Crosstalk Between Adipokines and Paraoxonase 1: A New Potential Axis Linking Oxidative Stress and Inflammation

 ,
,  ,
,  ,
,  ,
,  and
and
Abstract
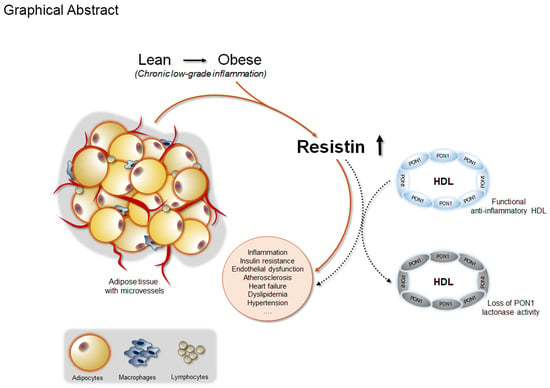
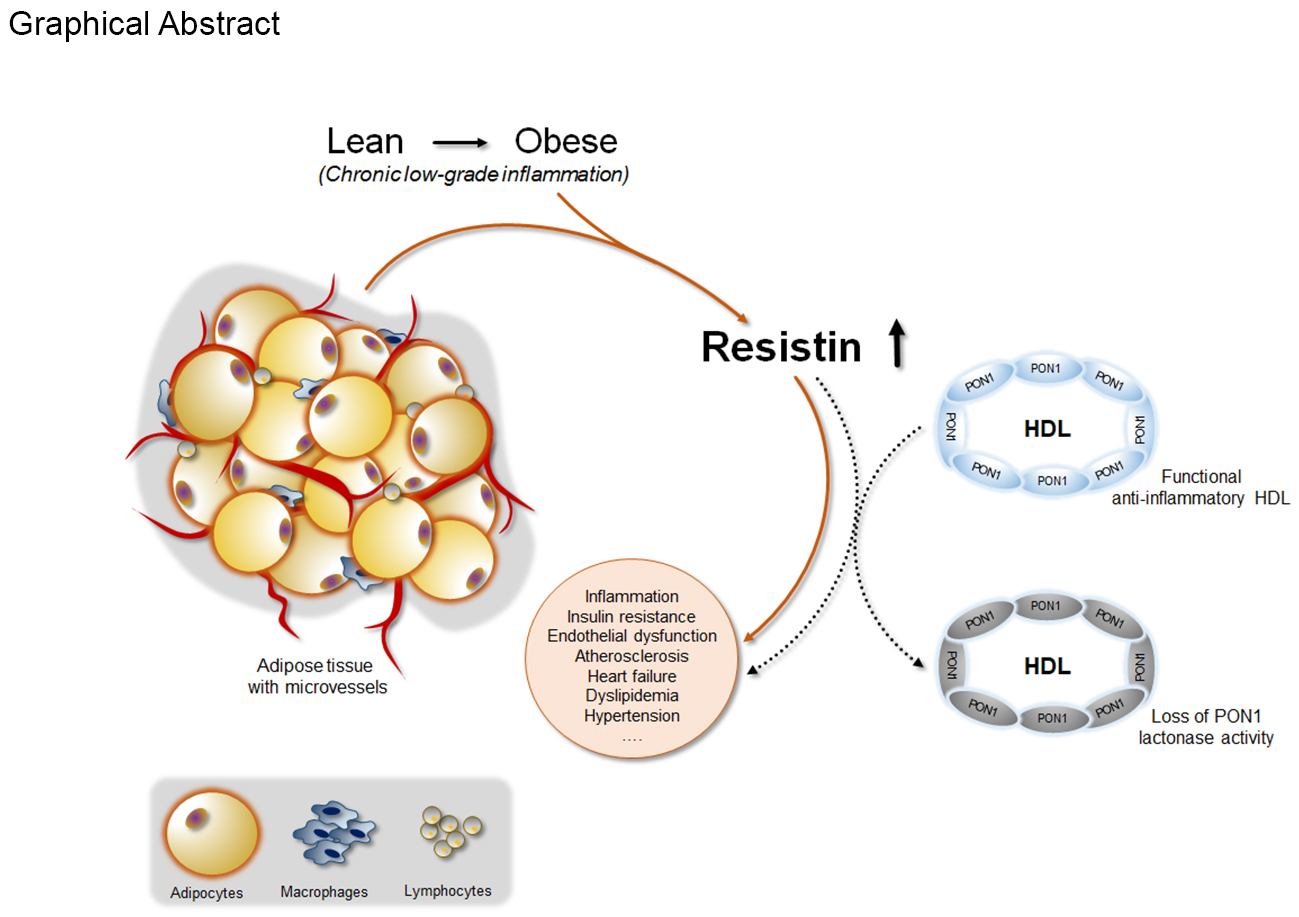
1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Biochemical Assays
2.3. Adipokines Assessment
2.4. PON1 Activities
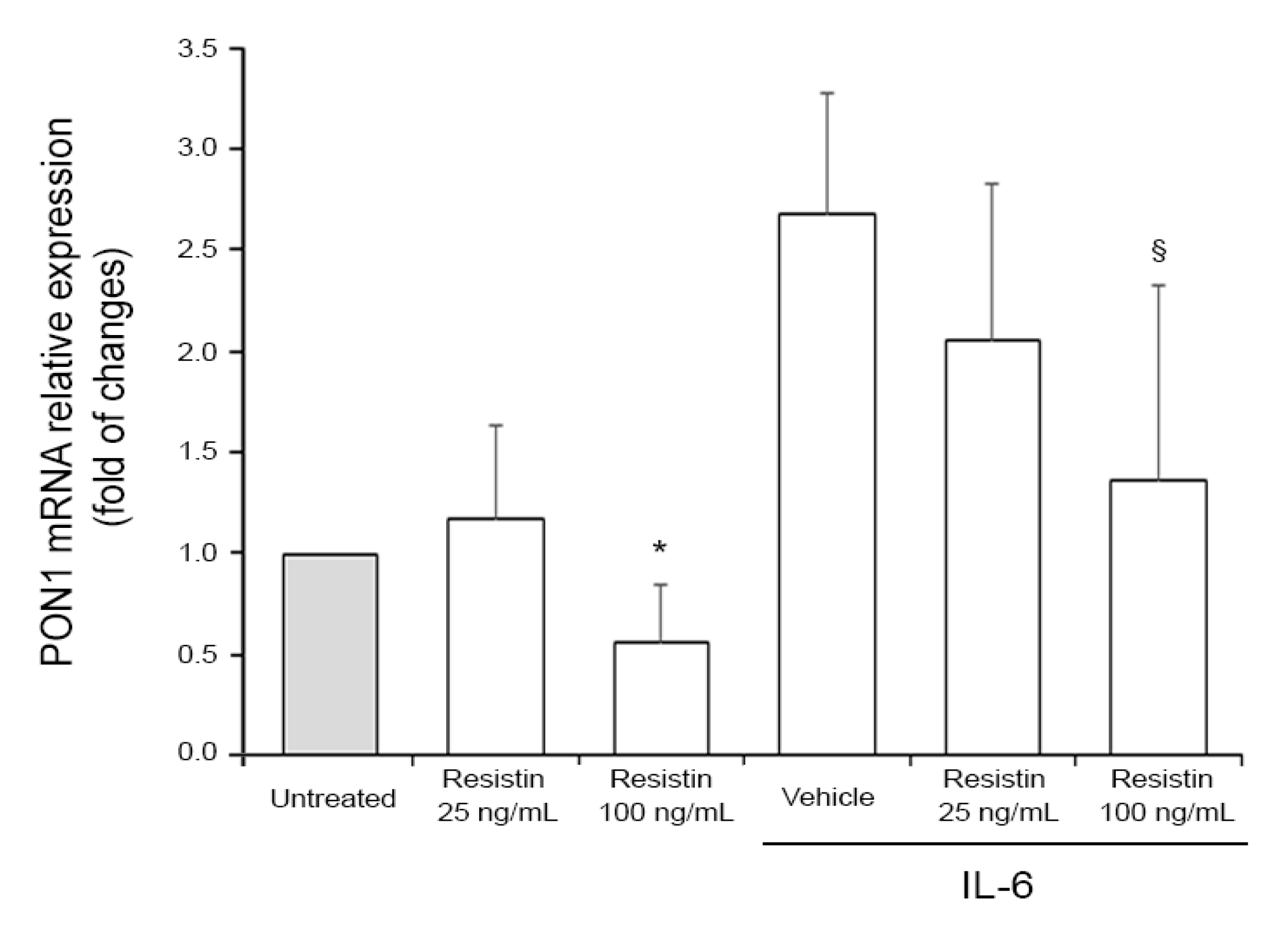
2.5. Cell Treatments and RT-PCR Analysis
2.6. Statistical Analysis
3. Results
Associations Between PON1 Activities and Adipokines: Study Population and In Vitro Assessment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Furlong, C.E.; Marsillach, J.; Jarvik, G.P.; Costa, L.G. Paraoxonases-1, -2 and -3: What are their functions? Chem. Biol. Interact. 2016, 259, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Cervellati, C.; Valacchi, G.; Tisato, V.; Zuliani, G.; Marsillach, J. Evaluating the link between Paraoxonase-1 levels and Alzheimer’s disease development. Minerva Med. 2019, 110, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Levy, D.; Reichert, C.O.; Bydlowski, S.P. Paraoxonases Activities and Polymorphisms in Elderly and Old-Age Diseases: An Overview. Antioxidants 2019, 8, 118. [Google Scholar] [CrossRef] [PubMed]
- Mackness, M.; Mackness, B. Human paraoxonase-1 (PON1): Gene structure and expression, promiscuous activities and multiple physiological roles. Gene 2015, 567, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Mineo, C.; Shaul, P.W. PON-dering differences in HDL function in coronary artery disease. J. Clin. Invest. 2011, 121, 2545–2548. [Google Scholar] [CrossRef] [PubMed]
- Cervellati, C.; Vigna, G.B.; Trentini, A.; Sanz, J.M.; Zimetti, F.; Dalla Nora, E.; Morieri, M.L.; Zuliani, G.; Passaro, A. Paraoxonase-1 activities in individuals with different HDL circulating levels: Implication in reverse cholesterol transport and early vascular damage. Atherosclerosis 2019, 285, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Camps, J.; Marsillach, J.; Joven, J. The paraoxonases: Role in human diseases and methodological difficulties in measurement. Crit. Rev. Clin. Lab. Sci. 2009, 46, 83–106. [Google Scholar] [CrossRef] [PubMed]
- Cervellati, C.; Trentini, A.; Romani, A.; Bellini, T.; Bosi, C.; Ortolani, B.; Zurlo, A.; Passaro, A.; Seripa, D.; Zuliani, G. Serum paraoxonase and arylesterase activities of paraoxonase-1 (PON-1), mild cognitive impairment, and 2-year conversion to dementia: A pilot study. J. Neurochem. 2015, 135, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, S.A.; Connell, J.M.C. The link between abdominal obesity, metabolic syndrome and cardiovascular disease. Nutr. Metab. Cardiovasc. Dis. 2007, 17, 319–326. [Google Scholar] [CrossRef]
- Keaney, J.F.; Larson, M.G.; Vasan, R.S.; Wilson, P.W.F.; Lipinska, I.; Corey, D.; Massaro, J.M.; Sutherland, P.; Vita, J.A.; Benjamin, E.J. Obesity and systemic oxidative stress: Clinical correlates of oxidative stress in the Framingham study. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 434–439. [Google Scholar] [CrossRef]
- Valacchi, G.; Virgili, F.; Cervellati, C.; Pecorelli, A. OxInflammation: From Subclinical Condition to Pathological Biomarker. Front. Physiol. 2018, 9, 858. [Google Scholar] [CrossRef] [PubMed]
- Thomàs-Moyà, E.; Gómez-Pérez, Y.; Fiol, M.; Gianotti, M.; Lladó, I.; Proenza, A.M. Gender related differences in paraoxonase 1 response to high-fat diet-induced oxidative stress. Obesity (Silver Spring) 2008, 16, 2232–2238. [Google Scholar] [CrossRef] [PubMed]
- Woudberg, N.J.; Goedecke, J.H.; Blackhurst, D.; Frias, M.; James, R.; Opie, L.H.; Lecour, S. Association between ethnicity and obesity with high-density lipoprotein (HDL) function and subclass distribution. Lipids Health Dis. 2016, 15, 92. [Google Scholar] [CrossRef] [PubMed]
- Cervellati, C.; Bonaccorsi, G.; Trentini, A.; Valacchi, G.; Sanz, J.M.; Squerzanti, M.; Spagnolo, M.; Massari, L.; Crivellari, I.; Greco, P.; et al. Paraoxonase, arylesterase and lactonase activities of paraoxonase-1 (PON1) in obese and severely obese women. Scand. J. Clin. Lab. Invest. 2018, 78, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Huen, K.; Harley, K.; Beckman, K.; Eskenazi, B.; Holland, N. Associations of PON1 and genetic ancestry with obesity in early childhood. PLoS ONE 2013, 8, e62565. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, T.; Nicholls, S.J.; Topol, E.J.; Zhang, R.; Yang, X.; Schmitt, D.; Fu, X.; Shao, M.; Brennan, D.M.; Ellis, S.G.; et al. Relationship of paraoxonase 1 (PON1) gene polymorphisms and functional activity with systemic oxidative stress and cardiovascular risk. JAMA 2008, 299, 1265–1276. [Google Scholar] [CrossRef]
- Van Himbergen, T.M.; van der Schouw, Y.T.; Voorbij, H.A.M.; van Tits, L.J.H.; Stalenhoef, A.F.H.; Peeters, P.H.M.; Roest, M. Paraoxonase (PON1) and the risk for coronary heart disease and myocardial infarction in a general population of Dutch women. Atherosclerosis 2008, 199, 408–414. [Google Scholar] [CrossRef]
- Costa, L.G.; Vitalone, A.; Cole, T.B.; Furlong, C.E. Modulation of paraoxonase (PON1) activity. Biochem. Pharmacol. 2005, 69, 541–550. [Google Scholar] [CrossRef]
- Kwon, H.; Pessin, J.E. Adipokines mediate inflammation and insulin resistance. Front. Endocrinol. (Lausanne) 2013, 4, 71. [Google Scholar] [CrossRef]
- Fain, J.N.; Madan, A.K.; Hiler, M.L.; Cheema, P.; Bahouth, S.W. Comparison of the release of adipokines by adipose tissue, adipose tissue matrix, and adipocytes from visceral and subcutaneous abdominal adipose tissues of obese humans. Endocrinology 2004, 145, 2273–2282. [Google Scholar] [CrossRef]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Cervellati, C.; Bonaccorsi, G.; Bergamini, C.M.; Fila, E.; Greco, P.; Valacchi, G.; Massari, L.; Gonelli, A.; Tisato, V. Association between circulatory levels of adipokines and bone mineral density in postmenopausal women. Menopause 2016, 23, 984–992. [Google Scholar] [CrossRef] [PubMed]
- Cervellati, C.; Pansini, F.S.; Bonaccorsi, G.; Pascale, G.; Bagni, B.; Castaldini, C.; Ferrazini, S.; Ridolfi, F.; Pedriali, M.; Guariento, A.; et al. Body mass index is a major determinant of abdominal fat accumulation in pre-, peri- and post-menopausal women. Gynecol. Endocrinol. 2009, 25, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Ley, C.J.; Lees, B.; Stevenson, J.C. Sex- and menopause-associated changes in body-fat distribution. Am. J. Clin. Nutr. 1992, 55, 950–954. [Google Scholar] [CrossRef] [PubMed]
- Cervellati, C.; Pansini, F.S.; Bonaccorsi, G.; Bergamini, C.M.; Patella, A.; Casali, F.; Fantini, G.F.; Pascale, G.; Castaldini, C.; Ferrazzini, S.; et al. 17β-estradiol levels and oxidative balance in a population of pre-, peri-, and post-menopausal women. Gynecol. Endocrinol. 2011, 27, 1028–1032. [Google Scholar] [CrossRef] [PubMed]
- Tisato, V.; Zauli, G.; Gianesini, S.; Menegatti, E.; Brunelli, L.; Manfredini, R.; Zamboni, P.; Secchiero, P. Modulation of Circulating Cytokine-Chemokine Profile in Patients Affected by Chronic Venous Insufficiency Undergoing Surgical Hemodynamic Correction. J. Immunol. Res. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Hayek, J.; Cervellati, C.; Crivellari, I.; Pecorelli, A.; Valacchi, G. Lactonase Activity and Lipoprotein-Phospholipase A2as Possible Novel Serum Biomarkers for the Differential Diagnosis of Autism Spectrum Disorders and Rett Syndrome: Results from a Pilot Study. Oxid. Med. Cell. Longev. 2017, 2017, 5694058. [Google Scholar] [CrossRef] [PubMed]
- Zauli, G.; Tisato, V.; Melloni, E.; Volpato, S.; Cervellati, C.; Bonaccorsi, G.; Radillo, O.; Marci, R.; Secchiero, P. Inverse correlation between circulating levels of TNF-related apoptosis-inducing ligand and 17β-estradiol. J. Clin. Endocrinol. Metab. 2014, 99, 659–664. [Google Scholar] [CrossRef] [PubMed]
- Ponce-Ruiz, N.; Murillo-González, F.E.; Rojas-García, A.E.; Mackness, M.; Bernal-Hernández, Y.Y.; Barrón-Vivanco, B.S.; González-Arias, C.A.; Medina-Díaz, I.M. Transcriptional regulation of human Paraoxonase 1 by nuclear receptors. Chem. Biol. Interact. 2017, 268, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Kumon, Y.; Suehiro, T.; Ikeda, Y.; Hashimoto, K. Human paraoxonase-1 gene expression by HepG2 cells is downregulated by interleukin-1β and tumor necrosis factor-α, but is upregulated by interleukin-6. Life Sci. 2003, 73, 2807–2815. [Google Scholar] [CrossRef]
- Cheng, C.-C.; Hsueh, C.-M.; Chen, C.-Y.; Chen, T.-H.; Hsu, S.-L. Interleukin-6 upregulates paraoxonase 1 gene expression via an AKT/NF-κB-dependent pathway. Biochem. Biophys. Res. Commun. 2013, 437, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Pecorelli, A.; Cervellati, C.; Hayek, J.; Valacchi, G. OxInflammation in Rett syndrome. Int. J. Biochem. Cell Biol. 2016, 81, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Mackness, B.; Hine, D.; Liu, Y.; Mastorikou, M.; Mackness, M. Paraoxonase-1 inhibits oxidised LDL-induced MCP-1 production by endothelial cells. Biochem. Biophys. Res. Commun. 2004, 318, 680–683. [Google Scholar] [CrossRef] [PubMed]
- Castellazzi, M.; Trentini, A.; Romani, A.; Valacchi, G.; Bellini, T.; Bonaccorsi, G.; Fainardi, E.; Cavicchio, C.; Passaro, A.; Zuliani, G.; et al. Decreased arylesterase activity of paraoxonase-1 (PON-1) might be a common denominator of neuroinflammatory and neurodegenerative diseases. Int. J. Biochem. Cell Biol. 2016, 81, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Patra, S.K.; Singh, K.; Singh, R. Paraoxonase 1: A better atherosclerotic risk predictor than HDL in type 2 diabetes mellitus. Diabetes Metab. Syndr. 2013, 7, 108–111. [Google Scholar] [CrossRef] [PubMed]
- Huen, K.; Richter, R.; Furlong, C.; Eskenazi, B.; Holland, N. Validation of PON1 enzyme activity assays for longitudinal studies. Clin. Chim. Acta 2009, 402, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Passaro, A.; Vigna, G.B.; Romani, A.; Sanz, J.M.; Cavicchio, C.; Bonaccorsi, G.; Valacchi, G.; Cervellati, C. Distribution of Paraoxonase-1 (PON-1) and Lipoprotein Phospholipase A2 (Lp-PLA2) across Lipoprotein Subclasses in Subjects with Type 2 Diabetes. Oxid. Med. Cell. Longev. 2018, 2018, 1752940. [Google Scholar] [CrossRef] [PubMed]
- Aharoni, S.; Aviram, M.; Fuhrman, B. Paraoxonase 1 (PON1) reduces macrophage inflammatory responses. Atherosclerosis 2013, 228, 353–361. [Google Scholar] [CrossRef]
- Fülöp, P.; Harangi, M.; Seres, I.; Paragh, G. Paraoxonase-1 and adipokines: Potential links between obesity and atherosclerosis. Chem. Biol. Interact. 2016, 259, 388–393. [Google Scholar] [CrossRef]
- Beltowski, J.; Jamroz-Wiśniewska, A.; Borkowska, E.; Wójcicka, G. Differential effect of antioxidant treatment on plasma and tissue paraoxonase activity in hyperleptinemic rats. Pharmacol. Res. 2005, 51, 523–532. [Google Scholar] [CrossRef]
- Varga, Z.; Paragh, G.; Seres, I.; Kakuk, G.; Karanyi, Z.; Karpati, I.; Matyus, J.; Csongradi, E.; Juhasz, A.; Balla, J.; et al. Hyperleptinemia Is Not Responsible for Decreased Paraoxonase Activity in Hemodialysis Patients. Nephron Clin. Pract. 2006, 103, c114–c120. [Google Scholar] [CrossRef] [PubMed]
- Bajnok, L.; Seres, I.; Varga, Z.; Jeges, S.; Peti, A.; Karanyi, Z.; Juhasz, A.; Csongradi, E.; Mezosi, E.; Nagy, E.V.; et al. Relationship of endogenous hyperleptinemia to serum paraoxonase 1, cholesteryl ester transfer protein, and lecithin cholesterol acyltransferase in obese individuals. Metabolism 2007, 56, 1542–1549. [Google Scholar] [CrossRef] [PubMed]
- Bajnok, L.; Csongradi, E.; Seres, I.; Varga, Z.; Jeges, S.; Peti, A.; Karanyi, Z.; Juhasz, A.; Mezosi, E.; Nagy, E.V.; et al. Relationship of adiponectin to serum paraoxonase 1. Atherosclerosis 2008, 197, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Koncsos, P.; Seres, I.; Harangi, M.; Illyés, I.; Józsa, L.; Gönczi, F.; Bajnok, L.; Paragh, G. Human paraoxonase-1 activity in childhood obesity and its relation to leptin and adiponectin levels. Pediatr. Res. 2010, 67, 309–313. [Google Scholar] [CrossRef]
- Bajnok, L.; Seres, I.; Varga, Z.; Jeges, S.; Peti, A.; Karanyi, Z.; Juhasz, A.; Csongradi, E.; Mezosi, E.; Nagy, E.; et al. Relationship of Serum Resistin Level to Traits of Metabolic Syndrome and Serum Paraoxonase 1 Activity in a Population with a Broad Range of Body Mass Index. Exp. Clin. Endocrinol. Diabetes 2008, 116, 592–599. [Google Scholar] [CrossRef]
- Bednarska-Makaruk, M.; Graban, A.; Wiśniewska, A.; Łojkowska, W.; Bochyńska, A.; Gugała-Iwaniuk, M.; Sławińska, K.; Ługowska, A.; Ryglewicz, D.; Wehr, H. Association of adiponectin, leptin and resistin with inflammatory markers and obesity in dementia. Biogerontology 2017, 18, 561–580. [Google Scholar] [CrossRef]
- Fernández-Sánchez, A.; Madrigal-Santillán, E.; Bautista, M.; Esquivel-Soto, J.; Morales-González, Á.; Esquivel-Chirino, C.; Durante-Montiel, I.; Sánchez-Rivera, G.; Valadez-Vega, C.; Morales-González, J.A. Inflammation, oxidative stress, and obesity. Int. J. Mol. Sci. 2011, 12, 3117–3132. [Google Scholar] [CrossRef]
- Oh, K.W.; Lee, W.Y.; Rhee, E.J.; Baek, K.H.; Yoon, K.H.; Kang, M.I.; Yun, E.J.; Park, C.Y.; Ihm, S.H.; Choi, M.G.; et al. The relationship between serum resistin, leptin, adiponectin, ghrelin levels and bone mineral density in middle-aged men. Clin. Endocrinol. (Oxf) 2005, 63, 131–138. [Google Scholar] [CrossRef]
- Aggarwal, G.; Prajapati, R.; Tripathy, R.K.; Bajaj, P.; Iyengar, A.R.S.; Sangamwar, A.T.; Pande, A.H. Toward Understanding the Catalytic Mechanism of Human Paraoxonase 1: Site-Specific Mutagenesis at Position 192. PLoS ONE 2016, 11, e0147999. [Google Scholar] [CrossRef]
- Marsillach, J.; Camps, J.; Ferré, N.; Beltran, R.; Rull, A.; Mackness, B.; Mackness, M.; Joven, J. Paraoxonase-1 is related to inflammation, fibrosis and PPAR delta in experimental liver disease. BMC Gastroenterol. 2009, 9, 3. [Google Scholar] [CrossRef]
- Nguyen, S.D.; Hung, N.D.; Cheon-Ho, P.; Ree, K.M.; Dai-Eun, S. Oxidative inactivation of lactonase activity of purified human paraoxonase 1 (PON1). Biochim. Biophys. Acta 2009, 1790, 155–160. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Demographic and Clinical Parameters | Value |
|---|---|
| Number of subjects, n | 107 |
| Age, years | 57 ± 4 |
| Years since menopause, years | 4 (2–6) |
| Smoking, n (%) | 8 (7) |
| Hypertension, n (%) | 10 (9) |
| BMI, kg/m2 | 24 ± 3 |
| Waist circumference, cm | 82 ± 10 |
| Total cholesterol, mmol/L | 5.1 ± 1.3 |
| HDL-C, mmol/L | 1.8 ± 0.6 |
| Triglycerides, mmol/L | 0.9 ± 0.6 |
| LDL-C, mmol/L | 3.0 ± 1.3 |
| hs-CRP, mg/dL | 1.3 (0.5–3.0) |
| Adipokines | Arylesterase | Paraoxonase | Log10 Lactonase |
|---|---|---|---|
| IL-8 | 0.265 a | −0.014 | 0.086 |
| TNF-α | 0.001 | 0.090 | −0.024 |
| IL-6 | −0.120 | 0.034 | −0.039 |
| Resistin | 0.069 | −0.102 | −0.322 b |
| Adiponectin | 0.085 | −0.049 | −0.152 |
| Leptin | 0.017 | 0.164 | 0.070 |
| MCP-1 | 0.022 | 0.012 | −0.022 |
| HGF | −0.037 | 0.053 | −0.007 |
| Dependent Variable | Predictor | β (p Value) | βage/HDL-C adjusted (p Value) | βfully adjusted * (p Value) |
|---|---|---|---|---|
| Arylesterase | IL-8 | 0.265 a | 0.221 | 0.185 |
| Lactonase | Resistin | −0.326 b | −0.344 b | −0.346 b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tisato, V.; Romani, A.; Tavanti, E.; Melloni, E.; Milani, D.; Bonaccorsi, G.; Sanz, J.M.; Gemmati, D.; Passaro, A.; Cervellati, C. Crosstalk Between Adipokines and Paraoxonase 1: A New Potential Axis Linking Oxidative Stress and Inflammation. Antioxidants 2019, 8, 287. https://doi.org/10.3390/antiox8080287
Tisato V, Romani A, Tavanti E, Melloni E, Milani D, Bonaccorsi G, Sanz JM, Gemmati D, Passaro A, Cervellati C. Crosstalk Between Adipokines and Paraoxonase 1: A New Potential Axis Linking Oxidative Stress and Inflammation. Antioxidants. 2019; 8(8):287. https://doi.org/10.3390/antiox8080287
Chicago/Turabian StyleTisato, Veronica, Arianna Romani, Elisa Tavanti, Elisabetta Melloni, Daniela Milani, Gloria Bonaccorsi, Juana M. Sanz, Donato Gemmati, Angelina Passaro, and Carlo Cervellati. 2019. "Crosstalk Between Adipokines and Paraoxonase 1: A New Potential Axis Linking Oxidative Stress and Inflammation" Antioxidants 8, no. 8: 287. https://doi.org/10.3390/antiox8080287
APA StyleTisato, V., Romani, A., Tavanti, E., Melloni, E., Milani, D., Bonaccorsi, G., Sanz, J. M., Gemmati, D., Passaro, A., & Cervellati, C. (2019). Crosstalk Between Adipokines and Paraoxonase 1: A New Potential Axis Linking Oxidative Stress and Inflammation. Antioxidants, 8(8), 287. https://doi.org/10.3390/antiox8080287