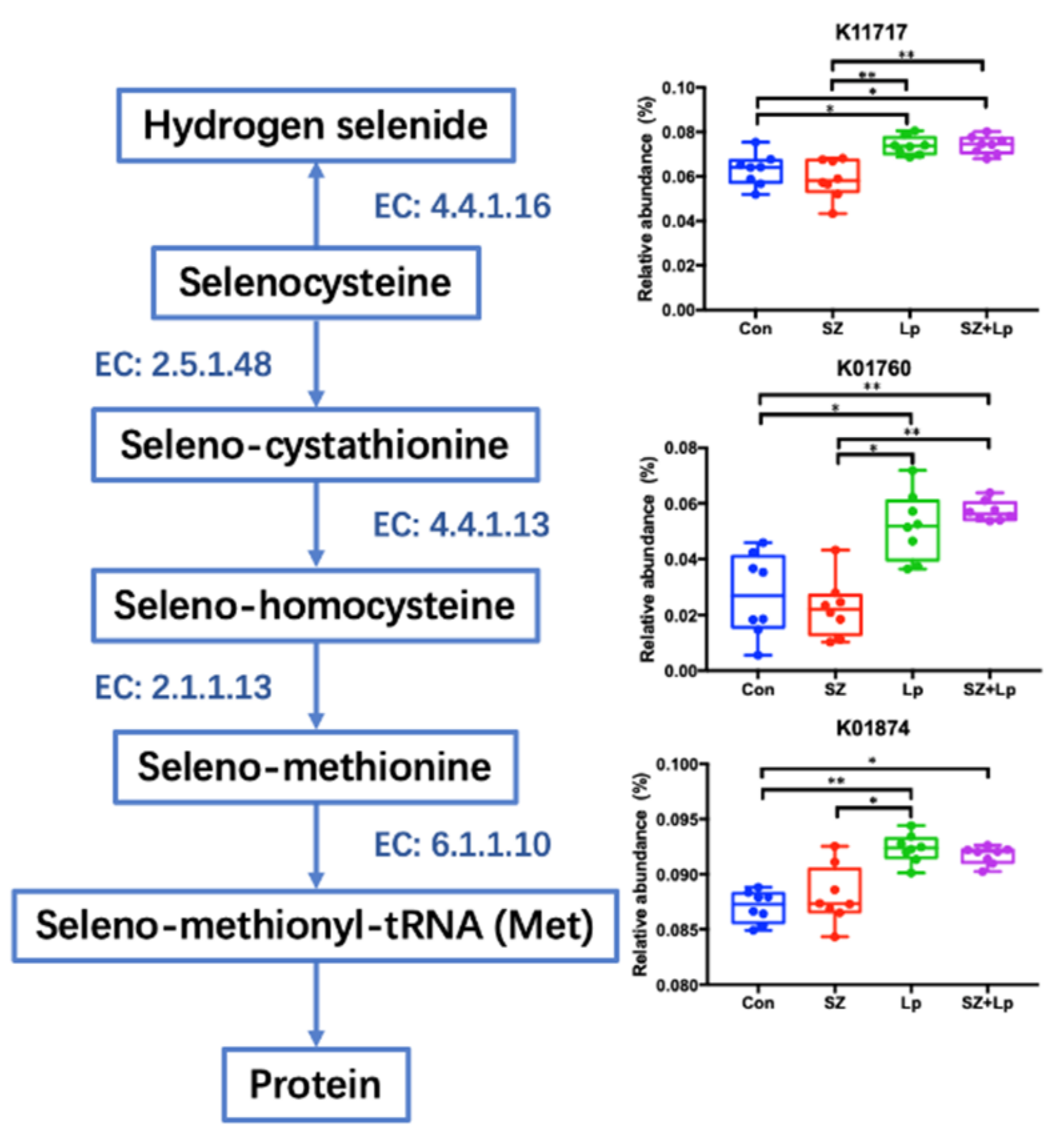
3.5. Microbial Function Analysis Related to Selenocompounds Metabolism
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mrvčić, J.; Stanzer, D.; Šolić, E.; Stehlik-Tomas, V. Interaction of lactic acid bacteria with metal ions: Opportunities for improving food safety and quality. World J. Microb. Biot. 2012, 28, 2771–2782. [Google Scholar] [CrossRef]
- Rayman, M.P. Selenium and adverse health conditions of human pregnancy. In Selenium; Springer: New York, NY, USA, 2011; pp. 531–544. [Google Scholar]
- Flores-Mateo, G.; Navas-Acien, A.; Pastor-Barriuso, R.; Guallar, E. Selenium and coronary heart disease: A meta-analysis. Am. J. Clin. Nutr. 2006, 84, 762–773. [Google Scholar] [CrossRef] [PubMed]
- Brenneisen, P.; Steinbrenner, H.; Sies, H. Selenium, oxidative stress, and health aspects. Mol. Asp. Med. 2005, 26, 256–267. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, L.R.; Karunasinghe, N.; Zhu, S.; Wang, A.H. Selenium and its’ role in the maintenance of genomic stability. Mutat. Res. Fundam. Mol. Mech. Mutagenesis 2012, 733, 100–110. [Google Scholar] [CrossRef]
- Rayman, M.P. Selenium and human health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- Arthur, J.R.; Nicol, F.; Beckett, G.J. The role of selenium in thyroid hormone metabolism and effects of selenium deficiency on thyroid hormone and iodine metabolism. Biol. Trace Elem. Res. 1992, 33, 37–42. [Google Scholar] [CrossRef]
- Burke, M.P.; Opeskin, K. Fulminant heart failure due to selenium deficiency cardiomyopathy (Keshan disease). Med. Sci. Law 2002, 42, 10–13. [Google Scholar] [CrossRef]
- Maksimović, Z.J. Selenium deficiency and Balkan endemic nephropathy. Kidney Int. Suppl. 1991, 34, S12–S14. [Google Scholar] [PubMed]
- Gropper, S.S.; Smith, J.L. Advanced Nutrition and Human Metabolism. Wadsworth Cengage Learning: Belmot, CA, USA, 2012. [Google Scholar]
- Chimienti, F. Zinc, pancreatic islet cell function and diabetes: New insights into an old story. Nutr. Res. Rev. 2013, 26, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Cruz, K.J.C.; de Oliveira, A.R.S.; Morais, J.B.S.; Severo, J.S.; Mendes, P.M.V.; de Sousa Melo, S.R.; de Sousa, G.S.; do Nascimento Marreiro, D. Zinc and insulin resistance: Biochemical and molecular aspects. Biol. Trace Elem. Res. 2018, 186, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.R.; Attia, J.; Ali, L.; McEvoy, M.; Selim, S.; Sibbritt, D.; Akhter, A.; Akter, S.; Peel, R.; Faruque, O. Zinc supplementation for improving glucose handling in pre-diabetes: A double blind randomized placebo controlled pilot study. Diabetes Res. Clin. Pract. 2016, 115, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Afolabi, O.B.; Balogun, B.D.; Oloyede, O.I.; Akinyemi, A.J. Zinc and neurodegenerative disorders. In Handbook of Research on Critical Examinations of Neurodegenerative Disorders; IGI Global: Harrisburg, PA, USA, 2019; pp. 176–193. [Google Scholar]
- Gomolak, J.R. Zinc accumulation in the midbrain following methamphetamine exposure as a potential biomarker for neurodegeneration. In Proceedings of the 25th UNG Annual Research Conference, Dahlonega, GA, USA, 11 November 2016. [Google Scholar]
- Kieliszek, M.; Błażejak, S. Selenium: Significance, and outlook for supplementation. Nutrition 2013, 29, 713–718. [Google Scholar] [CrossRef] [PubMed]
- Ledgham, F.; Quest, B.; Vallaeys, T.; Mergeay, M.; Covès, J. A probable link between the DedA protein and resistance to selenite. Res. Microbiol. 2005, 156, 367–374. [Google Scholar] [CrossRef]
- Ayangbenro, A.S.; Babalola, O.O. A new strategy for heavy metal polluted environments: A review of microbial biosorbents. Int. J. Environ. Res. Pub. He. 2017, 14, 94. [Google Scholar] [CrossRef]
- Blackwell, K.; Singleton, I.; Tobin, J.M. Metal cation uptake by yeast: A review. Appl. Microbiol. Biot. 1995, 43, 579–584. [Google Scholar] [CrossRef]
- Andreoni, V.; Luischi, M.M.; Cavalca, L.; Erba, D.; Ciappellano, S. Selenite tolerance and accumulation in the Lactobacillus species. Ann. Microbiol. 2000, 50, 77–88. [Google Scholar]
- Saini, K.; Tomar, S.K. In vitro evaluation of probiotic potential of Lactobacillus cultures of human origin capable of selenium bioaccumulation. LWT 2017, 84, 497–504. [Google Scholar] [CrossRef]
- Shu, G.; Mei, S.; Chen, L.; Zhang, B.; Guo, M.; Cui, X.; Chen, H. Screening, identification, and application of selenium--enriched Lactobacillus in goat milk powder and tablet. J. Food Process. Pres. 2020, e14470. [Google Scholar] [CrossRef]
- Xia, S.K.; Chen, L.; Liang, J.Q. Enriched selenium and its effects on growth and biochemical composition in Lactobacillus bulgaricus. J. Agric. Food. Chem. 2007, 55, 2413–2417. [Google Scholar] [CrossRef]
- Diowksz, A.; Ambroziak, W.; Wlodarczyk, M. Investigation of the ability of selenium accumulation by lactic acid bacteria of Lactobacillus species and yeast Saccharomyces cerevisiae. Pol. J. Food Nutr. Sci. 1999, 1, 17–22. [Google Scholar]
- Mudroňová, D.; Gancarčíková, S.; Nemcová, R. Influence of Zinc Sulphate on the Probiotic Properties of Lactobacillus plantarum CCM 7102. Folia Veterinaria 2019, 63, 45–54. [Google Scholar] [CrossRef]
- Jin, H.; Jeong, Y.; Yoo, S.H.; Johnston, T.V.; Ku, S.; Ji, G.E. Isolation and characterization of high exopolysaccharide-producing Weissella confusa VP30 from young children’s feces. Microb. Cell Fact. 2019, 18, 110. [Google Scholar] [CrossRef]
- Chaney, A.L.; Marbach, E.P. Modified reagents for determination of urea and ammonia. Clin. Chem. 1962, 8, 130–132. [Google Scholar] [CrossRef]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613. [Google Scholar] [CrossRef]
- Schattner, P.; Brooks, A.N.; Lowe, T.M. The tRNAscan-SE, snoscan and snoGPS web servers for the detection of tRNAs and snoRNAs. Nucleic Acids Res. 2005, 33 (Suppl. 2), W686–W689. [Google Scholar] [CrossRef]
- Nawrocki, E.P.; Eddy, S.R. Computational identification of functional RNA homologs in metagenomic data. RNA Biol. 2013, 10, 1170–1179. [Google Scholar] [CrossRef]
- Hyatt, D.; Chen, G.L.; LoCascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef]
- Powell, S.; Forslund, K.; Szklarczyk, D.; Trachana, K.; Roth, A.; Huerta-Cepas, J.; Gabaldon, T.; Rattei, T.; Creevey, C.; Kuhn, M. eggNOG v4. 0: Nested orthology inference across 3686 organisms. Nucleic Acids Res. 2014, 42, D231–D239. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Consortium, U. UniProt: A hub for protein information. Nucleic Acids Res. 2015, 43, D204–D212. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. Data, information, knowledge and principle: Back to metabolism in KEGG. Nucleic Acids Res. 2014, 42, D199–D205. [Google Scholar] [CrossRef]
- Overbeek, R.; Begley, T.; Butler, R.M.; Choudhuri, J.V.; Chuang, H.Y.; Cohoon, M.; de Crécy-Lagard, V.; Diaz, N.; Disz, T.; Edwards, R. The subsystems approach to genome annotation and its use in the project to annotate 1000 genomes. Nucleic Acids Res. 2005, 33, 5691–5702. [Google Scholar] [CrossRef]
- Kang, S.; You, H.J.; Lee, Y.G.; Jeong, Y.; Johnston, T.V.; Baek, N.I.; Ku, S.; Ji, G.E. Production, structural characterization, and In Vitro assessment of the prebiotic potential of butyl-fructooligosaccharides. Int. J. Mol. Sci. 2020, 21, 445. [Google Scholar] [CrossRef]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 1–5. [Google Scholar] [CrossRef]
- Younus, H. Therapeutic potentials of superoxide dismutase. Int. J. Health Sci. 2018, 12, 88. [Google Scholar]
- Blankenberg, S.; Rupprecht, H.J.; Bickel, C.; Torzewski, M.; Hafner, G.; Tiret, L.; Smieja, M.; Cambien, F.; Meyer, J.; Lackner, K.J. Glutathione peroxidase 1 activity and cardiovascular events in patients with coronary artery disease. N. Engl. J. Med. 2003, 349, 1605–1613. [Google Scholar] [CrossRef]
- Khoubnasabjafari, M.; Ansarin, K.; Jouyban, A. Reliability of malondialdehyde as a biomarker of oxidative stress in psychological disorders. BioImpacts 2015, 5, 123. [Google Scholar] [PubMed]
- Yang, H.; Chang, H. Antioxidant and antitumor activities of selenium and zinc-enriched oyster mushroom in mice. Biol. Trace Elem. Res. 2012, 150, 236–241. [Google Scholar]
- Xu, C.; Guo, Y.; Qiao, L.; Ma, L.; Cheng, Y.; Roman, A. Biogenic synthesis of novel functionalized selenium nanoparticles by Lactobacillus casei ATCC 393 and its protective effects on intestinal barrier dysfunction caused by enterotoxigenic Escherichia coli K88. Front. Microbiol. 2018, 9, 1129. [Google Scholar] [CrossRef] [PubMed]
- Nagy, G.; Pinczes, G.; Pinter, G.; Pocsi, I.; Prokisch, J.; Banfalvi, G. In situ electron microscopy of lactomicroselenium particles in probiotic bacteria. Int. J. Mol. Sci. 2016, 17, 1047. [Google Scholar] [CrossRef]
- Kurek, E.; Ruszczynska, A.; Wojciechowski, M.; Luciuk, A.; Michalska-Kacymirow, M.; Motyl, I.; Bulska, E. Bio-transformation of selenium in Se-enriched bacterial strains of Lactobacillus casei. Roczniki Państwowego Zakładu Higieny 2016, 67, 3. [Google Scholar]
- Yazdi, M.H.; Mahdavi, M.; Setayesh, N.; Esfandyar, M.; Shahverdi, A.R. Selenium n6noparticle-enriched Lactobacillus brevis causes more efficient immune responses in vivo and reduces the liver metastasis in metastatic form of mouse breast cancer. DARU 2013, 21, 33. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Dou, X.; Yan, S.; Zhang, B.; Xu, C. Biogenic selenium nanoparticles synthesized by Lactobacillus casei ATCC 393 alleviate diquat-induced intestinal barrier dysfunction in C57BL/6 mice through their antioxidant activity. Food Funct. 2020, 11, 3020–3031. [Google Scholar] [CrossRef] [PubMed]
- Turner, R.J.; Weiner, J.H.; Taylor, D.E. Selenium metabolism in Escherichia coli. Biometals 1998, 11, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, A.; Zanoni, S.; de Lucia, M.; Amaretti, A.; Raimondi, S.; Rossi, M. Zinc uptake by lactic acid bacteria. ISRN Biotechnol. 2013, 2013. [Google Scholar] [CrossRef]
- Góral, M.; Pankiewicz, U.; Sujka, M.; Kowalski, R. Bioaccumulation of zinc ions in Lactobacillus rhamnosus B 442 cells under treatment of the culture with pulsed electric field. Eur. Food Res. Technol. 2019, 245, 817–824. [Google Scholar] [CrossRef]
- Lule, V.K.; Tomar, S.K.; Chawla, P.; Pophaly, S.; Kapila, S.; Arora, S. Bioavailability assessment of zinc enriched lactobacillus biomass in a human colon carcinoma cell line (Caco-2). Food Chem. 2020, 309, 125583. [Google Scholar] [CrossRef]
- Malyar, R.M.; Li, H.; Liu, D.; Abdulrahim, Y.; Farid, R.A.; Gan, F.; Ali, W.; Enayatullah, H.; Banuree, S.A.H.; Huang, K. Selenium/Zinc-Enriched probiotics improve serum enzyme activity, antioxidant ability, inflammatory factors and related gene expression of Wistar rats inflated under heat stress. Life Sci. 2020, 248, 117464. [Google Scholar] [CrossRef]
- Benstoem, C.; Goetzenich, A.; Kraemer, S.; Borosch, S.; Manzanares, W.; Hardy, G.; Stoppe, C. Selenium and its supplementation in cardiovascular disease—what do we know? Nutrients 2015, 7, 3094–3118. [Google Scholar] [CrossRef]
- Fan, Y.; Zhang, J. Dietary modulation of intestinal microbiota: Future opportunities in experimental autoimmune encephalomyelitis and multiple sclerosis. Front. Microbiol. 2019, 10, 740. [Google Scholar] [CrossRef]
- Vogt, N.M.; Kerby, R.L.; Dill-McFarland, K.A.; Harding, S.J.; Merluzzi, A.P.; Johnson, S.C.; Carlsson, C.M.; Asthana, S.; Zetterberg, H.; Blennow, K. Gut microbiome alterations in Alzheimer’s disease. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Maruo, T.; Sakamoto, M.; Ito, C.; Toda, T.; Benno, Y. Adlercreutzia equolifaciens gen. nov., sp. nov., an equol-producing bacterium isolated from human faeces, and emended description of the genus Eggerthella. Int. J. Syst. Evol. Micr. 2008, 58, 1221–1227. [Google Scholar] [CrossRef]
- Martínez, F.G.; Moreno-Martin, G.; Pescuma, M.; Madrid-Albarrán, Y.; Mozzi, F. Biotransformation of selenium by lactic acid bacteria: Formation of seleno-nanoparticles and seleno-amino acids. Front. Bioeng. Biotechnol. 2020, 8, 506. [Google Scholar] [CrossRef]
- Llull, D.; Poquet, I. New expression system tightly controlled by zinc availability in Lactococcus lactis. Appl. Environ. Microbiol. 2004, 70, 5398–5406. [Google Scholar] [CrossRef]
- Lee, S.M.; Han, H.W.; Yim, S.Y. Beneficial effects of soymilk and fiber on high cholesterol diet-induced alteration of gut microbiota and inflammatory gene expression in rats. Food Funct. 2015, 6, 492–500. [Google Scholar] [CrossRef]
- Palm, N.W.; de Zoete, M.R.; Cullen, T.W.; Barry, N.A.; Stefanowski, J.; Hao, L.; Degnan, P.H.; Hu, J.; Peter, I.; Zhang, W. Immunoglobulin A coating identifies colitogenic bacteria in inflammatory bowel disease. Cell 2014, 158, 1000–1010. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Biomass (g/L) | Bioconversion Rate (%) | |
|---|---|---|---|
| Selenium | Zinc | ||
| Lactobacillus plantarum SeZi | 2.82 ± 0.28 a | 19.47 | 0.35 |
| Lactobacillus pentosus SeZi | 2.33 ± 0.11 ab | 8.93 | 0.36 |
| Lactobacillus fermentum SeZi | 1.94 ± 0.34 bc | 18.04 | 0.33 |
| Lactobacillus rhamnosus SeZi | 1.78 ± 0.34 c | 6.90 | 0.20 |
| Coding Region a | Length (aa) | Product | Function |
|---|---|---|---|
| 264974–265621 (−) | 648 | DedA protein | Detoxification and uptake of selenate |
| 193218–193874 (−) | 657 | DedA protein | Detoxification and uptake of selenate |
| 19823–20716 | 894 | Sulfate permease-CysA | Detoxification and uptake of selenate |
| Coding Region a | Length (aa) | Product | Function |
|---|---|---|---|
| 237659–238102 | 444 | ZUR | Zinc uptake regulation |
| 151–966 | 816 | Multidrug efflux transporter 1 regulator | Zinc resistance |
| 31131–31565 | 435 | Uncharacterized HTH-type transcriptional regulator | Zinc resistance |
| 174455–174916 | 462 | MerR family | Zinc resistance |
| 22703–23146 | 444 | MerR family | Zinc resistance |
| 115660–116040 | 381 | MerR family | Zinc resistance |
| 27565–27957 (−) | 393 | MerR family | Zinc resistance |
| 4022–4399 | 378 | MerR family | Zinc resistance |
| 227–619 (−) | 393 | MerR family | Zinc resistance |
| 342254–342718 | 465 | Hypothetical protein | Zinc resistance |
| 54502–54756 (−) | 255 | Hypothetical protein | Zinc resistance |
| 54351–54800 | 450 | Hypothetical protein | Zinc resistance |
| DedA | CysA | ZUR | MerR Family |
|---|---|---|---|
| L. acidophilus La-14 | - | L.rhamnosus LOCK908 | L. acidophilus La-14 |
| L. gasseri ATCC 33323 | L.rhamnosus LOCK900 | L. curvatus JCM 1096 | |
| L. dekbrueckii subsp. delbrueckii | L.rhamnosus LOCK919 | L. gasseri ATCC 33323 | |
| L. salivarius str. Ren | L. ruminis ATCC 25644 | ||
| L. buchneri subsp. silagei CD034 | L. buchneri subsp. silagei CD034 | ||
| L. rhamnosus GG | |||
| L. paracasei subsp. paracasei 8700:2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, S.; Li, R.; Jin, H.; You, H.J.; Ji, G.E. Effects of Selenium- and Zinc-Enriched Lactobacillus plantarum SeZi on Antioxidant Capacities and Gut Microbiome in an ICR Mouse Model. Antioxidants 2020, 9, 1028. https://doi.org/10.3390/antiox9101028
Kang S, Li R, Jin H, You HJ, Ji GE. Effects of Selenium- and Zinc-Enriched Lactobacillus plantarum SeZi on Antioxidant Capacities and Gut Microbiome in an ICR Mouse Model. Antioxidants. 2020; 9(10):1028. https://doi.org/10.3390/antiox9101028
Chicago/Turabian StyleKang, Sini, Rui Li, Hui Jin, Hyun Ju You, and Geun Eog Ji. 2020. "Effects of Selenium- and Zinc-Enriched Lactobacillus plantarum SeZi on Antioxidant Capacities and Gut Microbiome in an ICR Mouse Model" Antioxidants 9, no. 10: 1028. https://doi.org/10.3390/antiox9101028
APA StyleKang, S., Li, R., Jin, H., You, H. J., & Ji, G. E. (2020). Effects of Selenium- and Zinc-Enriched Lactobacillus plantarum SeZi on Antioxidant Capacities and Gut Microbiome in an ICR Mouse Model. Antioxidants, 9(10), 1028. https://doi.org/10.3390/antiox9101028