Disruption of Selenium Handling During Puberty Causes Sex-Specific Neurological Impairments in Mice



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Neuromotor and Behavioral Tests
2.2.1. Rotarod Assay for Motor Coordination (Ataxia)
2.2.2. Open Field Assay for Locomotion
2.2.3. Audio Open Field Audiogenic Seizure Test
2.3. Histology and Immunohistochemistry
2.4. Silver Staining and Quantification
2.5. Protein Extraction and Glutathione Peroxidase (Gpx) Activity Assay
2.6. Statistical Analysis
3. Results
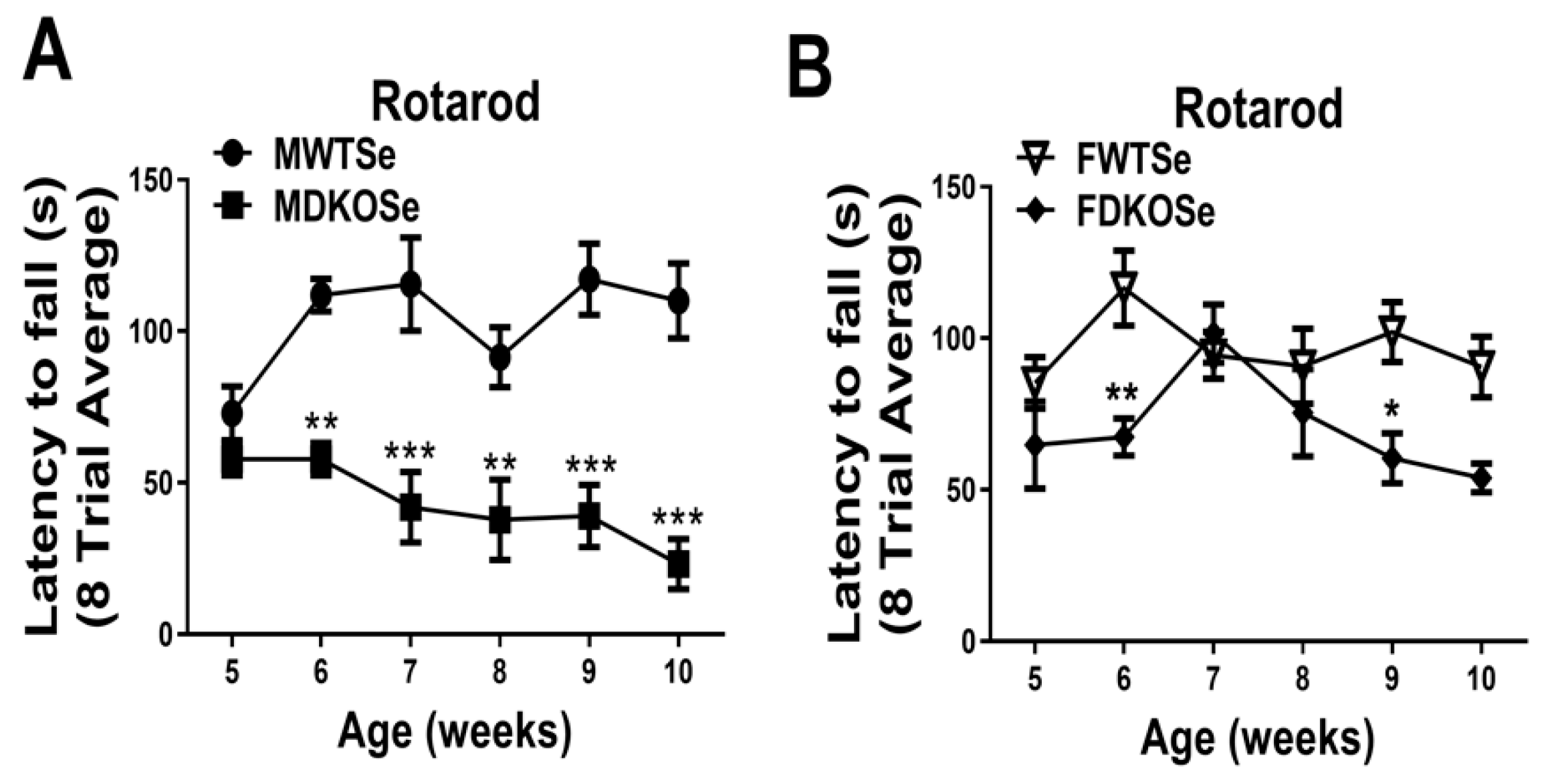
3.1. Male and Female DKOSe Mice Exhibited Impaired Motor Coordination Prior to 10 Weeks of Age
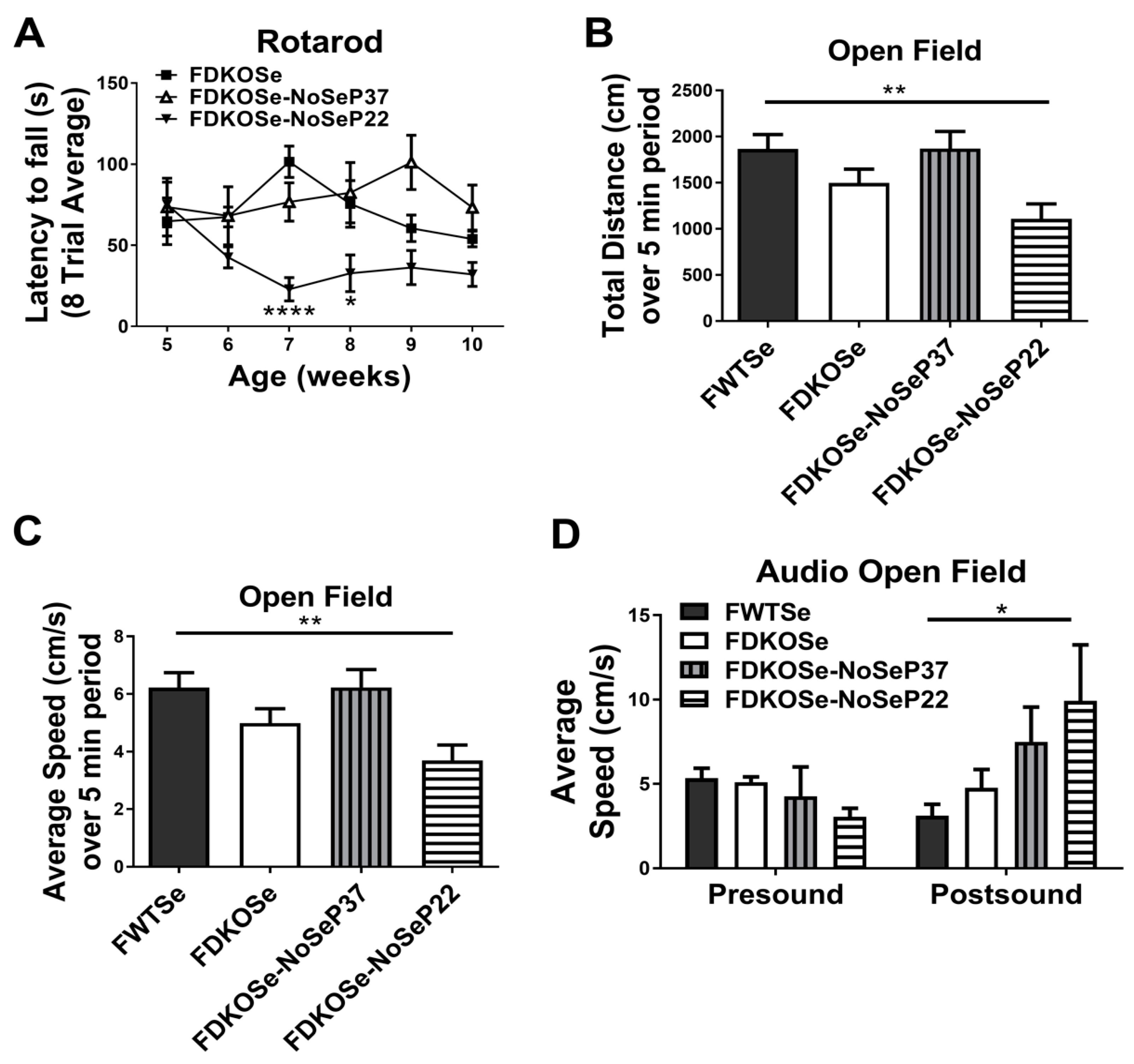
3.2. Neurological Dysfunction in FDKOSe Mice was Modulated by SeH2O Removal
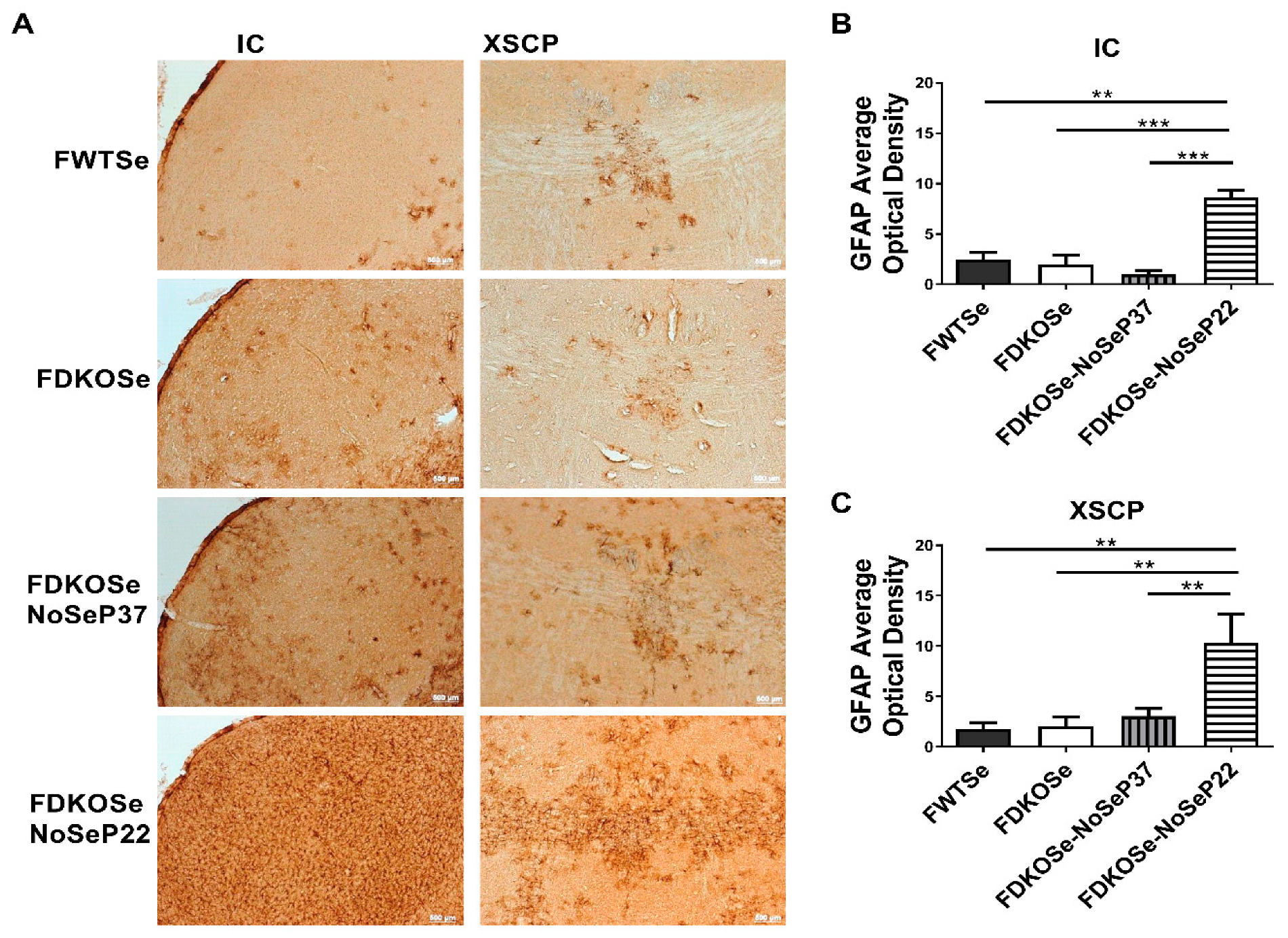
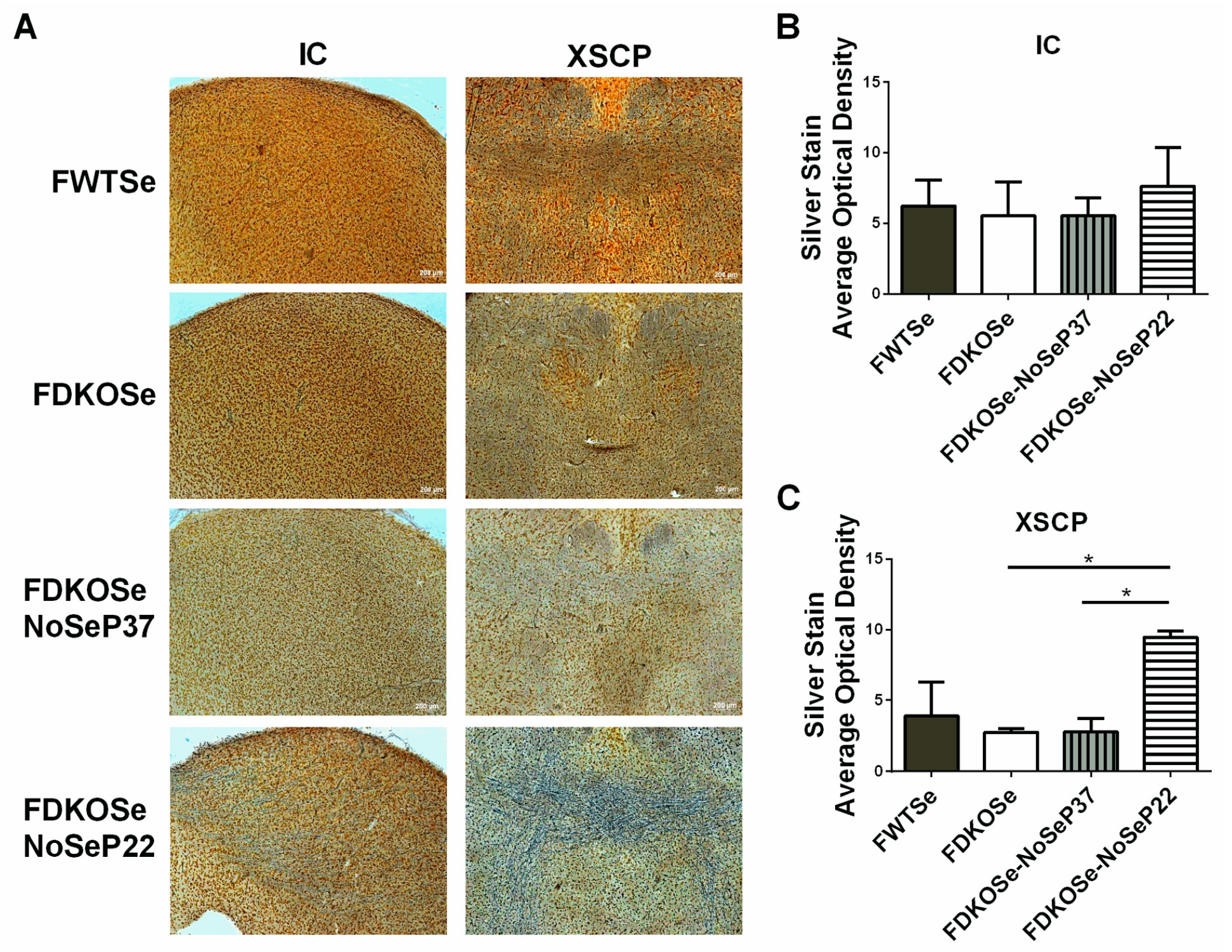
3.3. FDKOSe-NoSeP22 Mice Exhibited Neuroinflammation and Neurodegeneration in Auditory Brain Nuclei and Motor Tracts
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fairweather-Tait, S.J.; Bao, Y.; Broadley, M.R.; Collings, R.; Ford, D.; Hesketh, J.E.; Hurst, R. Selenium in human health and disease. Antioxid. Redox Signal. 2011, 14, 1337–1383. [Google Scholar] [CrossRef] [PubMed]
- Burk, R.F.; Hill, K.E. Selenoprotein P-expression, functions, and roles in mammals. Biochim. Biophys. Acta 2009, 1790, 1441–1447. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Berry, M.J. Selenium and selenoproteins in the brain and brain diseases. J. Neurochem. 2003, 86, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mihara, H.; Kurihara, T.; Watanabe, T.; Yoshimura, T.; Esaki, N. cDNA cloning, purification, and characterization of mouse liver selenocysteine lyase. Candidate for selenium delivery protein in selenoprotein synthesis. J. Biol. Chem. 2000, 275, 6195–6200. [Google Scholar] [CrossRef] [PubMed]
- Esaki, N.; Takeshi, N.; Tanaka, H.; Soda, K. Selenocysteine lyase, a novel enzyme that specifically acts on selenocysteine. J. Biol. Chem. 1982, 257, 4386–4391. [Google Scholar] [PubMed]
- Schomburg, L.; Schweizer, U.; Holtmann, B.; Flohe, L.; Sendtner, M.; Kohrle, J. Gene disruption discloses role of selenoprotein P in selenium delivery to target tissues. J. Biochem. 2003, 370, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Raman, A.V.; Pitts, M.W.; Seyedali, A.; Hashimoto, A.C.; Seale, L.A.; Bellinger, F.P.; Berry, M.J. Absence of selenoprotein P but not selenocysteine lyase results in severe neurological dysfunction. Genes Brain Behav. 2012, 11, 601–613. [Google Scholar] [CrossRef] [PubMed]
- Seale, L.A.; Hashimoto, A.C.; Kurokawa, S.; Gilman, C.L.; Seyedali, A.; Bellinger, F.P.; et al. Disruption of the selenocysteine lyase-mediated selenium recycling pathway leads to metabolic syndrome in mice. Mol. Cell. Biol. 2012, 32, 4141–4154. [Google Scholar] [CrossRef] [PubMed]
- Byrns, C.N.; Pitts, M.W.; Gilman, C.A.; Hashimoto, A.C.; Berry, M.J. Mice lacking selenoprotein P and selenocysteine lyase exhibit severe neurological dysfunction, neurodegeneration, and audiogenic seizures. J. Biol. Chem. 2014, 289, 9662–9674. [Google Scholar] [CrossRef] [PubMed]
- Pitts, M.W.; Kremer, P.M.; Hashimoto, A.C.; Torres, D.J.; Byrns, C.N.; Williams, C.S.; Berry, M.J. Competition between the brain and testes under selenium-compromised conditions: insight into sex differences in selenium metabolism and risk of neurodevelopmental disease. J. Neurosci. 2015, 35, 15326–15338. [Google Scholar] [CrossRef] [PubMed]
- Walls, A.B.; Eyjolfsson, E.M.; Smeland, O.B.; Nilsen, L.H.; Schousboe, I.; Schousboe, A.; Sonnewald, U.; Waagepetersen, H.S. Knockout of GAD65 has major impact on synaptic GABA synthesized from astrocyte-derived glutamine. J. Cereb. Blood Flow Metab. 2011, 31, 494–503. [Google Scholar] [CrossRef] [PubMed]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kremer, P.M.; Torres, D.J.; Hashimoto, A.C.; Berry, M.J. Disruption of Selenium Handling During Puberty Causes Sex-Specific Neurological Impairments in Mice. Antioxidants 2019, 8, 110. https://doi.org/10.3390/antiox8040110
Kremer PM, Torres DJ, Hashimoto AC, Berry MJ. Disruption of Selenium Handling During Puberty Causes Sex-Specific Neurological Impairments in Mice. Antioxidants. 2019; 8(4):110. https://doi.org/10.3390/antiox8040110
Chicago/Turabian StyleKremer, Penny M., Daniel J. Torres, Ann C. Hashimoto, and Marla J. Berry. 2019. "Disruption of Selenium Handling During Puberty Causes Sex-Specific Neurological Impairments in Mice" Antioxidants 8, no. 4: 110. https://doi.org/10.3390/antiox8040110
APA StyleKremer, P. M., Torres, D. J., Hashimoto, A. C., & Berry, M. J. (2019). Disruption of Selenium Handling During Puberty Causes Sex-Specific Neurological Impairments in Mice. Antioxidants, 8(4), 110. https://doi.org/10.3390/antiox8040110