Natural Antioxidants Beneficial Effects on Anion Exchange through Band 3 Protein in Human Erythrocytes




Abstract
1. Introduction
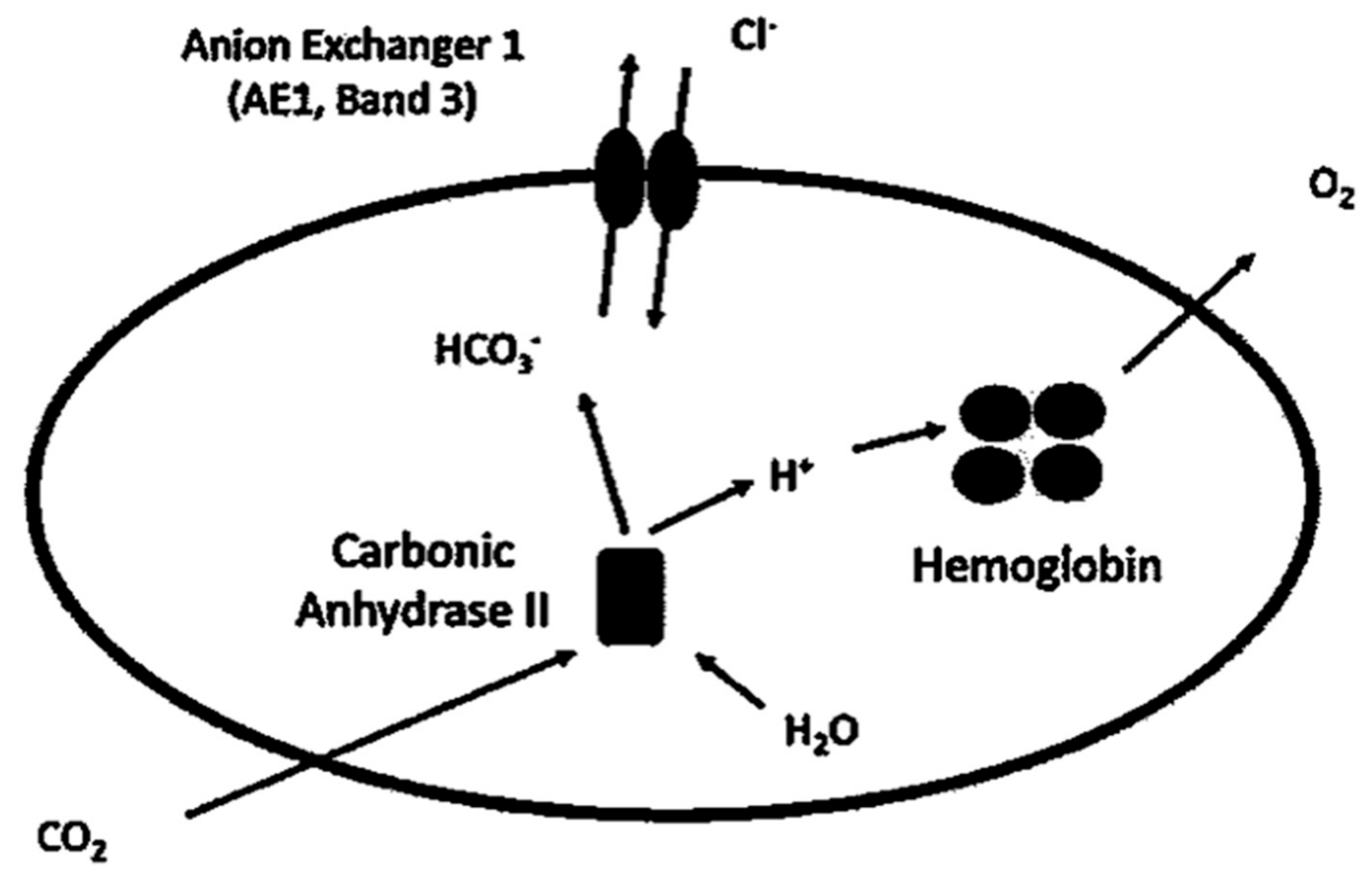
2. Role of Band 3 Protein in Erythrocytes
2.1. Role in CO2 Transport
2.2. Role in Erythrocyte Rigidity
3. Anion Exchange through Band 3 Protein in Different Oxidative Experimental Conditions and the Role of Antioxidants
3.1. Curcumin
3.2. Hydrogen Peroxide-Induced Oxidative Conditions and the Beneficial Effect of Antioxidants
3.3. Magnesium
3.4. Melatonin
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Halliwell, B.; Rafter, J.; Jenner, A. Health promotion by flavonoids, tocopherols, tocotrienols, and other phenols: Direct or indirect effects? Antioxidant or not? Am. J. Clin. Nutr. 2005, 81, 268S–276S. [Google Scholar] [CrossRef]
- Neha, K.; Haider, M.R.; Pathak, A.; Yar, M.S. Medicinal prospects of antioxidants: A review. Eur. J. Med. Chem. 2019, 178, 687–704. [Google Scholar] [CrossRef]
- Zujko, M.E.; Witkowska, A.M.; Waskiewicz, A.; Sygnowska, E. Estimation of dietary intake and patterns of polyphenol consumption in Polish adult population. Adv. Med. Sci. 2012, 57, 375–384. [Google Scholar] [CrossRef]
- Uttara, B.; Singh, A.V.; Zamboni, P.; Mahajan, R.T. Oxidative stress and neurodegenerative diseases: A review of upstream and downstream antioxidant therapeutic options. Curr. Neuropharmacol. 2009, 7, 65–74. [Google Scholar] [CrossRef]
- Khansari, N.; Shakiba, Y.; Mahmoudi, M. Chronic inflammation and oxidative stress as a major cause of age-related diseases and cancer. Recent Pat. Inflamm. Allergy Drug Discov. 2009, 3, 73–80. [Google Scholar] [CrossRef]
- Cervantes Gracia, K.; Llanas-Cornejo, D.; Husi, H. CVD and Oxidative Stress. J. Clin. Med. 2017, 6. [Google Scholar] [CrossRef]
- Ferru, E.; Giger, K.; Pantaleo, A.; Campanella, E.; Grey, J.; Ritchie, K.; Vono, R.; Turrini, F.; Low, P.S. Regulation of membrane-cytoskeletal interactions by tyrosine phosphorylation of erythrocyte band 3. Blood 2011, 117, 5998–6006. [Google Scholar] [CrossRef] [PubMed]
- Morabito, R.; Romano, O.; La Spada, G.; Marino, A. H2O2-Induced Oxidative Stress Affects SO4= Transport in Human Erythrocytes. PLoS ONE 2016, 11, e0146485. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Van Zwieten, R.; Verhoeven, A.J.; Roos, D. Inborn defects in the antioxidant systems of human red blood cells. Free Radic. Biol. Med. 2014, 67, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Steck, T.L. The organization of proteins in the human red blood cell membrane. A review. J. Cell Biol. 1974, 62, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Arakawa, T.; Kobayashi-Yurugi, T.; Alguel, Y.; Iwanari, H.; Hatae, H.; Iwata, M.; Abe, Y.; Hino, T.; Ikeda-Suno, C.; Kuma, H.; et al. Crystal structure of the anion exchanger domain of human erythrocyte band 3. Science 2015, 350, 680–684. [Google Scholar] [CrossRef] [PubMed]
- Bennett, V.; Baines, A.J. Spectrin and ankyrin-based pathways: Metazoan inventions for integrating cells into tissues. Physiol. Rev. 2001, 81, 1353–1392. [Google Scholar] [CrossRef] [PubMed]
- Reithmeier, R.A.; Casey, J.R.; Kalli, A.C.; Sansom, M.S.; Alguel, Y.; Iwata, S. Band 3, the human red cell chloride/bicarbonate anion exchanger (AE1, SLC4A1), in a structural context. Biochim. Biophys. Acta 2016, 1858, 1507–1532. [Google Scholar] [CrossRef] [PubMed]
- Sterling, D.; Reithmeier, R.A.; Casey, J.R. A transport metabolon. Functional interaction of carbonic anhydrase II and chloride/bicarbonate exchangers. J. Biol. Chem. 2001, 276, 47886–47894. [Google Scholar] [CrossRef] [PubMed]
- Frumence, E.; Genetet, S.; Ripoche, P.; Iolascon, A.; Andolfo, I.; Le Van Kim, C.; Colin, Y.; Mouro-Chanteloup, I.; Lopez, C. Rapid Cl−/HCO3− exchange kinetics of AE1 in HEK293 cells and hereditary stomatocytosis red blood cells. Am. J. Physiol. Cell Physiol. 2013, 305, C654–C662. [Google Scholar] [CrossRef]
- Hsu, K.; Lee, T.Y.; Periasamy, A.; Kao, F.J.; Li, L.T.; Lin, C.Y.; Lin, H.J.; Lin, M. Adaptable interaction between aquaporin-1 and band 3 reveals a potential role of water channel in blood CO2 transport. FASEB J. 2017, 31, 4256–4264. [Google Scholar] [CrossRef]
- Banerjee, S.; Banerjee, M.; Sarkar, R.N. Diabetes mellitus and aging. J. Indian Med. Assoc. 1998, 96, 147–148. [Google Scholar]
- Saldanha, C. Human Erythrocyte Acetylcholinesterase in Health and Disease. Molecules 2017, 22. [Google Scholar] [CrossRef]
- Anong, W.A.; Franco, T.; Chu, H.; Weis, T.L.; Devlin, E.E.; Bodine, D.M.; An, X.; Mohandas, N.; Low, P.S. Adducin forms a bridge between the erythrocyte membrane and its cytoskeleton and regulates membrane cohesion. Blood 2009, 114, 1904–1912. [Google Scholar] [CrossRef]
- Steck, T.L.; Ramos, B.; Strapazon, E. Proteolytic dissection of band 3, the predominant transmembrane polypeptide of the human erythrocyte membrane. Biochemistry 1976, 15, 1153–1161. [Google Scholar] [CrossRef]
- Tanner, M.J.; Martin, P.G.; High, S. The complete amino acid sequence of the human erythrocyte membrane anion-transport protein deduced from the cDNA sequence. Biochem. J. 1988, 256, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Lux, S.E.; John, K.M.; Kopito, R.R.; Lodish, H.F. Cloning and characterization of band 3, the human erythrocyte anion-exchange protein (AE1). Proc. Natl. Acad. Sci. USA 1989, 86, 9089–9093. [Google Scholar] [CrossRef] [PubMed]
- Kodippili, G.C.; Spector, J.; Hale, J.; Giger, K.; Hughes, M.R.; McNagny, K.M.; Birkenmeier, C.; Peters, L.; Ritchie, K.; Low, P.S. Analysis of the mobilities of band 3 populations associated with ankyrin protein and junctional complexes in intact murine erythrocytes. J. Biol. Chem. 2012, 287, 4129–4138. [Google Scholar] [CrossRef] [PubMed]
- Welbourn, E.M.; Wilson, M.T.; Yusof, A.; Metodiev, M.V.; Cooper, C.E. The mechanism of formation, structure and physiological relevance of covalent hemoglobin attachment to the erythrocyte membrane. Free Radic. Biol. Med. 2017, 103, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Jennings, M.L. Proton fluxes associated with erythrocyte membrane anion exchange. J. Membr. Biol. 1976, 28, 187–205. [Google Scholar] [CrossRef] [PubMed]
- Romano, L.; Passow, H. Characterization of anion transport system in trout red blood cell. Am. J. Physiol. 1984, 246, C330–C338. [Google Scholar] [CrossRef]
- Morabito, R.; Marino, A.; Romano, P.; Rigano, C.; La Spada, G. Sulphate and chloride-dependent potassium transport in human erythrocytes are affected by crude venom from nematocysts of the jellyfish Pelagia noctiluca. Cell. Physiol. Biochem. 2013, 32, 86–95. [Google Scholar] [CrossRef]
- Teti, D.; Crupi, M.; Busa, M.; Valenti, A.; Loddo, S.; Mondello, M.; Romano, L. Chemical and pathological oxidative influences on band 3 protein anion-exchanger. Cell. Physiol. Biochem. 2005, 16, 77–86. [Google Scholar] [CrossRef]
- Morabito, R.R.A.; Bagnato, G.; Neal, R.W.; Sciortino, D.; D’Angelo, T.; Iannelli, F.; Iannelli, F.; Cordova, F.; Cirillo, M.; La Spada, G. Band 3 Protein Function and Oxidative Stress in Erythrocytes from Systemic Sclerosis Patients with Interstitial Lung Disease. Eur. J. Clin. Biomed. Sci. 2017, 3, 80–84. [Google Scholar] [CrossRef]
- Morabito, R.; Remigante, A.; Cavallaro, M.; Taormina, A.; La Spada, G.; Marino, A. Anion exchange through band 3 protein in canine leishmaniasis at different stages of disease. Pflugers Arch. 2017, 469, 713–724. [Google Scholar] [CrossRef]
- Morabito, R.R.A.; Loddo, S.; Trichilo, V.; Dossena, S.; Marino, A. Hyperglycemia affects anion exchange through Band 3 protein: An In Vitro and In Vivo study on human erythrocytes. In Proceedings of the SIBS Società Italiana di Biologia Sperimentale Ancona, Ancona, Italy, 9–10 November 2018. [Google Scholar]
- Gupta, S.C.; Patchva, S.; Koh, W.; Aggarwal, B.B. Discovery of curcumin, a component of golden spice, and its miraculous biological activities. Clin. Exp. Pharmacol. Physiol. 2012, 39, 283–299. [Google Scholar] [CrossRef] [PubMed]
- Kelkel, M.; Jacob, C.; Dicato, M.; Diederich, M. Potential of the dietary antioxidants resveratrol and curcumin in prevention and treatment of hematologic malignancies. Molecules 2010, 15, 7035–7074. [Google Scholar] [CrossRef] [PubMed]
- Morabito, R.; Falliti, G.; Geraci, A.; Spada, G.L.; Marino, A. Curcumin Protects-SH Groups and Sulphate Transport after Oxidative Damage in Human Erythrocytes. Cell. Physiol. Biochem. 2015, 36, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Kamath, V.; Narasimhamurthy, K.; Rajini, P.S. Protective effect of potato peel extract against carbon tetrachloride-induced liver injury in rats. Environ. Toxicol. Pharmacol. 2008, 26, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Heo, S.J.; Park, E.J.; Lee, K.W.; Jeon, Y.J. Antioxidant activities of enzymatic extracts from brown seaweeds. Bioresour. Technol. 2005, 96, 1613–1623. [Google Scholar] [CrossRef]
- Morabito, R.; Remigante, A.; Di Pietro, M.L.; Giannetto, A.; La Spada, G.; Marino, A. SO4= uptake and catalase role in preconditioning after H2O2-induced oxidative stress in human erythrocytes. Pflugers Arch. 2017, 469, 235–250. [Google Scholar] [CrossRef]
- Shan, F.; Yang, R.; Ji, T.; Jiao, F. Vitamin C Inhibits Aggravated Eryptosis by Hydrogen Peroxide in Glucose-6-Phosphated Dehydrogenase Deficiency. Cell. Physiol. Biochem. 2016, 39, 1453–1462. [Google Scholar] [CrossRef]
- Minetti, M.; Mallozzi, C.; Di Stasi, A.M. Peroxynitrite activates kinases of the src family and upregulates tyrosine phosphorylation signaling. Free Radic. Biol. Med. 2002, 33, 744–754. [Google Scholar] [CrossRef]
- Morabito, R.; Remigante, A.; Marino, A. Melatonin Protects Band 3 Protein in Human Erythrocytes against H2O2-Induced Oxidative Stress. Molecules 2019, 24. [Google Scholar] [CrossRef]
- Morabito, R.; Remigante, A.; Marino, A. Protective Role of Magnesium against Oxidative Stress on SO4= Uptake through Band 3 Protein in Human Erythrocytes. Cell. Physiol. Biochem. 2019, 52, 1292–1308. [Google Scholar] [CrossRef]
- Zhu, J.; Kang, L.; Ye, Q.; Fan, G.; Liang, Y.; Yan, C.; Orgah, J. Effects of Shenfu injection and its main components on the contraction of isolated rat thoracic aortic rings. PLoS ONE 2013, 8, e78026. [Google Scholar] [CrossRef] [PubMed]
- Pantaleo, A.; Ferru, E.; Pau, M.C.; Khadjavi, A.; Mandili, G.; Matte, A.; Spano, A.; De Franceschi, L.; Pippia, P.; Turrini, F. Band 3 Erythrocyte Membrane Protein Acts as Redox Stress Sensor Leading to Its Phosphorylation by p (72) Syk. Oxid. Med. Cell. Longev. 2016, 2016, 6051093. [Google Scholar] [CrossRef] [PubMed]
- Bede, O.N.D.; Surányi, A.; Horváth, I.; Szlávik, M.; Gyurkovits, K. Effects of magnesium supplementation on the glutathione redox system in atopic asthmatic children. Inflamm. Res. 2008, 57, 279–286. [Google Scholar] [CrossRef] [PubMed]
- De Franceschi, L.; Brugnara, C.; Beuzard, Y. Dietary magnesium supplementation ameliorates anemia in a mouse model of β-Thalassemia. Blood 2016, 90, 1283–1290. [Google Scholar] [CrossRef]
- Chernyshova, E.S.; Zaikina, Y.S.; Tsvetovskaya, G.A.; Strokotov, D.I.; Yurkin, M.A.; Serebrennikova, E.S.; Volkov, L.; Maltsev, V.P.; Chernyshev, A.V. Influence of magnesium sulfate on HCO3/Cl transmembrane exchange rate in human erythrocytes. J. Theor. Biol. 2016, 393, 194–202. [Google Scholar] [CrossRef] [PubMed]
- De Franceschi, L.V.M.E.; Fumagalli, L.; Brugnara, C.; Turrini, F.; Motta, R.; Veghini, E.; Corato, C.; Alper, S.L.; Berton, G. K-Cl cotransport modulation by intracellular Mg in erythrocytes from mice bred for low and high Mg levels. Am. J. Physiol. Cell Physiol. 2001, 281, C1385–C1395. [Google Scholar] [CrossRef] [PubMed]
- Freedman, A.M.T.; Stafford, R.E.; Dickens, B.F.; Cassidy, M.M.; Muesing, R.A.; Weglicki, W.B. Erythrocytes from mag-nesium-deficient hamsters display an enhanced susceptibility to oxidative stress. Am. J. Physiol. Cell Physiol. 1992, 262, C1371–C1375. [Google Scholar] [CrossRef]
- Crupi, M.; Romano, L.; Romano, P.; Venza, M.; Venza, I.; Teti, D. Erythrocytes anion transport and oxidative change in beta-thalassaemias. Cell Biol. Int. 2010, 34, 655–662. [Google Scholar] [CrossRef]
- Pantaleo, A.; Ferru, E.; Giribaldi, G.; Mannu, F.; Carta, F.; Matte, A.; de Franceschi, L.; Turrini, F. Oxidized and poorly glycosylated band 3 is selectively phosphorylated by Syk kinase to form large membrane clusters in normal and G6PD-deficient red blood cells. Biochem. J. 2009, 418, 359–367. [Google Scholar] [CrossRef]
- Bordin, L.; Fiore, C.; Bragadin, M.; Brunati, A.M.; Clari, G. Regulation of membrane band 3 Tyr-phosphorylation by proteolysis of p72(Syk) and possible involvement in senescence process. Acta Biochim. Biophys. Sin. 2009, 41, 846–851. [Google Scholar] [CrossRef][Green Version]
- Bordin, L.; Ion-Popa, F.; Brunati, A.M.; Clari, G.; Low, P.S. Effector-induced Syk-mediated phosphorylation in human erythrocytes. Biochim. Biophys. Acta 2005, 1745, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Rahman, K. Studies on free radicals, antioxidants, and co-factors. Clin. Interv. Aging 2007, 2, 219–236. [Google Scholar] [PubMed]
- Korkmaz, A.; Reiter, R.J.; Topal, T.; Manchester, L.C.; Oter, S.; Tan, D.X. Melatonin: An established antioxidant worthy of use in clinical trials. Mol. Med. 2009, 15, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Mayo, J.C.; Sainz, R.M.; Leon, J.; Czarnocki, Z. Melatonin as an antioxidant: Biochemical mechanisms and pathophysiological implications in humans. Acta Biochim. Pol. 2003, 50, 1129–1146. [Google Scholar] [CrossRef] [PubMed]
- Radogna, F.; Diederich, M.; Ghibelli, L. Melatonin: A pleiotropic molecule regulating inflammation. Biochem. Pharmacol. 2010, 80, 1844–1852. [Google Scholar] [CrossRef]
- Anisimov, V.N.; Popovich, I.G.; Zabezhinski, M.A.; Anisimov, S.V.; Vesnushkin, G.M.; Vinogradova, I.A. Melatonin as antioxidant, geroprotector and anticarcinogen. Biochim. Biophys. Acta 2006, 1757, 573–589. [Google Scholar] [CrossRef] [PubMed]
- Tesoriere, L.; D’Arpa, D.; Conti, S.; Giaccone, V.; Pintaudi, A.M.; Livrea, M.A. Melatonin protects human red blood cells from oxidative hemolysis: New insights into the radical-scavenging activity. J. Pineal Res. 1999, 27, 95–105. [Google Scholar] [CrossRef]
- Srour, M.A.; Bilto, Y.Y.; Juma, M. Evaluation of different methods used to measure malonyldialdehyde in human erythrocytes. Clin. Hemorheol. Microcirc. 2000, 23, 23–30. [Google Scholar]



{kind=link}
{kind=link}
| Condition | Rate Constant (min−1) | Time (min) | n |
|---|---|---|---|
| Human erythrocytes (ctr) [8,34,37,40,41] | 0.066 ± 0.001 | 18 | 18 |
| Medium pH 6.5 [34] | 0.035 ± 0.001 | 29 | 6 |
| 300 μM H2O2 [37] | 0.032 ± 0.001 | 31 | 6 |
| 600 μM H2O2 [37] | 0.030 ± 0.001 | 33 | 3 |
| 1 mM Diamide [28] | 0.029 ± 0.002 | 32 | 5 |
| 0.1 mM Orthovanadate [28] | 0.031 ± 0.001 | 34 | 5 |
| 0.5 mM NEM [41] | 0.030 ± 0.001 | 33 | 5 |
| 1 mM NEM [41] | 0.033 ± 0.003 | 30 | 5 |
| 2 mM NEM [41] | 0.023 ± 0.002 | 43 | 7 |
| 10 μM Curcumin in Medium pH 6.5 [34] | 0.048 ± 0.001 | 20 | 6 |
| 10 mM Mg2+ 300 μM H2O2 [41] | 0.058 ± 0.005 | 17 | 5 |
| 10 mM Mg2+ 600 μM H2O2 [41] | 0.057 ± 0.001 | 17 | 5 |
| 10 mM Mg2+ 0.5 mM NEM [41] | 0.060 ± 0.002 | 16 | 6 |
| 10 mM Mg2+ 1 mM NEM [41] | 0.056 ± 0.002 | 18 | 6 |
| 10 mM Mg2+ 2 mM NEM [41] | 0.055 ± 0.002 | 18 | 6 |
| 100 μM Melatonin + 300 μM H2O2 [40] | 0.078 ± 0.001 | 13.5 | 10 |
| 10 μM H2O2 (Preconditioning) + 300 μM H2O2 [37] | 0.051 ± 0.001 | 19 | 4 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Remigante, A.; Morabito, R.; Marino, A. Natural Antioxidants Beneficial Effects on Anion Exchange through Band 3 Protein in Human Erythrocytes. Antioxidants 2020, 9, 25. https://doi.org/10.3390/antiox9010025
Remigante A, Morabito R, Marino A. Natural Antioxidants Beneficial Effects on Anion Exchange through Band 3 Protein in Human Erythrocytes. Antioxidants. 2020; 9(1):25. https://doi.org/10.3390/antiox9010025
Chicago/Turabian StyleRemigante, Alessia, Rossana Morabito, and Angela Marino. 2020. "Natural Antioxidants Beneficial Effects on Anion Exchange through Band 3 Protein in Human Erythrocytes" Antioxidants 9, no. 1: 25. https://doi.org/10.3390/antiox9010025
APA StyleRemigante, A., Morabito, R., & Marino, A. (2020). Natural Antioxidants Beneficial Effects on Anion Exchange through Band 3 Protein in Human Erythrocytes. Antioxidants, 9(1), 25. https://doi.org/10.3390/antiox9010025