Qualitative Chemical Characterization and Multidirectional Biological Investigation of Leaves and Bark Extracts of Anogeissus leiocarpus (DC.) Guill. & Perr. (Combretaceae)


 ,
,  ,
,  , ,
, , 


 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Preparation of Extracts
2.2. Profile of Bioactive Compounds
2.3. Determination of Antioxidant and Enzyme Inhibitory Effects
2.4. Pharmacological Assays
2.4.1. Allelopathy Assay
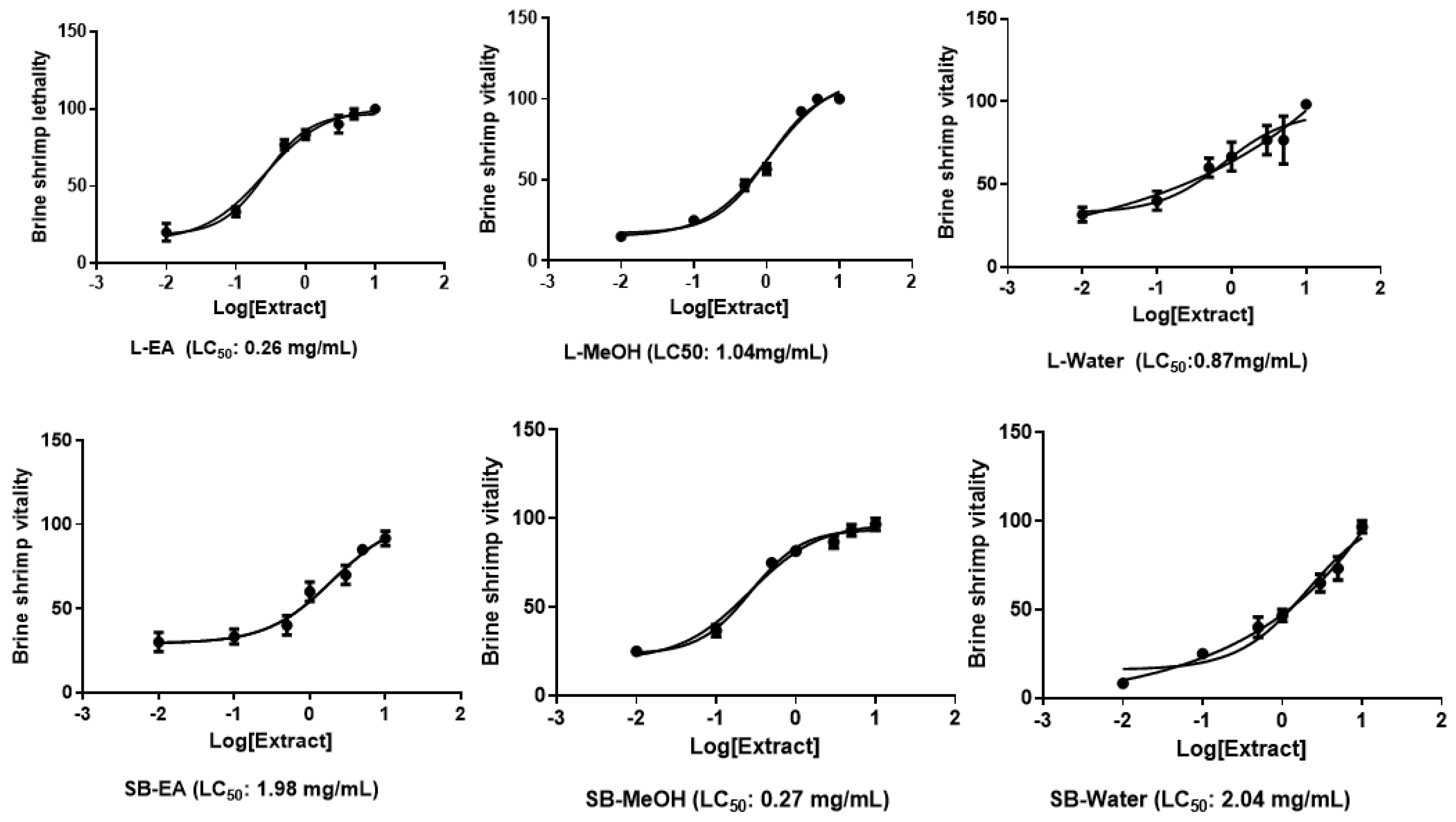
2.4.2. Brine Shrimp Lethality Assay
2.5. In Vitro Studies
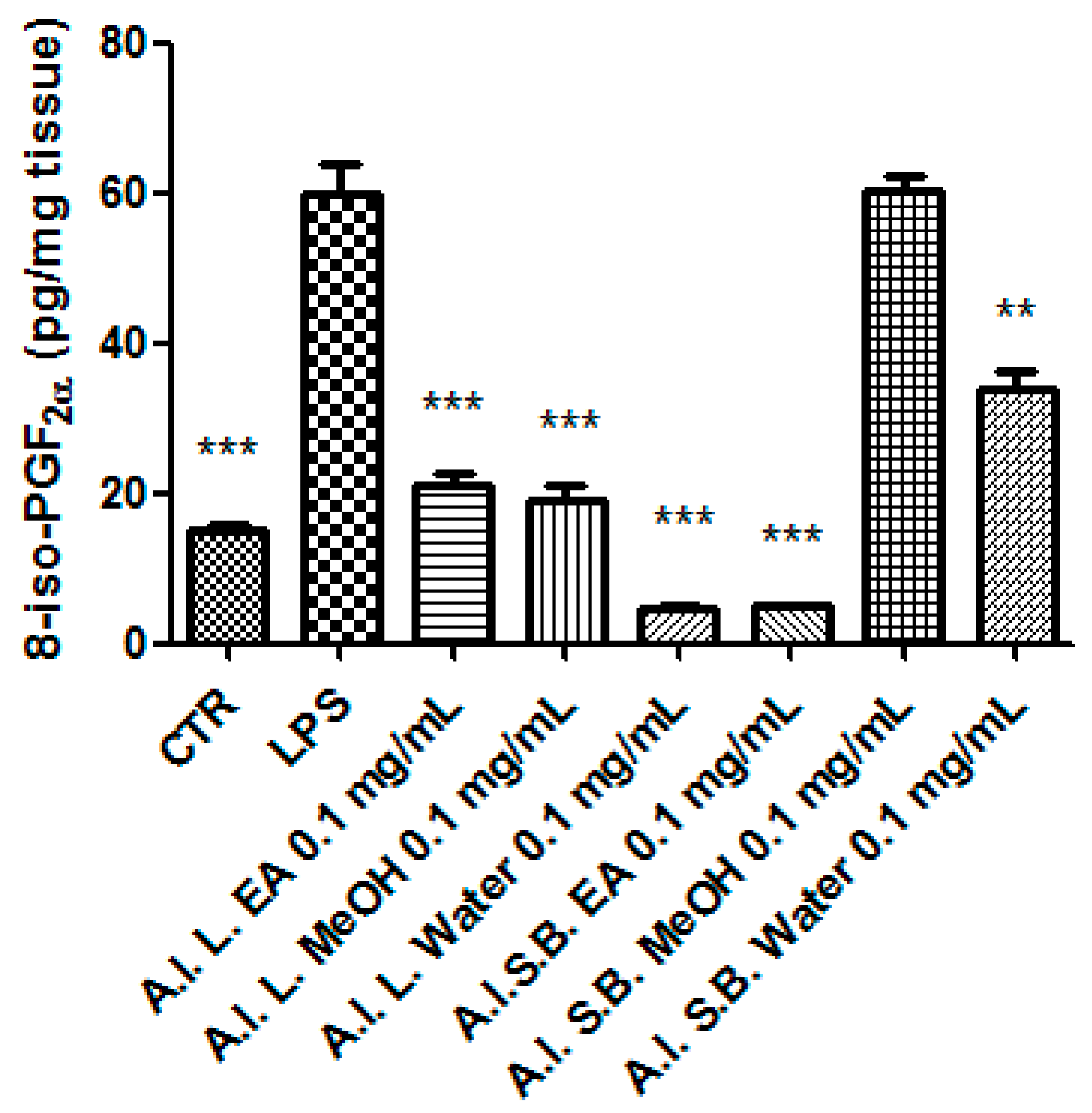
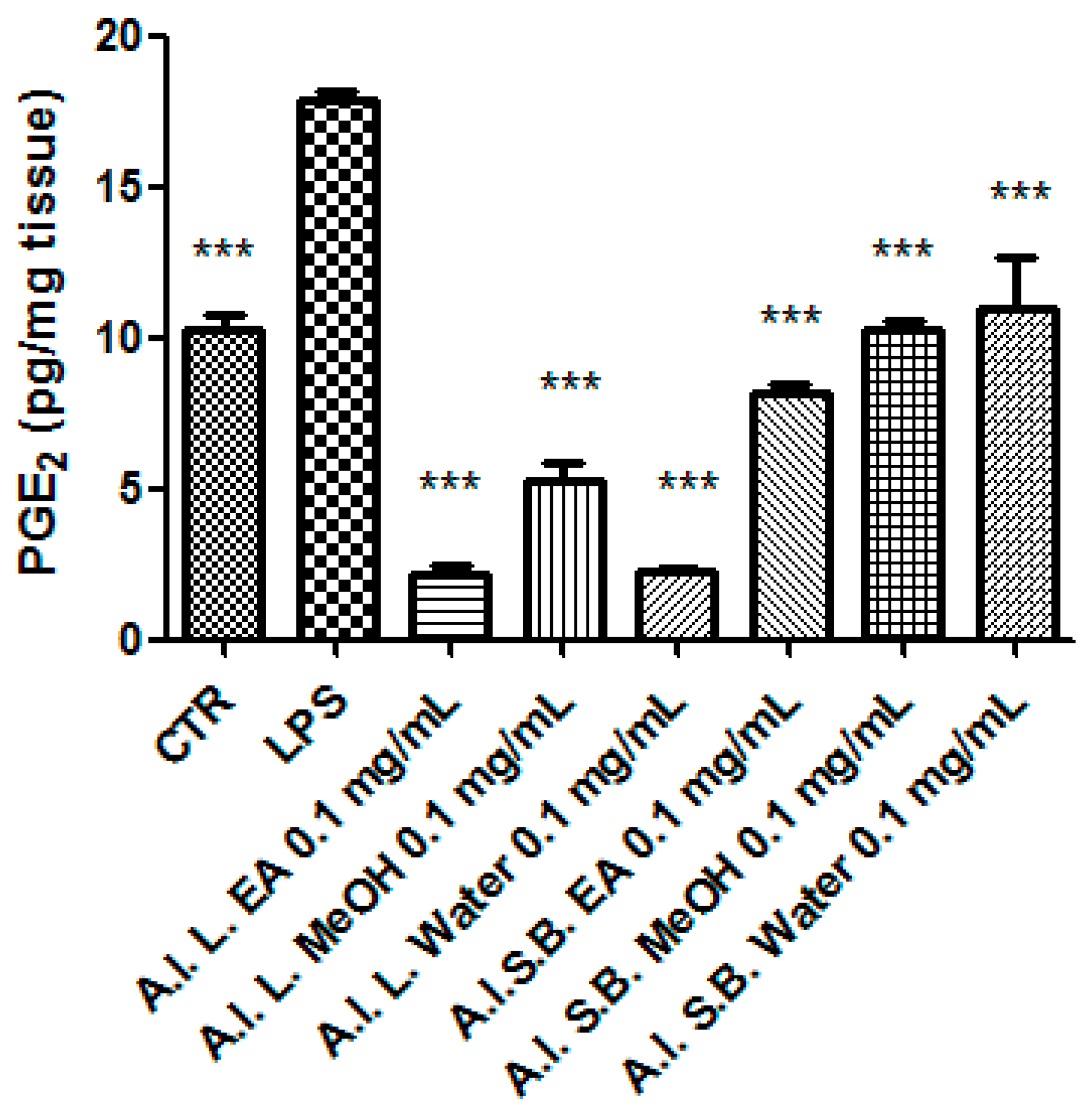
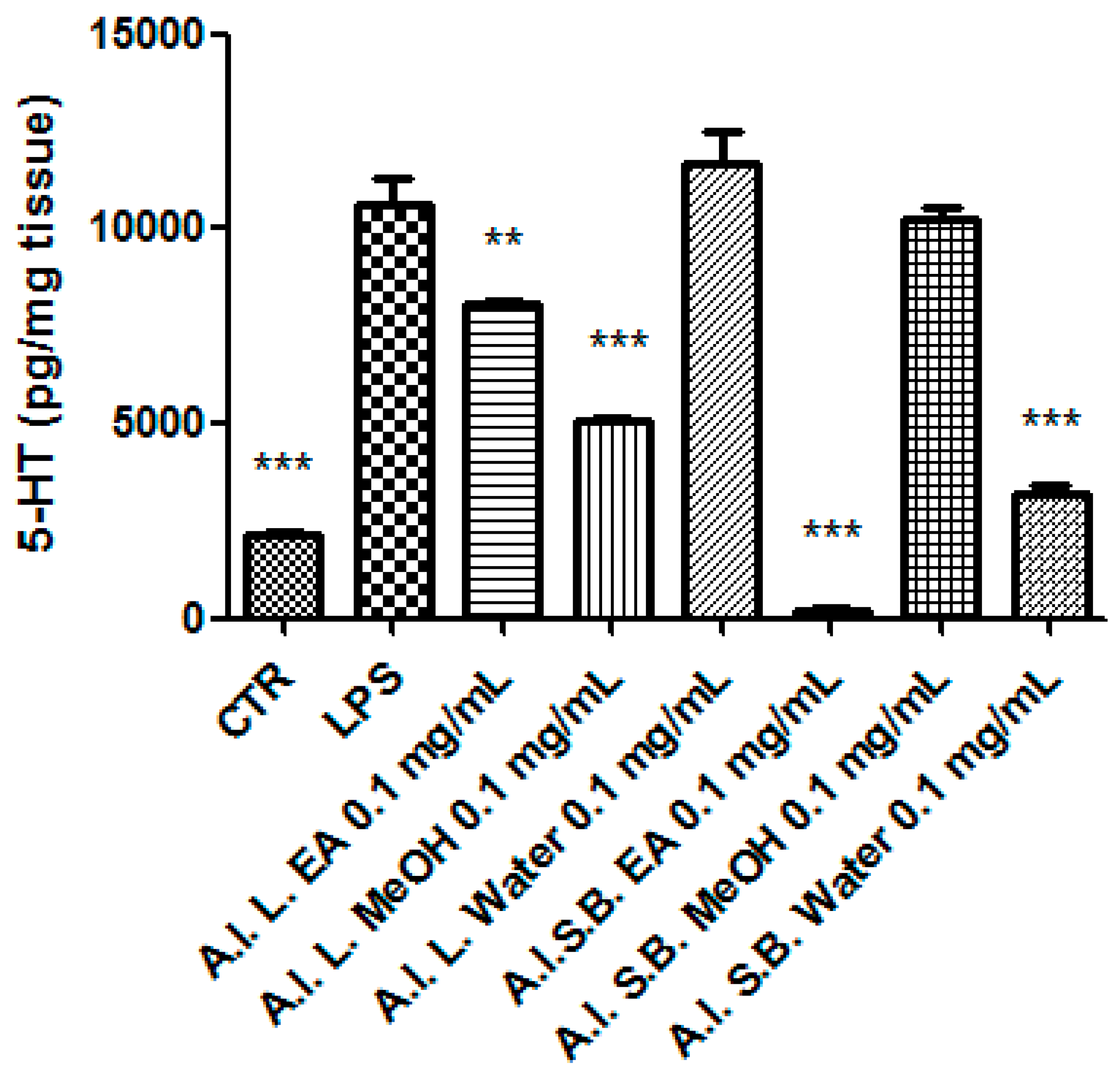
2.6. Ex Vivo Studies
2.7. Statistical Analysis
3. Results
3.1. Phytochemical Profile
3.2. Phenolic Acids
3.3. Flavonoids
3.4. Tannins Derivatives
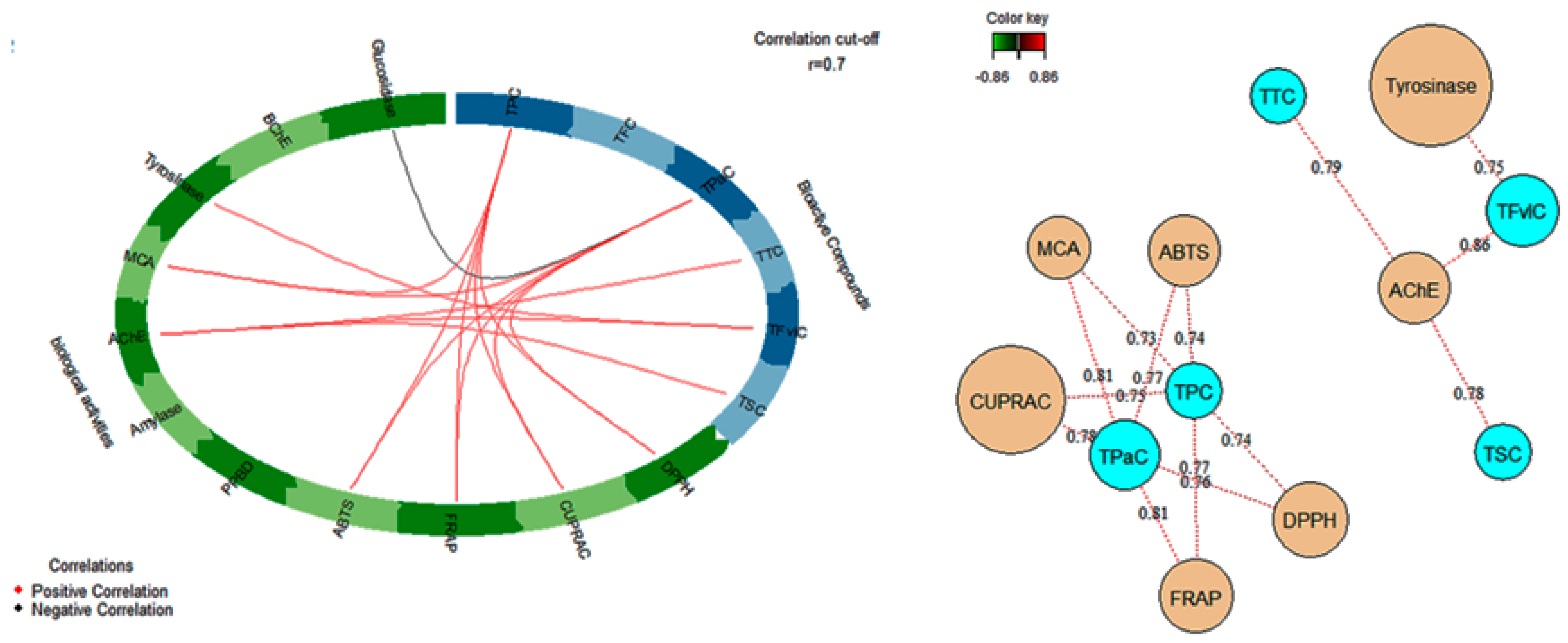
3.5. Antioxidant Activities
3.6. Enzyme Inhibitory Activities
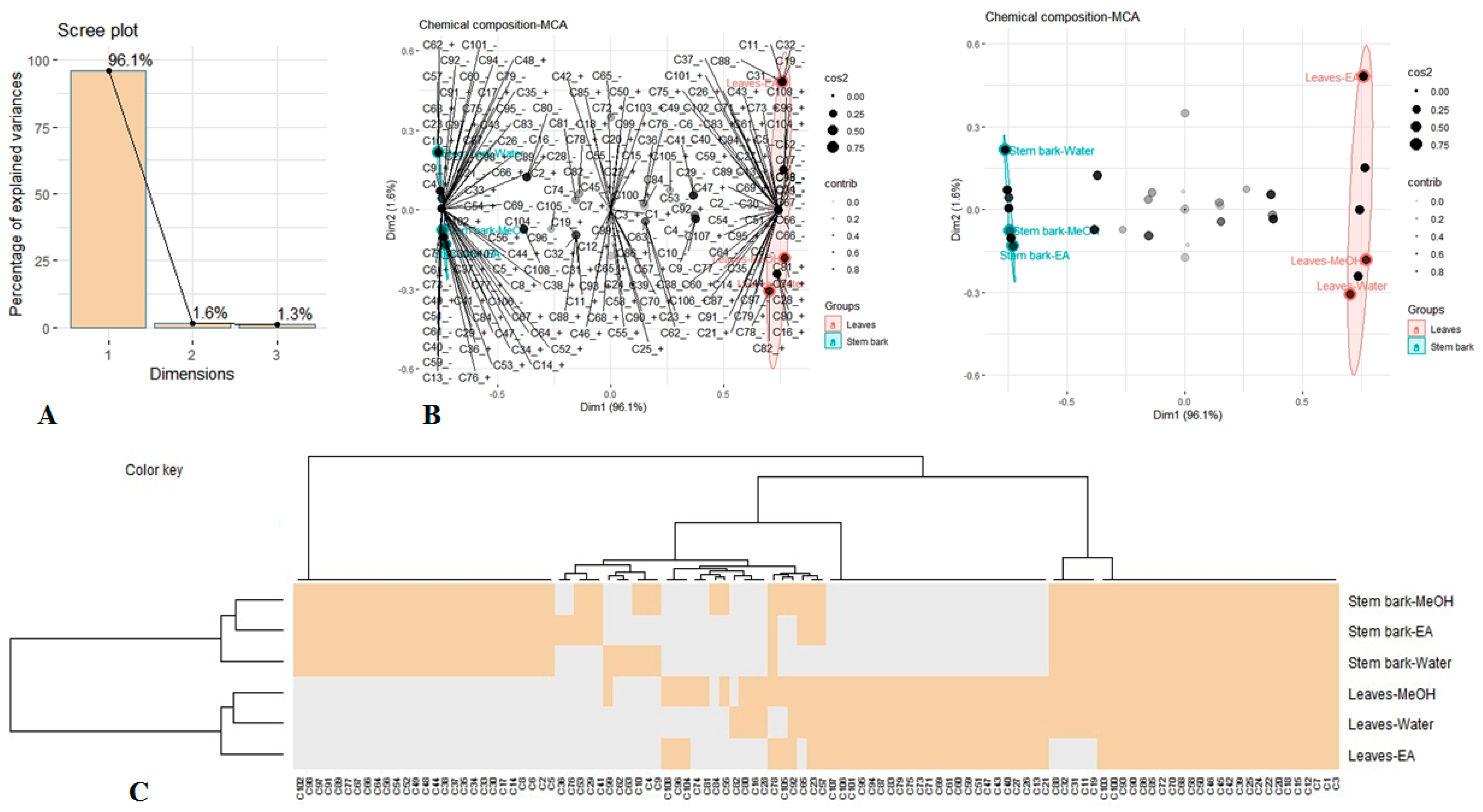
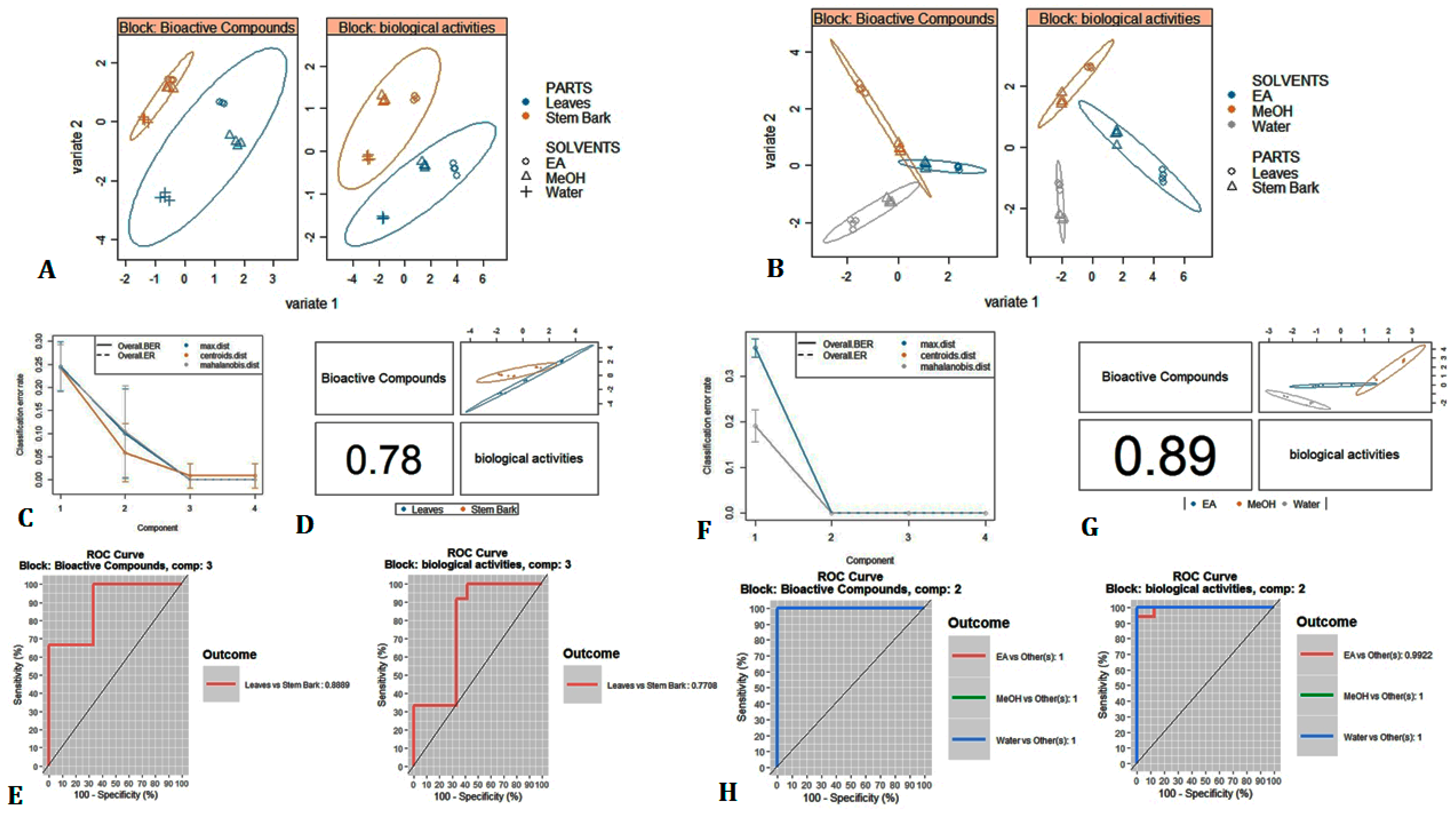
3.7. Multivariate Analysis
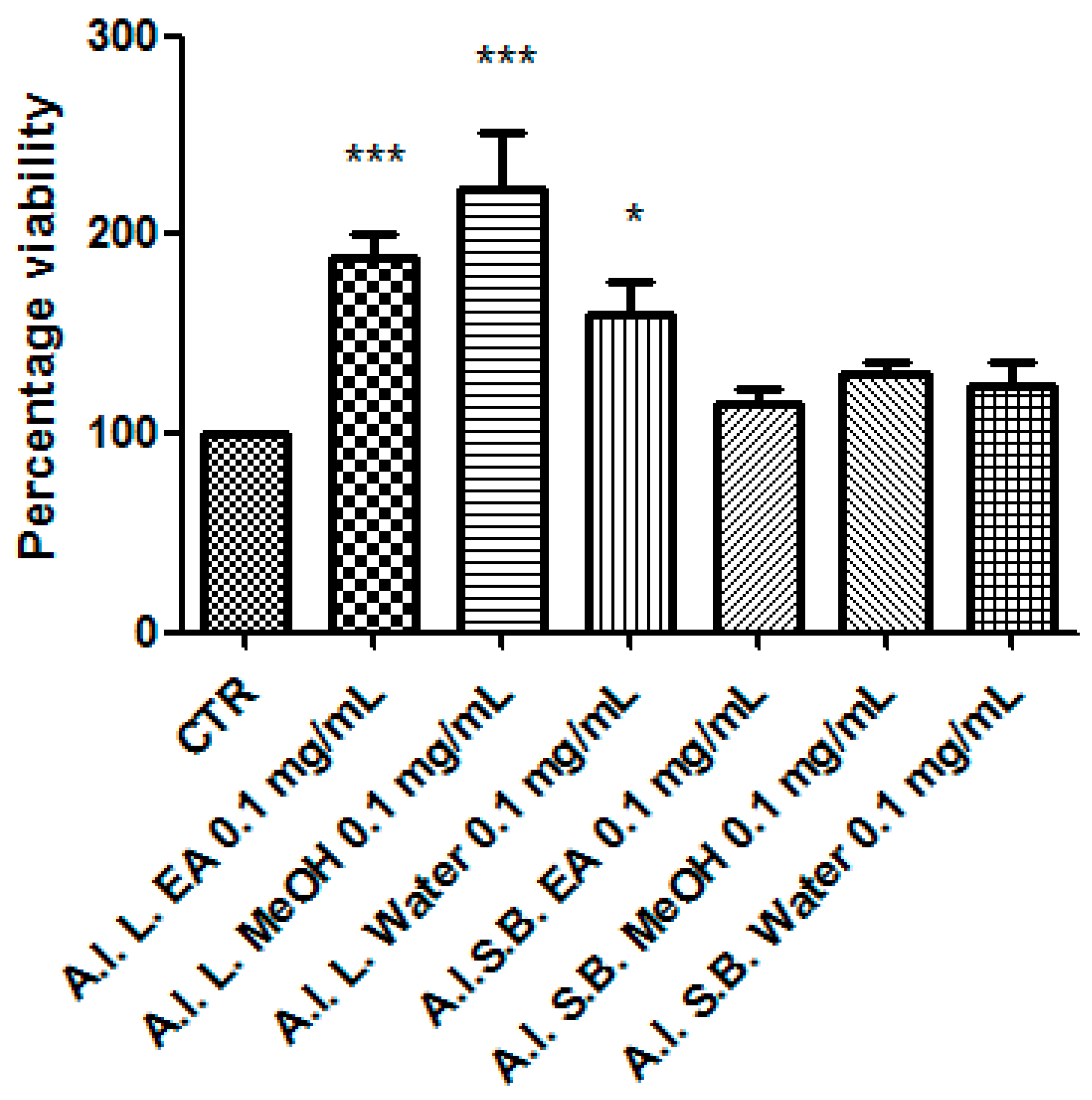
3.8. Pharmacological Studies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Chaabi, M.; Benayache, S.; Benayache, F.; N’Gom, S.; Koné, M.; Anton, R.; Weniger, B.; Lobstein, A. Triterpenes and polyphenols from Anogeissus leiocarpus (Combretaceae). Biochem. Syst. Ecol. 2008, 36, 59–62. [Google Scholar] [CrossRef]
- Salih, E.Y.A.; Kanninen, M.; Sipi, M.; Luukkanen, O.; Hiltunen, R.; Vuorela, H.; Julkunen-Tiitto, R.; Fyhrquist, P. Tannins, flavonoids and stilbenes in extracts of African savanna woodland trees Terminalia brownii, Terminalia laxiflora and Anogeissus leiocarpus showing promising antibacterial potential. S. Afr. J. Bot. 2017, 108, 370–386. [Google Scholar] [CrossRef]
- Ademosun, A.O.; Adebayo, A.A.; Oboh, G. Anogeissus leiocarpus attenuates paroxetine-induced erectile dysfunction in male rats via enhanced sexual behavior, nitric oxide level and antioxidant status. Biomed. Pharmacother. 2019, 111, 1029–1035. [Google Scholar] [CrossRef] [PubMed]
- Etuk, E.; Mohammed, B. Informant consensus selection method: A reliability assessment on medicinal plants used in north western Nigeria for the treatment of diabetes mellitus. Afr. J. Pharm. Pharmacol. 2009, 3, 496–500. [Google Scholar]
- Adefegha, S.A.; Oboh, G.; Omojokun, O.S.; Jimoh, T.O.; Oyeleye, S.I. In vitro antioxidant activities of African birch (Anogeissus leiocarpus) leaf and its effect on the α-amylase and α-glucosidase inhibitory properties of acarbose. J. Taibah Univ. Med. Sci. 2016, 11, 236–242. [Google Scholar] [CrossRef]
- Belemnaba, L.; Nitiema, M.; Ilboudo, S.; Ouedraogo, N.; Ouedraogo, G.G.; Belemlilga, M.B.; Ouedraogo, S.; Guissou, I.P. O8 Study on antihypertensive activity of an aqueous extract of Anogeissus leiocarpus (AEAL) DC Guill et Perr bark of trunk in L-NAME-induced hypertensive rats. Biochem. Pharmacol. 2017, 139, 112. [Google Scholar] [CrossRef]
- Victor, B.Y.; Grace, A. Phytochemical studies, in-vitro antibacterial activities and antioxidant properties of the methanolic and ethyl acetate extracts of the leaves of Anogeissus leiocarpus. Int. J. Biochem. Res. Rev. 2013, 3, 137. [Google Scholar] [CrossRef]
- Shuaibu, M.N.; Wuyep, P.T.; Yanagi, T.; Hirayama, K.; Ichinose, A.; Tanaka, T.; Kouno, I. Trypanocidal activity of extracts and compounds from the stem bark of Anogeissus leiocarpus and Terminalia avicennoides. Parasitol. Res. 2008, 102, 697–703. [Google Scholar] [CrossRef]
- Shuaibu, M.N.; Pandey, K.; Wuyep, P.A.; Yanagi, T.; Hirayama, K.; Ichinose, A.; Tanaka, T.; Kouno, I. Castalagin from Anogeissus leiocarpus mediates the killing of Leishmania in vitro. Parasitol. Res. 2008, 103, 1333–1338. [Google Scholar] [CrossRef]
- Vonthron-Sénécheau, C.; Weniger, B.; Ouattara, M.; Bi, F.T.; Kamenan, A.; Lobstein, A.; Brun, R.; Anton, R. In vitro antiplasmodial activity and cytotoxicity of ethnobotanically selected Ivorian plants. J. Ethnopharmacol. 2003, 87, 221–225. [Google Scholar] [CrossRef]
- Akanbi, O.M.; Omonkhua, A.A.; Cyril-Olutayo, C.M. Effect of crude methanolic extract of Anogeissus leiocarpus on the liver function of P. berghei infected mice. Int. J. Infect. Dis. 2014, 21, 196. [Google Scholar] [CrossRef][Green Version]
- Diab, K.A.; Guru, S.K.; Bhushan, S.; Saxena, A.K. In vitro anticancer activities of Anogeissus latifolia, Terminalia bellerica, Acacia catechu and Moringa oleiferna Indian plants. Asian Pac. J. Cancer Prev. 2015, 16, 6423–6428. [Google Scholar] [CrossRef] [PubMed]
- Ferrante, C.; Recinella, L.; Ronci, M.; Menghini, L.; Brunetti, L.; Chiavaroli, A.; Leone, S.; Di Iorio, L.; Carradori, S.; Tirillini, B.; et al. Multiple pharmacognostic characterization on hemp commercial cultivars: Focus on inflorescence water extract activity. Food Chem. Toxicol. 2019, 125, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Zengin, G.; Aktumsek, A. Investigation of antioxidant potentials of solvent extracts from different anatomical parts of Asphodeline anatolica E. Tuzlaci: An endemic plant to Turkey. Afr. J. Tradit. Complement. Altern. Med. 2014, 11, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Zengin, G.; Llorent-Martínez, E.J.; Fernández-de Córdova, M.L.; Bahadori, M.B.; Mocan, A.; Locatelli, M.; Aktumsek, A. Chemical composition and biological activities of extracts from three Salvia species: S. blepharochlaena, S. euphratica var. leiocalycina, and S. verticillata subsp. amasiaca. Ind. Crops Prod. 2018, 111, 11–21. [Google Scholar] [CrossRef]
- Zengin, G.; Uysal, A.; Diuzheva, A.; Gunes, E.; Jekő, J.; Cziáky, Z.; Picot-Allain, C.M.N.; Mahomoodally, M.F. Characterization of phytochemical components of Ferula halophila extracts using HPLC-MS/MS and their pharmacological potentials: A multi-functional insight. J. Pharm. Biomed. Anal. 2018, 160, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Rodrıguez-Delgado, M.; Malovana, S.; Perez, J.; Borges, T.; Montelongo, F.G. Separation of phenolic compounds by high-performance liquid chromatography with absorbance and fluorimetric detection. J. Chromatogr. A 2001, 912, 249–257. [Google Scholar] [CrossRef]
- Uysal, S.; Zengin, G.; Locatelli, M.; Bahadori, M.B.; Mocan, A.; Bellagamba, G.; De Luca, E.; Mollica, A.; Aktumsek, A. Cytotoxic and enzyme inhibitory potential of two Potentilla species (P. speciosa L. and P. reptans Willd.) and their chemical composition. Front. Pharmacol. 2017, 8, 290. [Google Scholar] [CrossRef]
- Mahmoodzadeh, H.; GhasemI, M.; Zanganeh, H. Allelopathic effect of medicinal plant Cannabis sativa L. on Lactuca sativa L. seed germination. Acta Agric. Slov. 2015, 105, 233–239. [Google Scholar] [CrossRef]
- Brunetti, L.; Orlando, G.; Ferrante, C.; Recinella, L.; Leone, S.; Chiavaroli, A.; Di Nisio, C.; Shohreh, R.; Manippa, F.; Ricciuti, A.; et al. Peripheral chemerin administration modulates hypothalamic control of feeding. Peptides 2014, 51, 115–121. [Google Scholar] [CrossRef]
- Ferrante, C.; Orlando, G.; Recinella, L.; Leone, S.; Chiavaroli, A.; Di Nisio, C.; Shohreh, R.; Manippa, F.; Ricciuti, A.; Vacca, M.; et al. Central inhibitory effects on feeding induced by the adipo-myokine irisin. Eur. J. Pharmacol. 2016, 791, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Chiavaroli, A.; Brunetti, L.; Orlando, G.; Recinella, L.; Ferrante, C.; Leone, S.; Di Michele, P.; Di Nisio, C.; Vacca, M. Resveratrol inhibits isoprostane production in young and aged rat brain. J. Biol. Regul. Homeost. Agents 2010, 24, 441. [Google Scholar] [PubMed]
- Locatelli, M.; Macchione, N.; Ferrante, C.; Chiavaroli, A.; Recinella, L.; Carradori, S.; Zengin, G.; Cesa, S.; Leporini, L.; Leone, S. Graminex pollen: Phenolic pattern, colorimetric analysis and protective effects in immortalized prostate cells (PC3) and rat prostate challenged with LPS. Molecules 2018, 23, 1145. [Google Scholar] [CrossRef] [PubMed]
- National Centre for Replacement Refinement& Reduction of Animals in Research. Available online: https://www.nc3rs.org.uk/experimental-designstatistics (accessed on 16 August 2019).
- Galanakis, C.M.; Goulas, V.; Tsakona, S.; Manganaris, G.A.; Gekas, V. A Knowledge Base for the Recovery of Natural Phenols with Different Solvents. Int. J. Food Prop. 2013, 16, 382–396. [Google Scholar] [CrossRef]
- Ji, W.; Meng, Q.; Ding, L.; Wang, F.; Dong, J.; Zhou, G.; Wang, B. Measurement and correlation of the solubility of caffeic acid in eight mono and water+ethanol mixed solvents at temperatures from (293.15 to 333.15) K. J. Mol. Liq. 2016, 224, 1275–1281. [Google Scholar] [CrossRef]
- Lin, L.-T.; Chen, T.-Y.; Chung, C.-Y.; Noyce, R.S.; Grindley, T.B.; McCormick, C.; Lin, T.-C.; Wang, G.-H.; Lin, C.-C.; Richardson, C.D. Hydrolyzable Tannins (Chebulagic Acid and Punicalagin) Target Viral Glycoprotein-Glycosaminoglycan Interactions To Inhibit Herpes Simplex Virus 1 Entry and Cell-to-Cell Spread. J. Virol. 2011, 85, 4386–4398. [Google Scholar] [CrossRef] [PubMed]
- Perera, H.D.S.M.; Samarasekera, J.K.R.R.; Handunnetti, S.M.; Weerasena, O.V.D.S.J. In vitro anti-inflammatory and anti-oxidant activities of Sri Lankan medicinal plants. Ind. Crops Prod. 2016, 94, 610–620. [Google Scholar] [CrossRef]
- Holtz, R.W. Chapter 13—In Vitro Methods to Screen Materials for Anti-aging Effects. In Skin Aging Handbook; Dayan, N., Ed.; William Andrew Publishing: Norwich, NY, USA, 2009; pp. 329–362. [Google Scholar]
- Cerretani, L.; Bendini, A. Chapter 67—Rapid Assays to Evaluate the Antioxidant Capacity of Phenols in Virgin Olive Oil. In Olives and Olive Oil in Health and Disease Prevention; Preedy, V.R., Watson, R.R., Eds.; Academic Press: San Diego, CA, USA, 2010; pp. 625–635. [Google Scholar]
- Özyürek, M.; Güçlü, K.; Tütem, E.; Başkan, K.S.; Erçağ, E.; Celik, S.E.; Baki, S.; Yıldız, L.; Karaman, Ş.; Apak, R. A comprehensive review of CUPRAC methodology. Anal. Methods 2011, 3, 2439–2453. [Google Scholar] [CrossRef]
- Puntarulo, S. Iron, oxidative stress and human health. Mol. Asp. Med. 2005, 26, 299–312. [Google Scholar] [CrossRef]
- Mira, L.; Fernandez, M.T.; Santos, M.; Rocha, R.; Florencio, M.H.; Jennings, K.R. Interactions of flavonoids with iron and copper ions: A mechanism for their antioxidant activity. Free Radic. Res. 2002, 36, 1199–1208. [Google Scholar] [CrossRef]
- Mandel, S.; Amit, T.; Reznichenko, L.; Weinreb, O.; Youdim, M.B. Green tea catechins as brain-permeable, natural iron chelators-antioxidants for the treatment of neurodegenerative disorders. Mol. Nutr. Food Res. 2006, 50, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Guo, M. Studies on Transition Metal-Quercetin Complexes Using Electrospray Ionization Tandem Mass Spectrometry. Molecules 2015, 20, 8583–8594. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.A.; Chand, K.; Chaves, S. Recent progress in multifunctional metal chelators as potential drugs for Alzheimer’s disease. Coord. Chem. Rev. 2016, 327, 287–303. [Google Scholar] [CrossRef]
- Ragab, H.M.; Teleb, M.; Haidar, H.R.; Gouda, N. Chlorinated tacrine analogs: Design, synthesis and biological evaluation of their anti-cholinesterase activity as potential treatment for Alzheimer’s disease. Bioorg. Chem. 2019, 86, 557–568. [Google Scholar] [CrossRef] [PubMed]
- de Andrade, P.; Mantoani, S.P.; Gonçalves Nunes, P.S.; Magadán, C.R.; Pérez, C.; Xavier, D.J.; Hojo, E.T.S.; Campillo, N.E.; Martínez, A.; Carvalho, I. Highly potent and selective aryl-1,2,3-triazolyl benzylpiperidine inhibitors toward butyrylcholinesterase in Alzheimer’s disease. Bioorg. Med. Chem. 2019, 27, 931–943. [Google Scholar] [CrossRef] [PubMed]
- Liyanaarachchi, G.D.; Samarasekera, J.K.R.R.; Mahanama, K.R.R.; Hemalal, K.D.P. Tyrosinase, elastase, hyaluronidase, inhibitory and antioxidant activity of Sri Lankan medicinal plants for novel cosmeceuticals. Ind. Crops Prod. 2018, 111, 597–605. [Google Scholar] [CrossRef]
- Wang, R.; Wang, G.; Xia, Y.; Sui, W.; Si, C. Functionality study of lignin as a tyrosinase inhibitor: Influence of lignin heterogeneity on anti-tyrosinase activity. Int. J. Biol. Macromol. 2019, 128, 107–113. [Google Scholar] [CrossRef]
- Chen, Y.H.; Huang, L.; Wen, Z.H.; Zhang, C.; Liang, C.H.; Lai, S.T.; Luo, L.Z.; Wang, Y.Y.; Wang, G.H. Skin whitening capability of shikimic acid pathway compound. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 1214–1220. [Google Scholar]
- Mapunya, M.B.; Hussein, A.A.; Rodriguez, B.; Lall, N. Tyrosinase activity of Greyia flanaganii (Bolus) constituents. Phytomedicine 2011, 18, 1006–1012. [Google Scholar] [CrossRef]
- Yao, Y.; Cheng, X.; Wang, L.; Wang, S.; Ren, G. Mushroom tyrosinase inhibitors from mung bean (Vigna radiatae L.) extracts. Int. J. Food Sci. Nutr. 2012, 63, 358–361. [Google Scholar] [CrossRef]
- Ohikhena, F.U.; Wintola, O.A.; Afolayan, A.J. Toxicity Assessment of Different Solvent Extracts of the Medicinal Plant, Phragmanthera capitata (Sprengel) Balle on Brine Shrimp (Artemia salina). Int. J. Pharmacol. 2016, 12, 701–710. [Google Scholar]
- Taviano, M.F.; Marino, A.; Trovato, A.; Bellinghieri, V.; Melchini, A.; Dugo, P.; Cacciola, F.; Donato, P.; Mondello, L.; Güvenç, A. Juniperus oxycedrus L. subsp. oxycedrus and Juniperus oxycedrus L. subsp. macrocarpa (Sibth. & Sm.) Ball. “berries” from Turkey: Comparative evaluation of phenolic profile, antioxidant, cytotoxic and antimicrobial activities. Food Chem. Toxicol. 2013, 58, 22–29. [Google Scholar] [PubMed]
- Belemnaba, L.; Ouédraogo, S.; Nitiéma, M.; Chataigneau, T.; Guissou, I.P.; Schini-Kerth, V.B.; Bucher, B.; Auger, C. An aqueous extract of the Anogeissus leiocarpus bark (AEAL) induces the endothelium-dependent relaxation of porcine coronary artery rings involving predominantly nitric oxide. J. Basic Clin. Physiol. Pharmacol. 2018, 29, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Locatelli, M.; Ferrante, C.; Carradori, S.; Secci, D.; Leporini, L.; Chiavaroli, A.; Leone, S.; Recinella, L.; Orlando, G.; Martinotti, S. Optimization of aqueous extraction and biological activity of Harpagophytum procumbens root on ex vivo rat colon inflammatory model. Phytother. Res. 2017, 31, 937–944. [Google Scholar] [CrossRef] [PubMed]
- Menghini, L.; Ferrante, C.; Leporini, L.; Recinella, L.; Chiavaroli, A.; Leone, S.; Pintore, G.; Vacca, M.; Orlando, G.; Brunetti, L. An hydroalcoholic chamomile extract modulates inflammatory and immune response in HT29 cells and isolated rat colon. Phytother. Res. 2016, 30, 1513–1518. [Google Scholar] [CrossRef]
- Uttara, B.; Singh, A.V.; Zamboni, P.; Mahajan, R. Oxidative stress and neurodegenerative diseases: A review of upstream and downstream antioxidant therapeutic options. Curr. Neuropharmacol. 2009, 7, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Achitei, D.; Ciobica, A.; Balan, G.; Gologan, E.; Stanciu, C.; Stefanescu, G. Different profile of peripheral antioxidant enzymes and lipid peroxidation in active and non-active inflammatory bowel disease patients. Dig. Dis. Sci. 2013, 58, 1244–1249. [Google Scholar] [CrossRef] [PubMed]
- Praticò, D. Alzheimer’s disease and oxygen radicals: New insights. Biochem. Pharmacol. 2002, 63, 563–567. [Google Scholar] [CrossRef]
- Brunetti, L.; Menghini, L.; Orlando, G.; Recinella, L.; Leone, S.; Epifano, F.; Lazzarin, F.; Chiavaroli, A.; Ferrante, C.; Vacca, M. Antioxidant effects of garlic in young and aged rat brain in vitro. J. Med. Food 2009, 12, 1166–1169. [Google Scholar] [CrossRef]
- Akanbi, O.M.; Omonkhua, A.A.; Cyril-Olutayo, C.M.; Fasimoye, R.Y. The antiplasmodial activity of Anogeissus leiocarpus and its effect on oxidative stress and lipid profile in mice infected with Plasmodium bergheii. Parasitol. Res. 2012, 110, 219–226. [Google Scholar] [CrossRef]
- Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Yan, J.; Sun, Z. D-limonene exhibits anti-inflammatory and antioxidant properties in an ulcerative colitis rat model via regulation of iNOS, COX-2, PGE2 and ERK signaling pathways. Mol. Med. Rep. 2017, 15, 2339–2346. [Google Scholar] [CrossRef] [PubMed]
- Regmi, S.C.; Park, S.-Y.; Ku, S.K.; Kim, J.-A. Serotonin regulates innate immune responses of colon epithelial cells through Nox2-derived reactive oxygen species. Free Radic. Biol. Med. 2014, 69, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Mousavizadeh, K.; Rahimian, R.; Fakhfouri, G.; Aslani, F.; Ghafourifar, P. Anti-inflammatory effects of 5-HT3 receptor antagonist, tropisetron on experimental colitis in rats. Eur. J. Clin. Investig. 2009, 39, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Bungo, T.; Shiraishi, J.-i.; Yanagita, K.; Ohta, Y.; Fujita, M. Effect of nociceptin/orphanin FQ on feeding behavior and hypothalamic neuropeptide expression in layer-type chicks. Gen. Comp. Endocrinol. 2009, 163, 47–51. [Google Scholar] [CrossRef]
- Clark, K.A.; MohanKumar, S.M.; Kasturi, B.S.; MohanKumar, P.S. Effects of central and systemic administration of leptin on neurotransmitter concentrations in specific areas of the hypothalamus. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, R306–R312. [Google Scholar] [CrossRef] [PubMed]
- Salem, I.B.; Boussabbeh, M.; Graiet, I.; Rhouma, A.; Bacha, H.; Essefi, S.A. Quercetin protects HCT116 cells from Dichlorvos-induced oxidative stress and apoptosis. Cell Stress Chaperones 2016, 21, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, K.; Ruan, M.; Yang, J.; Gao, Z. Gallic acid inhibits fibroblast growth and migration in keloids through the AKT/ERK signaling pathway. Acta Biochim. Biophys. Sin. 2018, 50, 1114–1120. [Google Scholar] [CrossRef]
- Sui, X.; Zhu, J.; Tang, H.; Wang, C.; Zhou, J.; Han, W.; Wang, X.; Fang, Y.; Xu, Y.; Li, D.; et al. p53 controls colorectal cancer cell invasion by inhibiting the NF-κB-mediated activation of Fascin. Oncotarget 2015, 6, 22869–22879. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
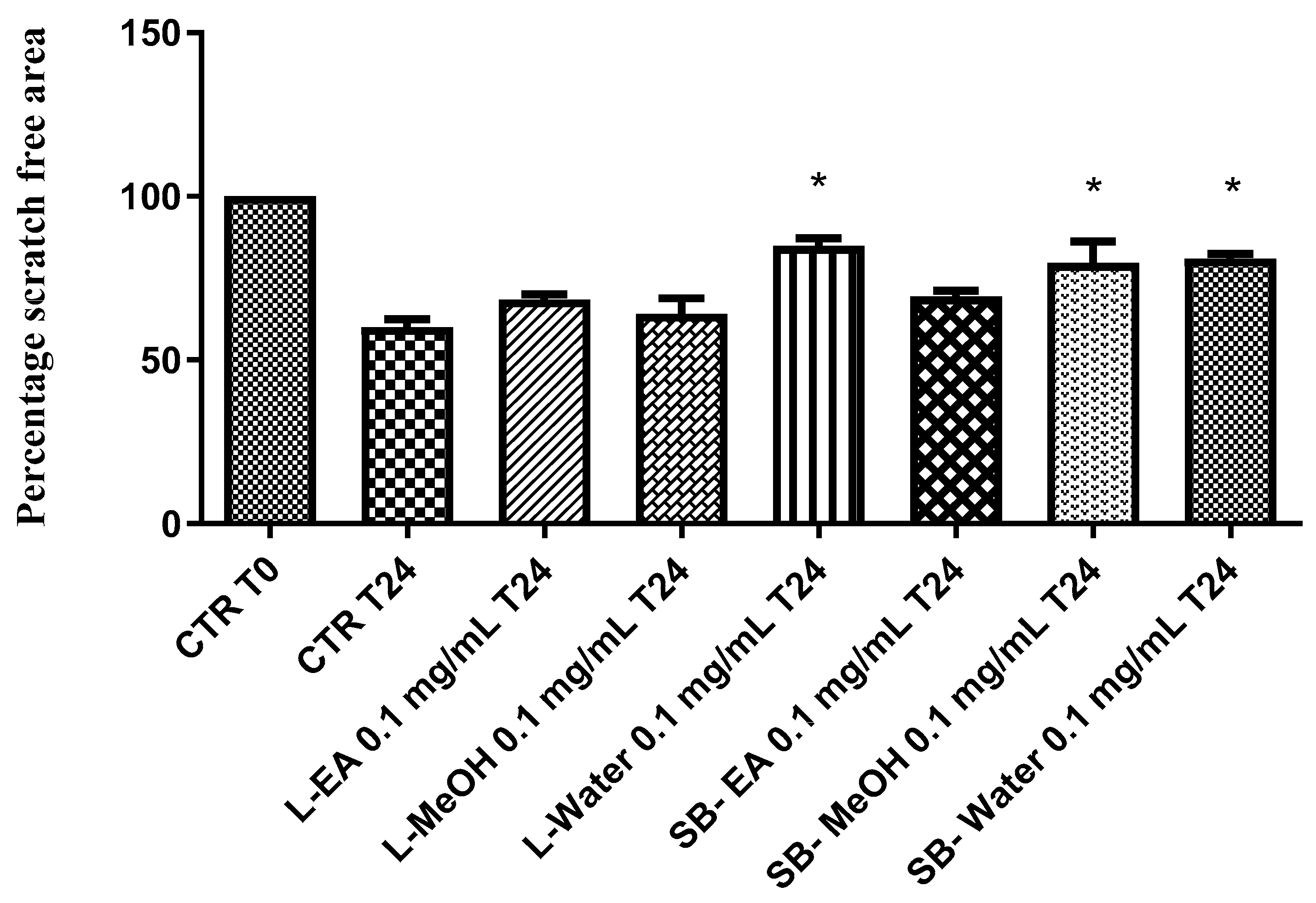{kind=link}
| Samples | Total Phenolic Content (mg GAE/g) | Total Flavonoid Content (mg RE/g) | Total Flavonol (mg CE/g) | Total Phenolic acid (mg CAE/g) | Total Tannin Content (mg CE/g) | Total Saponin Content (mgQE/g) |
|---|---|---|---|---|---|---|
| Leaves-EA | 49 ± 1e | 35.0 ± 0.6c | 6.0 ± 0.1d | nd | 6.0 ± 0.4e | 190 ± 17bc |
| Leaves-MeOH | 223 ± 2c | 54.0 ± 0.6b | 79 ± 3a | 8 ± 1b | 77.0 ± 0.7a | 438 ± 54a |
| Leaves-Water | 257 ± 3b | 89.0 ± 0.2a | 3.46 ± 0.02e | 14 ± 1a | 18 ± 4c | 200 ± 30bc |
| Stem barks-EA | 207 ± 2d | 16 ± 0.3f | 14.0 ± 0.3c | nd | 17 ± 0.5c | 171 ± 24bc |
| Stem barks-MeOH | 271 ± 1a | 27.0 ± 0.3e | 28.0 ± 0.4b | nd | 33 ± 1b | 230 ± 34b |
| Stem barks-Water | 274 ± 2a | 33.0 ± 0.2d | 2.09 ± 0.01e | 7.0 ± 0.6c | 10.0 ± 0.1d | 163 ± 28c |
| Phenolic Compound | Leaves-EA mg/g | Leaves-MeOH mg/g | Leaves-Water Extract mg/g | Stem Barks-EA mg/g | Stem Barks MeOH mg/g | Stem Barks-Water mg/g |
|---|---|---|---|---|---|---|
| Gallic acid | 226 ± 21 | 89 ± 28 | 30 ± 3 | 65 ± 6 | 38 ± 2 | 37 ± 1 |
| Catechin | 7.0 ± 0.8 | 3.0 ± 0.2 | 9 ± 1 | 1.0 ± 0.1 | 0.29 ± 0.02 | 0.28 ± 0.02 |
| Epicatechin | 0.29 ± 0.02 | 2.0 ± 0.1 | 0.27 ± 0.02 | 0.27 ± 0.02 | 0.18 ± 0.01 | 1.0 ± 0.1 |
| No. | Name | Formula | [M + H]+ | [M – H]– | Leaves-EA | Leaves-MeOH | Leaves-Water | Stem Bark-EA | Stem Bark-MeOH | Stem Bark-Water |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Quinic acid | C7H12O6 | 19,105,557 | + | + | + | + | + | + | |
| 2 | Hexahydroxydiphenoylhexose | C20H18O14 | 48,106,184 | – | – | – | + | + | + | |
| 3 | Shikimic acid | C7H10O5 | 17,304,500 | + | + | + | + | + | + | |
| 4 | Galloylquinic acid isomer 1 | C14H16O10 | 34,306,653 | – | – | – | – | + | + | |
| 5 | Galloylhexose isomer 1 | C13H16O10 | 33,106,653 | – | – | – | + | + | + | |
| 6 | Galloylhexose isomer 2 | C13H16O10 | 33,106,653 | – | – | – | + | + | + | |
| 71 | Gallic acid (3,4,5-Trihydroxybenzoic acid) | C7H6O5 | 16,901,370 | + | + | + | + | + | + | |
| 8 | Galloylhexose isomer 3 | C13H16O10 | 33,106,653 | – | – | – | + | + | + | |
| 9 | Galloylquinic acid isomer 2 | C14H16O10 | 34,306,653 | – | – | – | – | + | + | |
| 10 | Galloylquinic acid isomer 3 | C14H16O10 | 34,306,653 | – | – | – | – | + | + | |
| 11 | Gallocatechin | C15H14O7 | 30,506,613 | – | + | + | + | + | + | |
| 12 | Protocatechuic acid (3,4-Dihydroxybenzoic acid) | C7H6O4 | 15,301,879 | + | + | + | + | + | + | |
| 13 | 3-Hydroxybenzaldehyde | C7H6O2 | 12,304,461 | + | + | + | – | – | – | |
| 14 | Procyanidin B isomer 1 | C30H26O12 | 57,713,460 | – | – | – | + | + | + | |
| 15 | Punicalagin | C48H28O30 | 108,305,872 | + | + | + | + | + | + | |
| 16 | Kynurenic acid | C10H7NO3 | 19,005,042 | – | + | + | – | – | – | |
| 17 | Procyanidin B isomer 2 | C30H26O12 | 57,713,460 | – | – | – | + | + | + | |
| 181 | Catechin | C15H14O6 | 28,907,121 | + | + | + | + | + | + | |
| 191 | Epigallocatechin | C15H14O7 | 30,506,613 | – | + | + | + | + | + | |
| 20 | Casuarinin | C41H28O26 | 93,507,906 | + | + | + | + | + | + | |
| 21 | Chlorogenic acid (3-O-Caffeoylquinic acid) | C16H18O9 | 35,510,291 | + | + | + | – | – | – | |
| 22 | Caffeic acid | C9H8O4 | 17,903,444 | + | + | + | + | + | + | |
| 23 | Cornusiin B or isomer | C48H30O30 | 108,507,437 | + | + | + | + | + | – | |
| 24 | Ampelopsin (Dihydromyricetin) | C15H12O8 | 31,904,540 | + | + | + | + | + | + | |
| 25 | Tellimagrandin I or isomer | C34H26O22 | 78,508,375 | + | + | + | + | + | + | |
| 26 | Coumaroylquinic acid | C16H18O8 | 33,709,235 | + | + | + | – | – | – | |
| 27 | Corilagin or isomer | C27H22O18 | 63,307,279 | + | + | + | – | – | – | |
| 28 | Caffeoylshikimic acid | C16H16O8 | 33,507,670 | – | + | + | – | – | – | |
| 29 | Procyanidin B isomer 3 | C30H26O12 | 57,713,460 | – | – | – | + | + | – | |
| 30 | Digalloylhexose | C20H20O14 | 48,307,749 | – | – | – | + | + | + | |
| 311 | Epigallocatechin-3-O-gallate (Teatannin II) | C22H18O11 | 45,707,709 | – | + | + | + | + | + | |
| 321 | Epicatechin | C15H14O6 | 28,907,121 | – | + | + | + | + | + | |
| 33 | Punicacortein C or D | C48H28O30 | 108,305,872 | – | – | – | + | + | + | |
| 34 | Trigalloylhexose isomer 1 | C27H24O18 | 63,508,844 | – | – | – | + | + | + | |
| 35 | Trigalloylhexose isomer 2 | C27H24O18 | 63,508,844 | – | – | – | + | + | + | |
| 36 | Di-O-methylcoruleoellagic acid | C16H10O10 | 36,101,958 | – | – | – | + | – | – | |
| 37 | Mangiferin | C19H18O11 | 42,107,709 | – | – | – | + | + | + | |
| 38 | Trigalloylhexose isomer 3 | C27H24O18 | 63,508,844 | – | – | – | + | + | + | |
| 391 | Taxifolin (Dihydroquercetin) | C15H12O7 | 30,305,048 | + | + | + | + | + | + | |
| 40 | Ferulic acid | C10H10O4 | 19,305,009 | + | + | + | – | – | – | |
| 41 | Tetragalloylhexose | C34H28O22 | 78,709,940 | – | – | – | + | + | – | |
| 421 | Epicatechin-3-O-gallate | C22H18O10 | 44,108,218 | + | + | + | + | + | + | |
| 43 | Chebulagic acid | C41H30O27 | 95,308,963 | + | + | + | – | – | – | |
| 44 | Ellagic acid O-glucuronide | C20H14O14 | 47,703,054 | – | – | – | + | + | + | |
| 45 | Ellagic acid O-hexoside isomer 1 | C20H16O13 | 46,305,127 | + | + | + | + | + | + | |
| 46 | Ellagic acid O-hexoside isomer 2 | C20H16O13 | 46,305,127 | + | + | + | + | + | + | |
| 47 | Coumaroylshikimic acid | C16H16O7 | 31,908,178 | + | + | + | – | – | – | |
| 48 | O–Methylellagic acid O-hexoside isomer 1 | C21H18O13 | 47,706,692 | – | – | – | + | + | + | |
| 49 | Ellagic acid C-hexoside isomer 1 | C20H16O13 | 46,506,692 | – | – | – | + | + | + | |
| 50 | Myricetin-O-hexoside | C21H20O13 | 47,908,257 | + | + | + | + | + | + | |
| 51 | Quercetin-O-galloylhexoside | C28H24O16 | 61,509,862 | + | + | + | – | – | – | |
| 52 | Vitexin (Apigenin-8-C-glucoside) | C21H20O10 | 43,311,348 | – | – | – | + | + | + | |
| 53 | Pentagalloylhexose | C41H32O26 | 93,911,036 | – | – | – | + | + | – | |
| 54 | Di-O-methylflavellagic acid O-hexoside | C22H20O14 | 50,707,749 | – | – | – | + | + | + | |
| 55 | Theaflavin or isomer | C29H24O12 | 56,513,461 | – | + | – | – | + | – | |
| 56 | O-Methylellagic acid O-hexoside isomer 2 | C21H18O13 | 47,706,692 | – | – | – | + | + | + | |
| 57 | Aromadendrin (Dihydrokaempferol) | C15H12O6 | 28,705,557 | + | + | + | + | + | – | |
| 58 | 3,3′-Di-O-methylellagic acid-4-O-glucoside | C22H20O13 | 49,108,257 | + | + | + | + | + | + | |
| 59 | Quercetin-3-O-glucuronide | C21H18O13 | 47,706,692 | + | + | + | – | – | – | |
| 60 | Isoquercitrin (Hirsutrin, Quercetin-3-O-glucoside) | C21H20O12 | 46,308,765 | + | + | + | – | – | – | |
| 61 | Rutin (Quercetin-3-O-rutinoside) | C27H30O16 | 61,116,122 | + | + | + | – | – | – | |
| 62 | Luteolin-7-O-glucoside (Cynaroside) | C21H20O11 | 44,709,274 | – | – | – | – | – | + | |
| 63 | Luteolin-O-deoxyhexosylhexoside | C27H30O15 | 59,315,065 | – | – | – | – | – | + | |
| 64 | Isovitexin (Apigenin-6-C-glucoside) | C21H20O10 | 43,311,348 | – | – | – | + | + | + | |
| 65 | Coatline A isomer | C21H24O10 | 43,512,913 | – | + | + | + | + | – | |
| 66 | Ellagic acid O-pentoside | C19H14O12 | 43,304,071 | – | – | – | + | + | + | |
| 67 | Ellagic acid C-hexoside isomer 2 | C20H16O13 | 46,506,692 | – | – | – | + | + | + | |
| 68 | Eschweilenol C (Ellagic acid-4-O-rhamnoside) | C20H16O12 | 44,705,636 | + | + | + | + | + | + | |
| 69 | Reinutrin (Quercetin-3-O-xyloside) | C20H18O11 | 43,307,709 | + | + | + | – | – | – | |
| 70 | Ellagic acid | C14H6O8 | 30,099,845 | + | + | + | + | + | + | |
| 71 | Avicularin (Quercetin-3-O-arabinoside) | C20H18O11 | 43,307,709 | + | + | + | – | – | – | |
| 721 | Myricetin (3,3′,4′,5,5′,7-Hexahydroxyflavone) | C15H10O8 | 31,702,974 | + | + | + | + | + | + | |
| 73 | Guaijaverin (Quercetin-3-O-arabinoside) | C20H18O11 | 43,307,709 | + | + | + | – | – | – | |
| 74 | Isorhamnetin-O-glucuronide isomer 1 | C22H20O13 | 49,108,257 | – | + | – | – | – | – | |
| 75 | Quercitrin (Quercetin-3-O-rhamnoside) | C21H20O11 | 44,709,274 | + | + | + | – | – | – | |
| 76 | Di-O-methylflavellagic acid O-pentoside | C21H18O13 | 47,706,692 | – | – | – | + | – | – | |
| 77 | Ducheside A (3-O-Methylellagic acid-4′-O-xyloside) | C20H16O12 | 44,705,636 | – | – | – | + | + | + | |
| 78 | Eriodictyol | C15H12O6 | 28,705,557 | + | + | – | + | + | + | |
| 79 | Isorhamnetin-3-O-glucoside | C22H22O12 | 47,710,330 | + | + | + | – | – | – | |
| 80 | Dimethoxy-tetrahydroxy(iso)flavone-O-hexoside | C23H24O13 | 50,711,387 | – | + | + | – | – | – | |
| 81 | Isorhamnetin-O-glucuronide isomer 2 | C22H20O13 | 49,108,257 | – | + | – | – | – | – | |
| 82 | Isorhamnetin-O-glucuronide | C22H20O13 | 49,108,257 | – | – | + | – | – | – | |
| 83 | Isorhamnetin-3-O-rutinoside (Narcissin) | C28H32O16 | 62,316,122 | + | + | + | – | – | – | |
| 84 | 4-Methoxycinnamic acid | C10H10O3 | 17,907,082 | – | – | – | – | + | – | |
| 85 | Di-O-methylellagic acid-O-pentoside | C21H18O12 | 46,107,200 | + | + | + | + | + | + | |
| 86 | 3,3′,4-Tri-O-methylflavellagic acid-4-O-glucoside | C23H22O14 | 52,109,314 | + | + | + | + | + | + | |
| 87 | Quercetin | C15H10O7 | 30,103,483 | + | + | + | – | – | – | |
| 88 | 3-O-Methylellagic acid | C15H8O8 | 31,501,410 | – | + | + | + | + | + | |
| 89 | Di-O-methylellagic acid-O-deoxyhexoside | C22H20O12 | 47,508,766 | – | – | – | + | + | + | |
| 901 | Naringenin | C15H12O5 | 27,106,065 | + | + | + | + | + | + | |
| 91 | Di-O-methylflavellagic acid isomer 1 | C16H10O9 | 34,502,466 | – | – | – | + | + | + | |
| 921 | Luteolin (3′,4′,5,7-Tetrahydroxyflavone) | C15H10O6 | 28,503,991 | + | + | + | – | + | – | |
| 93 | 3,3′-Di-O-methylellagic acid | C16H10O8 | 32,902,975 | + | + | + | + | + | + | |
| 94 | Kaempferol (3,4′,5,7-Tetrahydroxyflavone) | C15H10O6 | 28,705,556 | + | + | + | – | – | – | |
| 95 | Isorhamnetin (3′-Methoxy-3,4′,5,7-tetrahydroxyflavone) | C16H12O7 | 31,505,048 | + | + | + | – | – | – | |
| 96 | Dimethoxy-trihydroxy(iso)flavone | C17H14O7 | 32,906,613 | + | + | – | – | – | – | |
| 97 | Di-O-methylflavellagic acid isomer 2 | C16H10O9 | 34,502,466 | – | – | – | + | + | + | |
| 98 | Tetra-O-methylflavellagic acid isomer 1 | C18H14O9 | 37,305,596 | – | – | – | + | + | + | |
| 99 | Apigenin (4′,5,7-Trihydroxyflavone) | C15H10O5 | 26,904,500 | – | + | – | – | – | + | |
| 100 | 3,3′,4-Tri-O-methylellagic acid | C17H12O8 | 34,304,540 | + | + | + | + | + | + | |
| 101 | Undecanedioic acid | C11H20O4 | 21,512,834 | + | + | + | – | – | – | |
| 102 | Tetra-O-methylflavellagic acid isomer 2 | C18H14O9 | 37,305,596 | – | – | – | + | + | + | |
| 103 | 3,3′,4-Tri-O-methylflavellagic acid | C17H12O9 | 35,904,031 | + | + | + | + | + | + | |
| 104 | Dihydroxy-dimethoxy(iso)flavone | C17H14O6 | 31,307,122 | + | + | – | – | – | – | |
| 105 | Pinocembrin (5,7-Dihydroxyflavanone) | C15H12O4 | 25,506,573 | + | + | – | – | + | – | |
| 106 | Dihydroxy-trimethoxy(iso)flavone | C18H16O7 | 34,308,178 | + | + | + | – | – | – | |
| 107 | Dodecanedioic acid | C12H22O4 | 22,914,399 | + | + | + | – | – | – | |
| 108 | Hexadecanedioic acid | C16H30O4 | 28,520,659 | + | + | – | – | – | – |
| Samples | DPPH (mmol TE/g) | ABTS (mmol TE/g) | CUPRAC (mmol TE/g) | FRAP (mmol TE/g) | Metal Chelating (mg EDTAE/g) | Phosphomolybdenum (mmol TE/g) |
|---|---|---|---|---|---|---|
| Leaves-EA | 30 ± 0.01f | 0.26 ± 0.02e | 0.50 ± 0.04e | 0.26 ± 0.03d | 10.0 ± 0.8f | 2.0 ± 0.1e |
| Leaves-MeOH | 5.0 ± 0.1d | 3.0 ± 0.4c | 7.0 ± 0.2c | 4.0 ± 0.1c | 47.0 ± 0.8c | 4.0 ± 0.1d |
| Leaves-Water | 5.0 ± 0.1c | 4.0 ± 0.3b | 7.0 ± 0.2b | 6.0 ± 0.4b | 79.0 ± 0.9a | 4.0 ± 0.1d |
| Stem barks-EA | 3.0 ± 0.1e | 2.0 ± 0.1d | 5.0 ± 0.2d | 4.0 ± 0.1c | 30.0 ± 0.5e | 4.0 ± 0.1c |
| Stem barks-MeOH | 6.0 ± 0.1a | 5.0 ± 0.1a | 8.0 ± 0.2a | 6.0 ± 0.3a | 45.0 ± 0.6d | 6.0 ± 0.2a |
| Stem barks-Water | 5.0 ± 0.1b | 4.0 ± 0.4b | 8.0 ± 0.1b | 6.0 ± 0.2a | 61.0 ± 0.4b | 5.0 ± 0.1a |
| Samples | AChE Inhibition (mg GALAE/g) | BChE Inhibition (mg GALAE/g) | Tyrosinase Inhibition (mg KAE/g) | Amylase Inhibition (mmol ACAE/g) | Glucosidase Inhibition (mmol ACAE/g) |
|---|---|---|---|---|---|
| Leaves-EA | 4.0 ± 0.2d | 3.0 ± 0.2b | 131.0 ± 0.2d | 0.79 ± 0.04b | 15.0±0.1b |
| Leaves-MeOH | 4.68 ± 0.02a | 4.0 ± 0.1a | 154.0 ± 0.2b | 1.0 ± 0.1a | nd |
| Leaves-Water | 4.19 ± 0.04c | nd | 113 ± 1e | 0.53 ± 0.04c | nd |
| Stem barks-EA | 4.0 ± 0.1bc | 2.0 ± 0.2c | 152.0 ± 0.6c | 1.0 ± 0.1b | 15.0 ± 0.1a |
| Stem barks-MeOH | 4.0 ± 0.1b | 1.0 ± 0.3c | 155.26 ± 0.04a | 0.85 ± 0.03b | nd |
| Stem barks-Water | 4.0 ± 0.1e | 0.5 ± 0.1d | 113.0 ± 0.6e | 0.19 ± 0.01d | nd |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orlando, G.; Ferrante, C.; Zengin, G.; Sinan, K.I.; Bene, K.; Diuzheva, A.; Jekő, J.; Cziáky, Z.; Di Simone, S.; Recinella, L.; et al. Qualitative Chemical Characterization and Multidirectional Biological Investigation of Leaves and Bark Extracts of Anogeissus leiocarpus (DC.) Guill. & Perr. (Combretaceae). Antioxidants 2019, 8, 343. https://doi.org/10.3390/antiox8090343
Orlando G, Ferrante C, Zengin G, Sinan KI, Bene K, Diuzheva A, Jekő J, Cziáky Z, Di Simone S, Recinella L, et al. Qualitative Chemical Characterization and Multidirectional Biological Investigation of Leaves and Bark Extracts of Anogeissus leiocarpus (DC.) Guill. & Perr. (Combretaceae). Antioxidants. 2019; 8(9):343. https://doi.org/10.3390/antiox8090343
Chicago/Turabian StyleOrlando, Giustino, Claudio Ferrante, Gokhan Zengin, Kouadio Ibrahime Sinan, Kouadio Bene, Alina Diuzheva, József Jekő, Zoltán Cziáky, Simonetta Di Simone, Lucia Recinella, and et al. 2019. "Qualitative Chemical Characterization and Multidirectional Biological Investigation of Leaves and Bark Extracts of Anogeissus leiocarpus (DC.) Guill. & Perr. (Combretaceae)" Antioxidants 8, no. 9: 343. https://doi.org/10.3390/antiox8090343
APA StyleOrlando, G., Ferrante, C., Zengin, G., Sinan, K. I., Bene, K., Diuzheva, A., Jekő, J., Cziáky, Z., Di Simone, S., Recinella, L., Chiavaroli, A., Leone, S., Brunetti, L., Picot-Allain, C. M. N., Mahomoodally, M. F., & Menghini, L. (2019). Qualitative Chemical Characterization and Multidirectional Biological Investigation of Leaves and Bark Extracts of Anogeissus leiocarpus (DC.) Guill. & Perr. (Combretaceae). Antioxidants, 8(9), 343. https://doi.org/10.3390/antiox8090343