Effect οf Genotype and Growing Year on the Nutritional, Phytochemical, and Antioxidant Properties of Industrial Hemp (Cannabis sativa L.) Seeds

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Material and Agricultural Conditions
2.3. Chemical Composition
2.4. Fatty Acid Composition
2.5. Tocopherols and Carotenoids Composition
2.6. Polyphenol Extraction
2.7. Total Phenolic Content
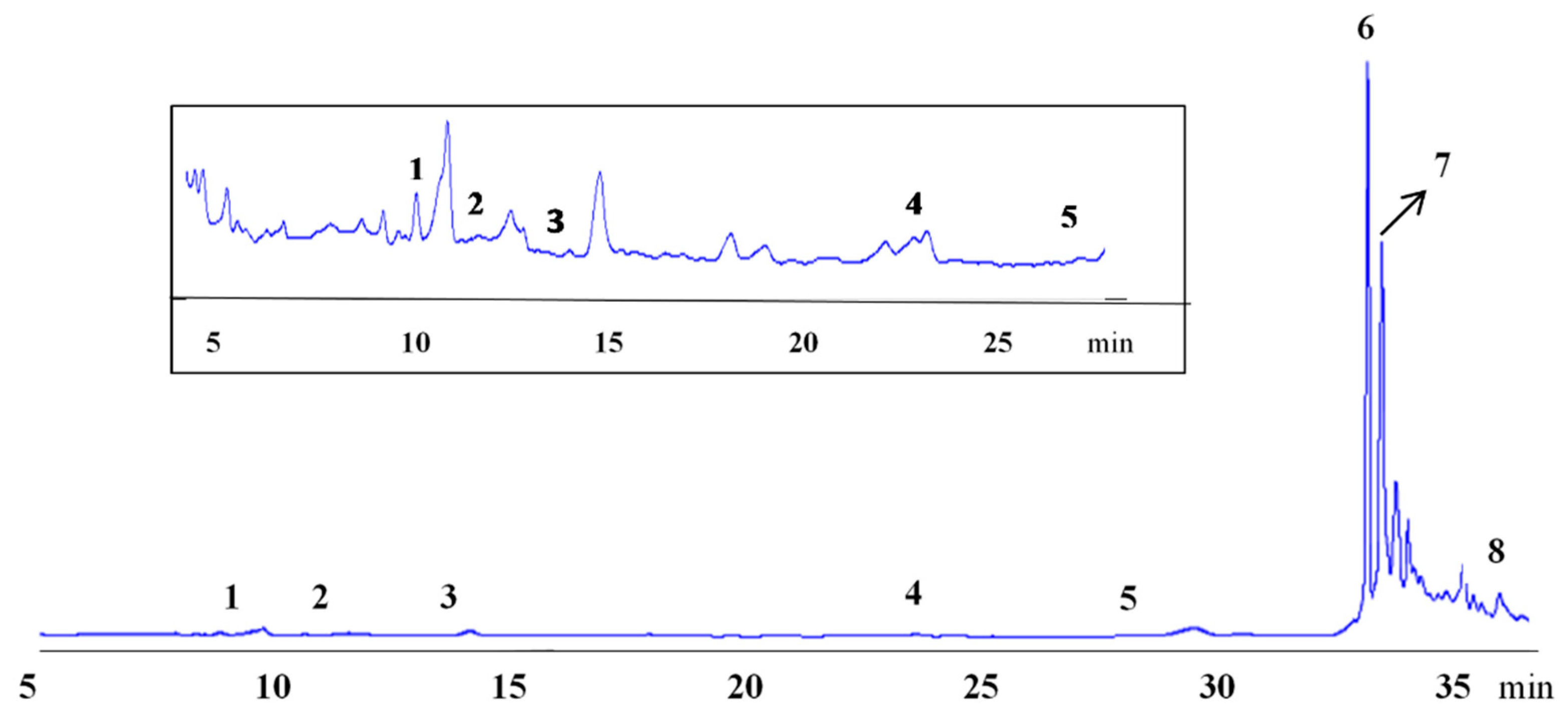
2.8. Phenolic Acid Composition
2.9. Antioxidant Activity Assays
2.10. Statistical Analysis
3. Results and Discussion
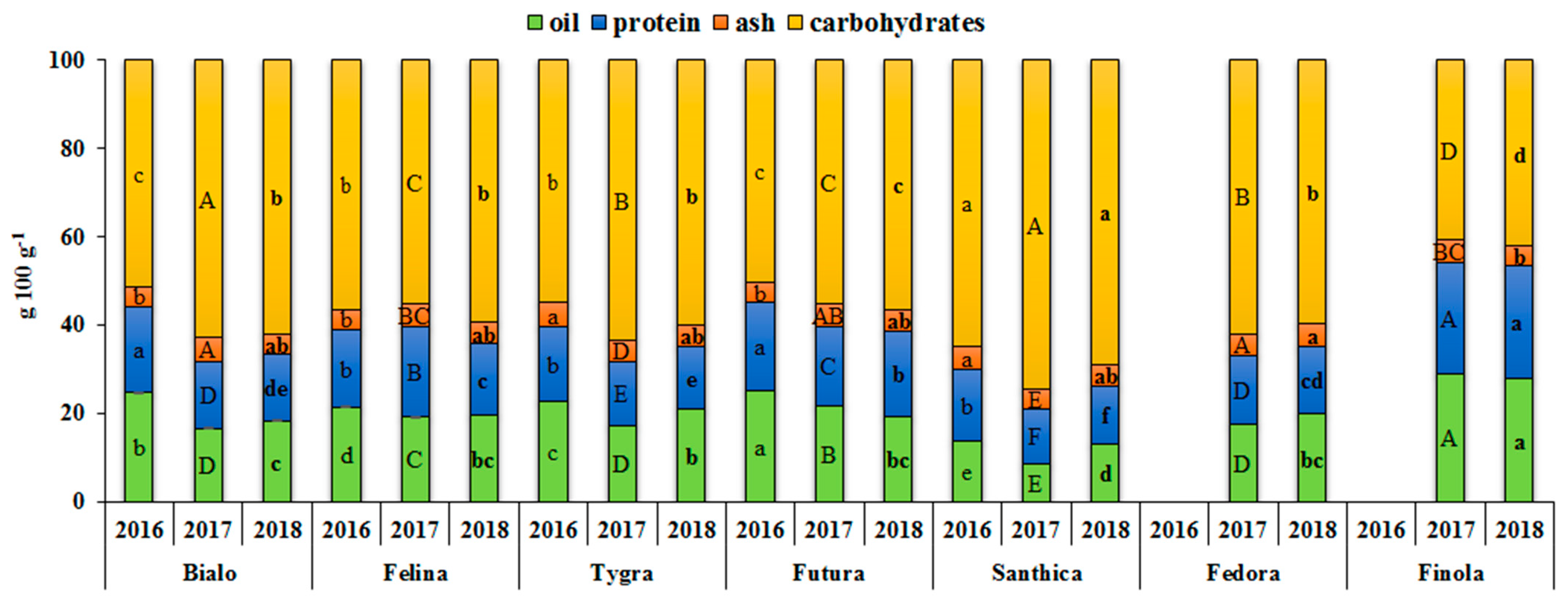
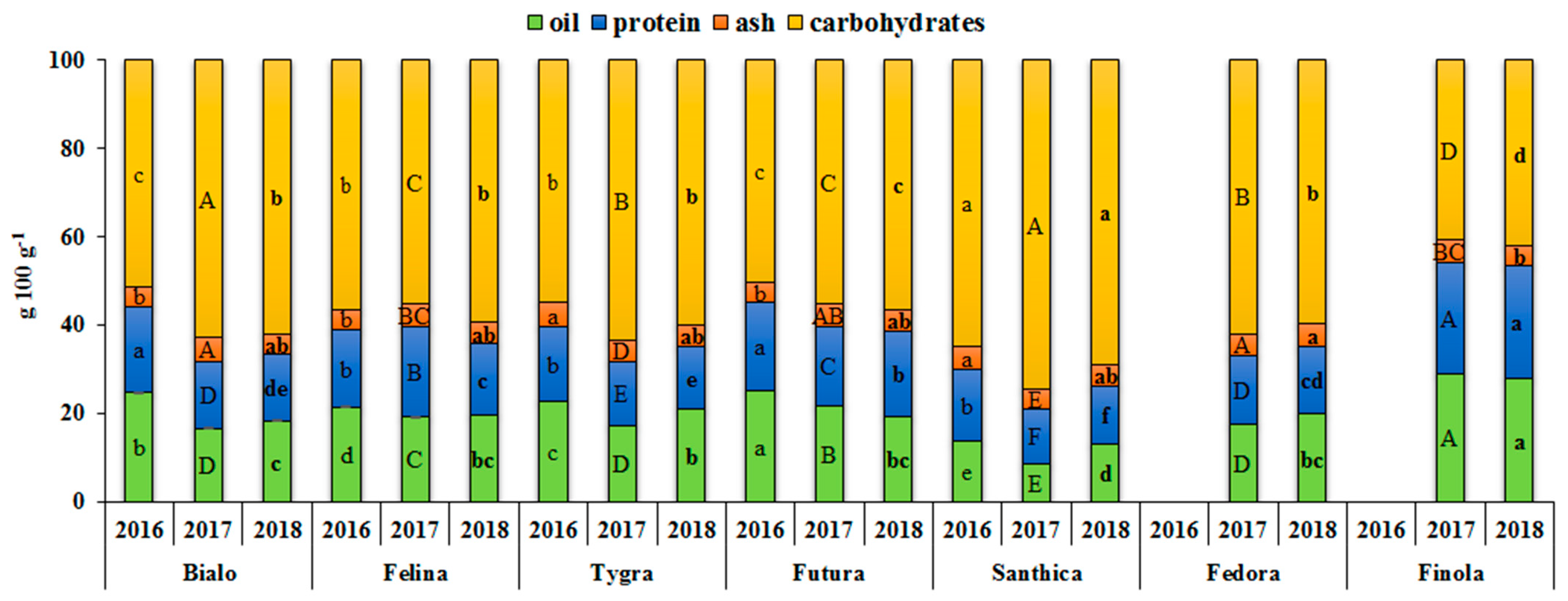
3.1. Effect of Genotype and Growing Year on the Nutritional Components
3.2. Effect of Genotype and Growing Year on the Fatty Acid Profile
3.3. Effect of Genotype and Growing Year on the Tocopherols and Carotenoid Content
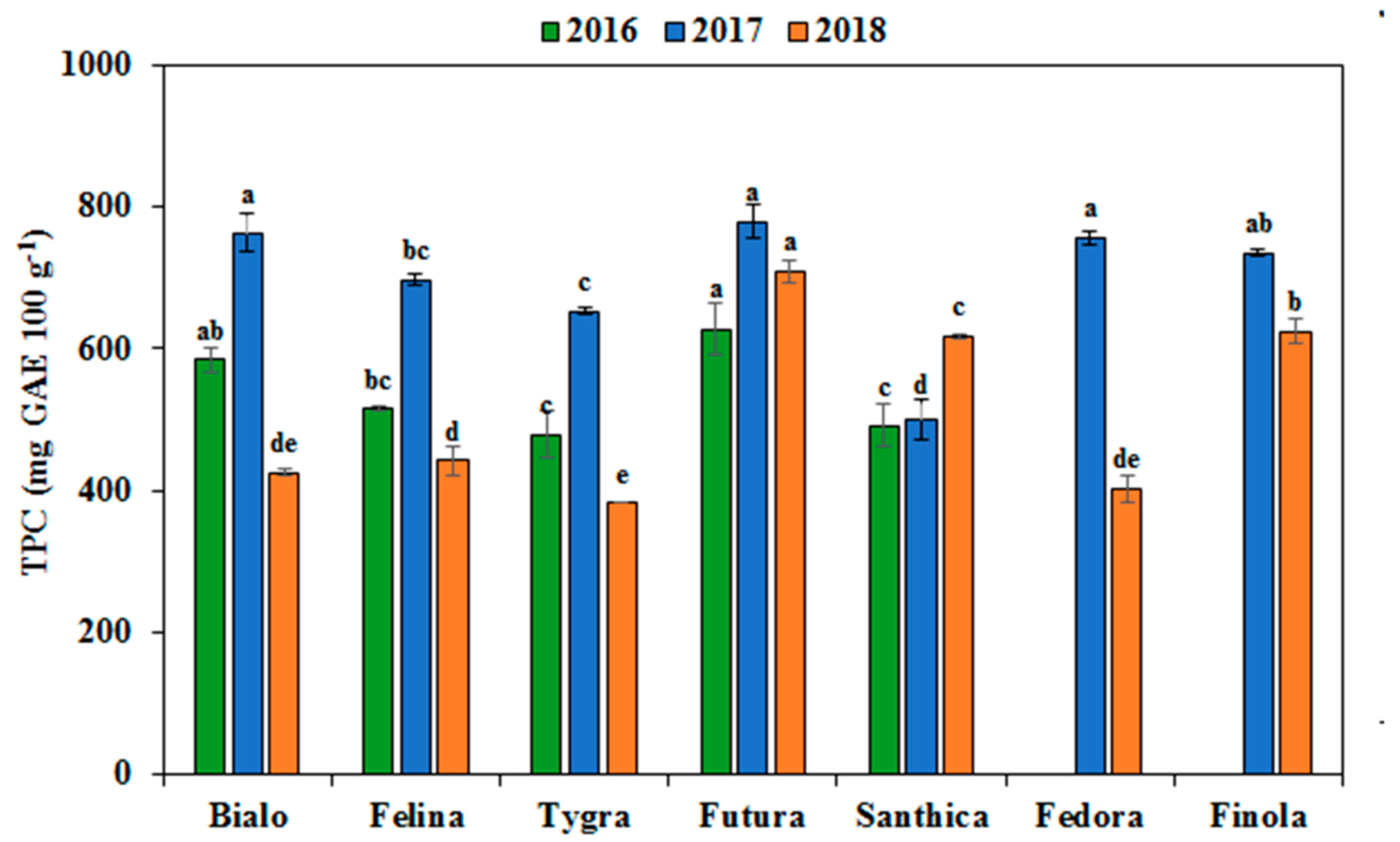
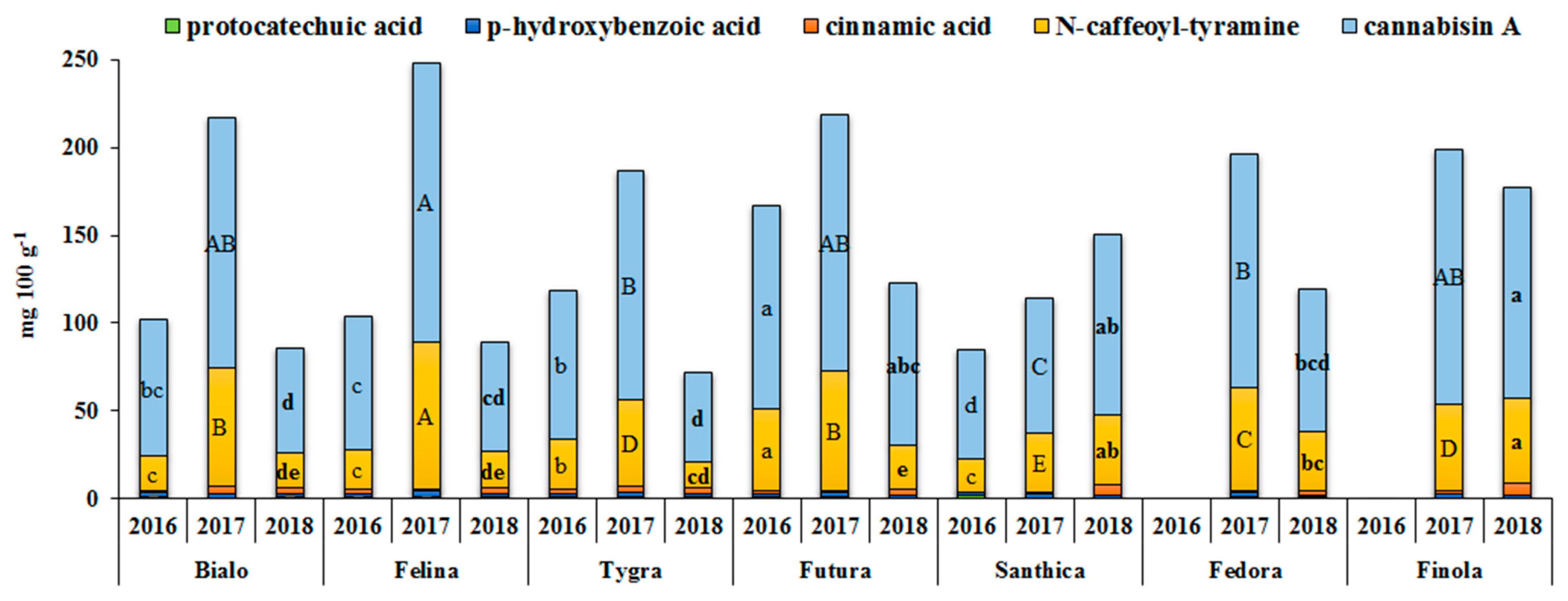
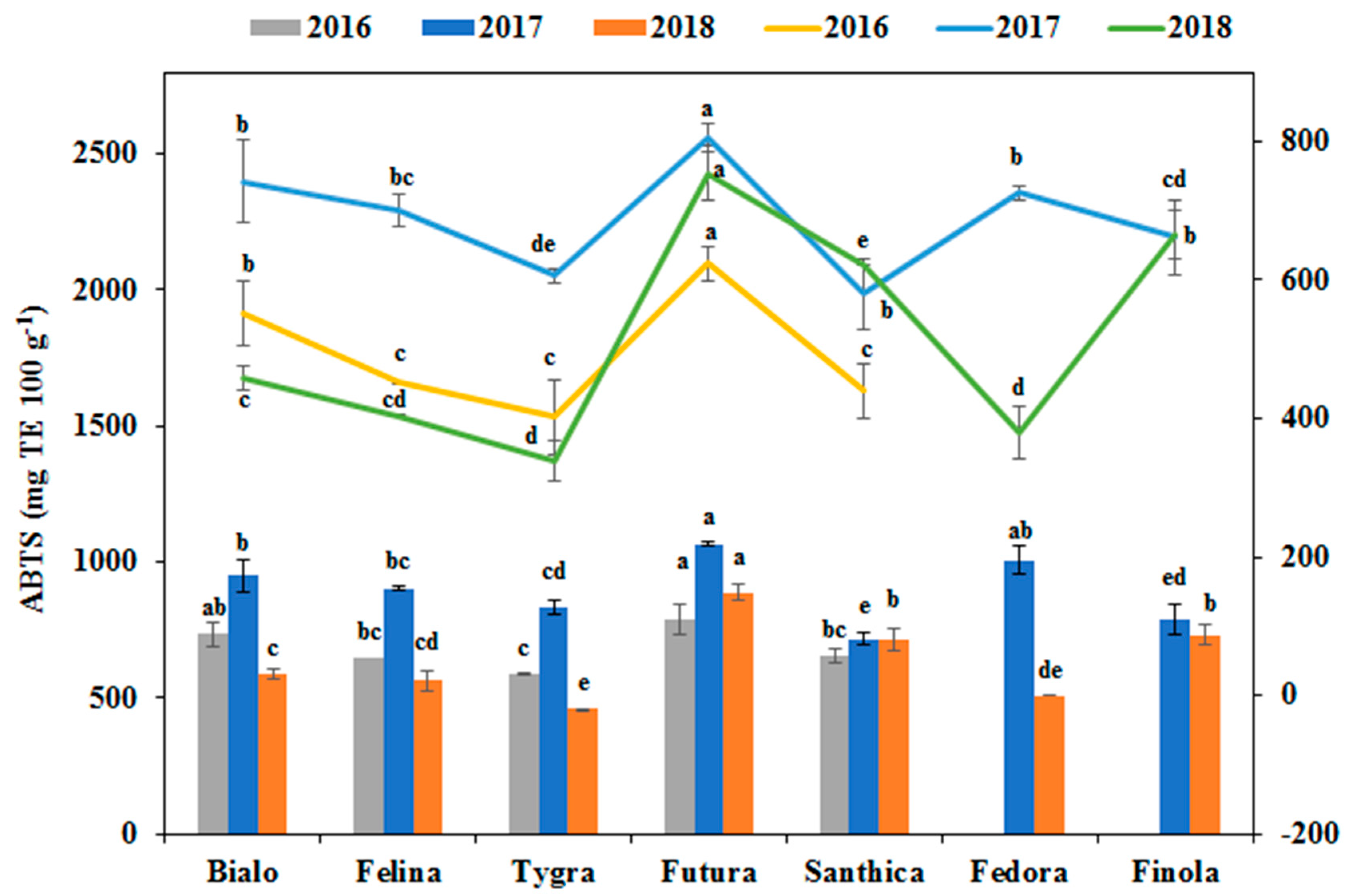
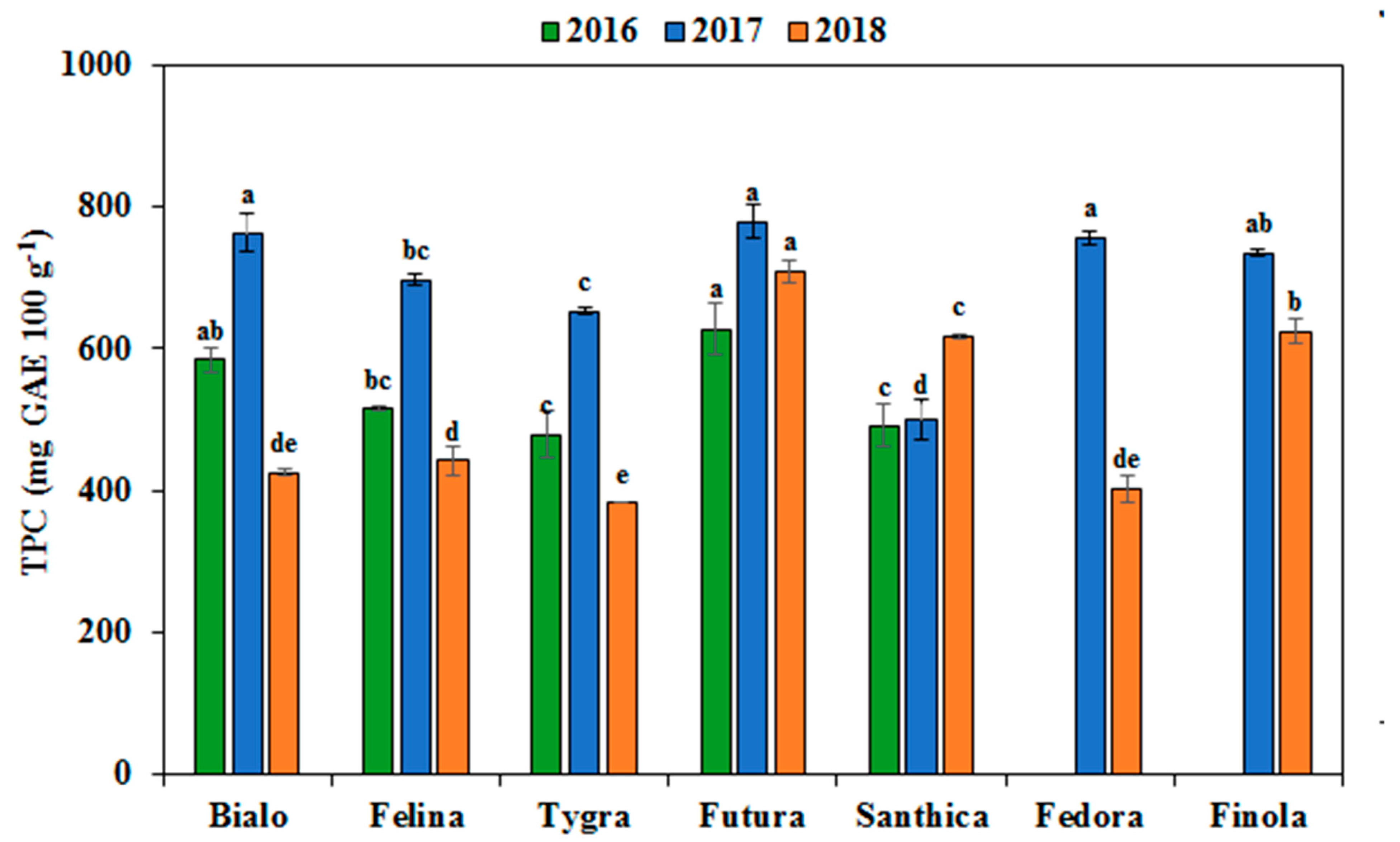
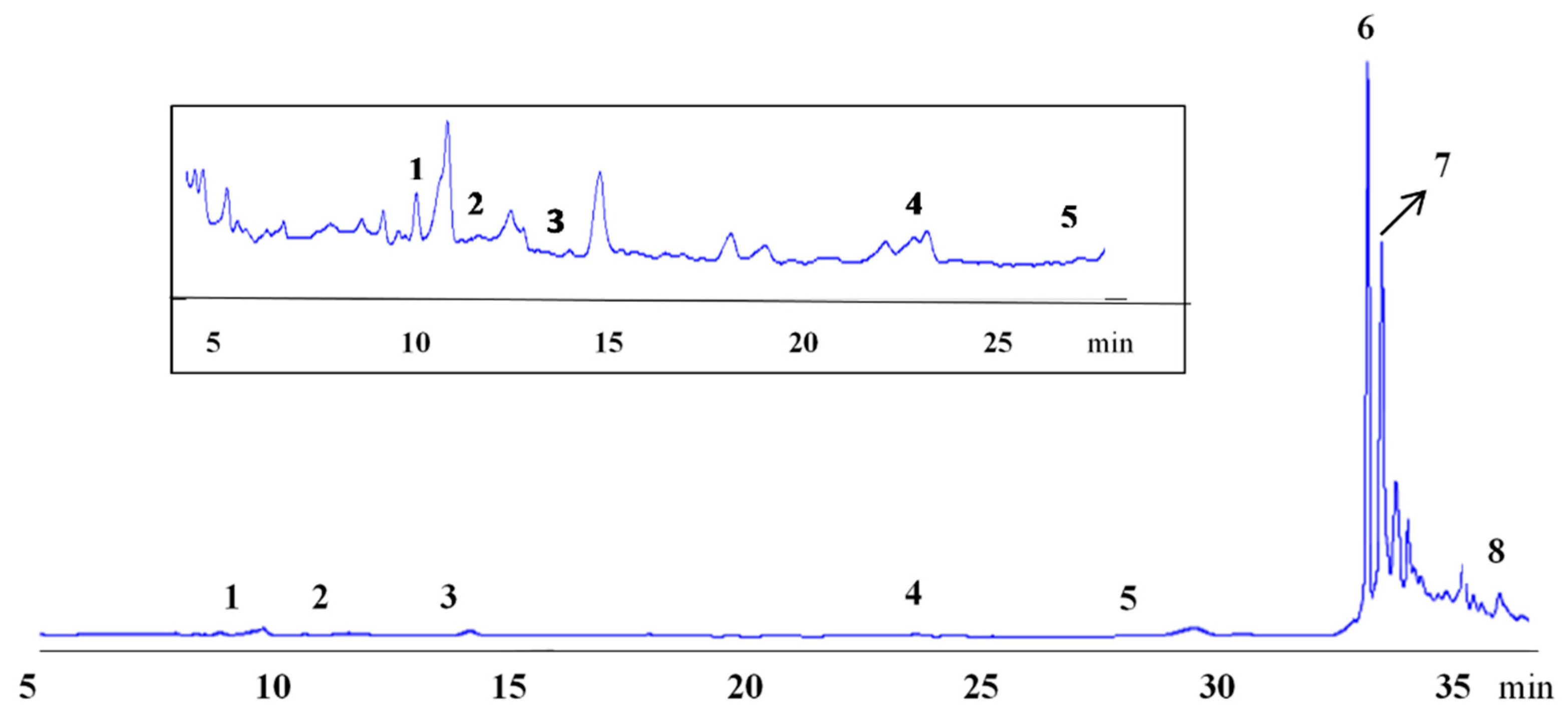
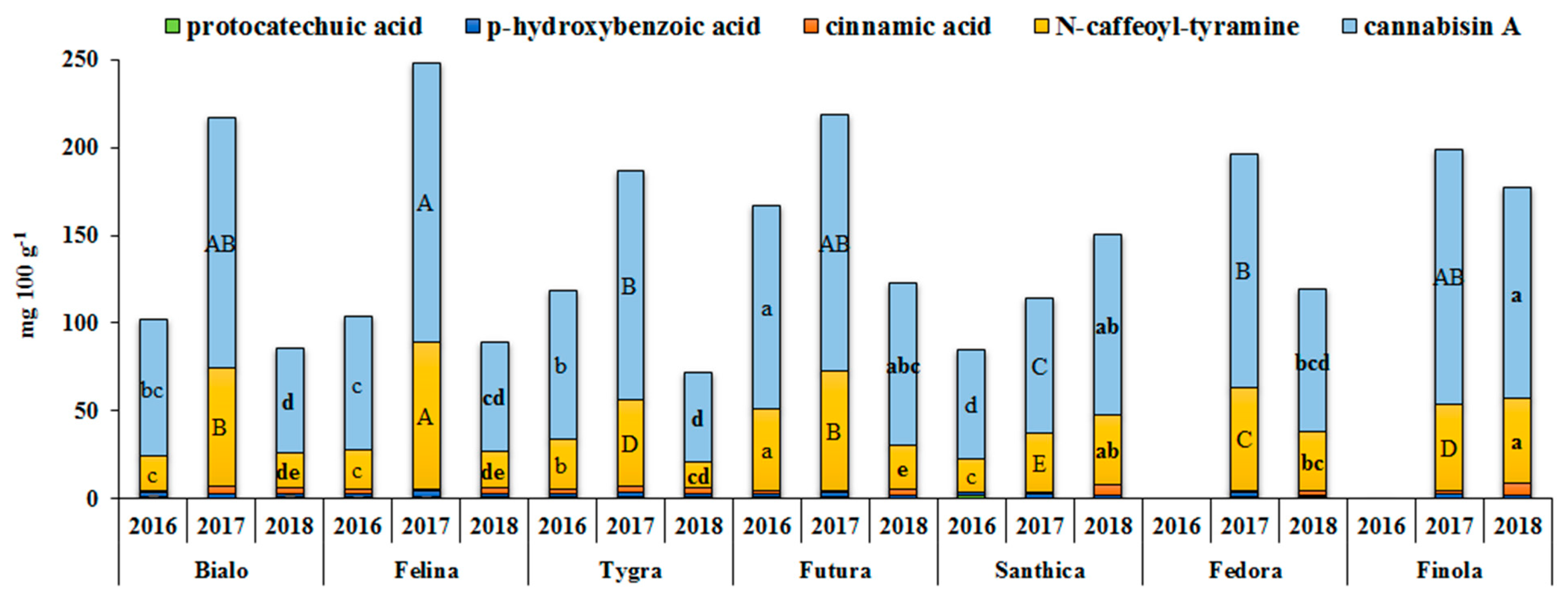
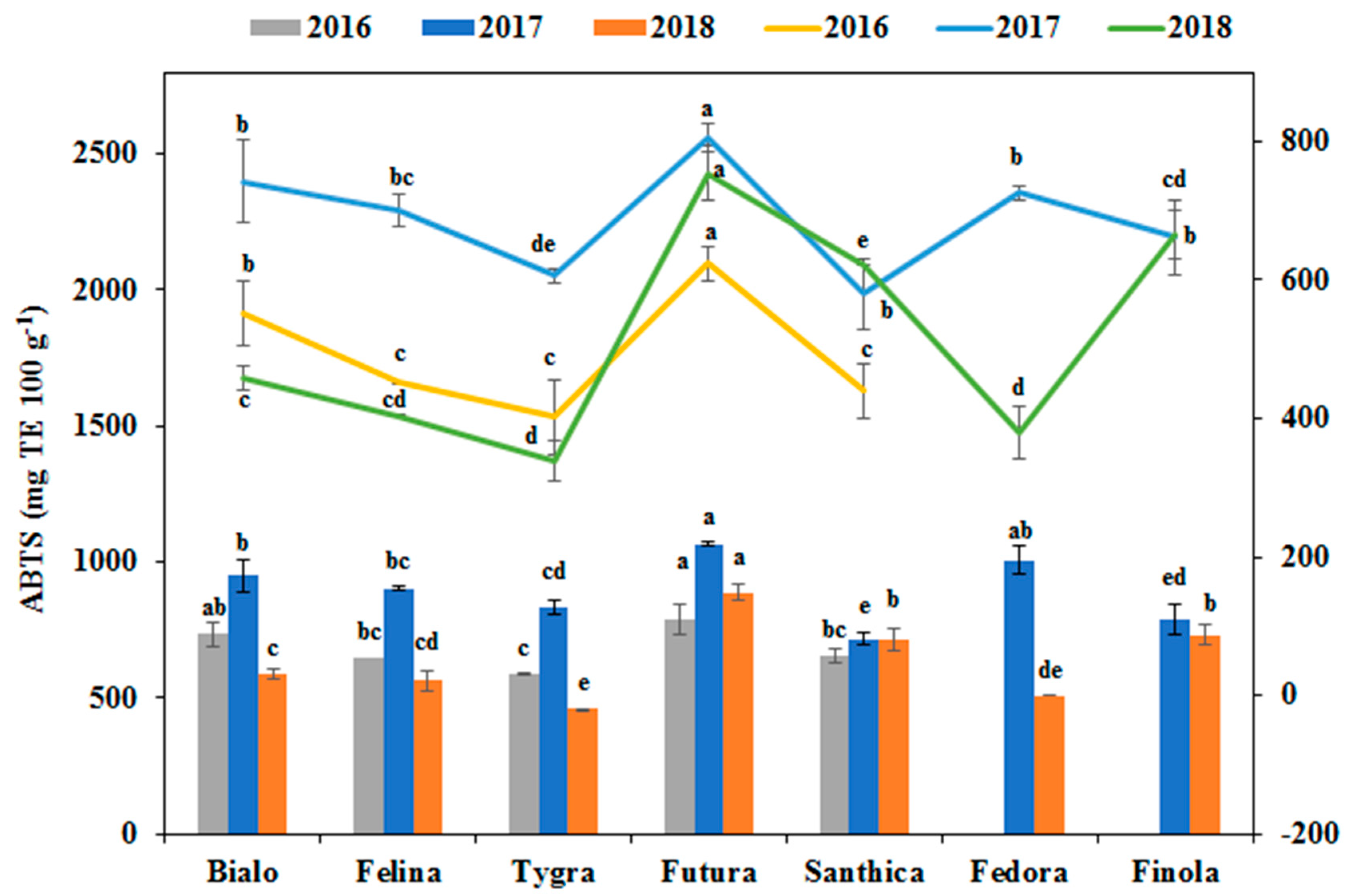
3.4. Effect of Genotype and Cultivation Year on the Phenolics Profile and Antioxidant Activity
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Amaducci, S.; Scordia, D.; Liu, F.H.; Zhang, Q.; Guo, H.; Testa, G.S.; Cosentino, L. Key cultivation techniques for hemp in Europe and China. Ind. Crops Prod. 2015, 68, 2–16. [Google Scholar] [CrossRef]
- Salentijn, E.M.J.; Zhang, Q.; Amaducci, S.; Yang, M.; Trindade, L.M. New developments in fiber hemp (Cannabis sativa L.) breeding. Ind. Crops Prod. 2015, 68, 32–41. [Google Scholar] [CrossRef]
- Andre, C.M.; Hausman, J.F.; Guerriero, G. Cannabis sativa: The plant of the thousand and one molecules. Front. Plant Sci. 2016, 7, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Callaway, J.C. Hemp seed as a nutritional resource: An overview. Euphytica 2004, 140, 65–72. [Google Scholar] [CrossRef]
- Kriese, U.; Schumann, E.; Weber, W.E.; Beyer, M.; Bruhl, L.; Matthaus, B. Oil content, tocopherol composition and fatty acid patterns of the seeds of 51 Cannabis sativa L. genotypes. Euphytica 2004, 137, 339–351. [Google Scholar] [CrossRef]
- Baldini, M.; Ferfuia, C.; Piani, B.; Sepulcri, A.; Dorigo, G.; Zuliani, F.; Danuso, F.; Cattivello, C. The performance and potentiality of monoecious hemp (Cannabis sativa L.) cultivars as a multipurpose crop. Agronomy 2018, 8, 162. [Google Scholar] [CrossRef]
- Fike, J. Industrial hemp: Renewed opportunities for an ancient crop. Crit. Rev. Plant Sci. 2016, 35, 406–424. [Google Scholar] [CrossRef]
- Kaul, N.; Kreml, R.; Austria, A.J.; Richard, M.N.; Edel, A.L.; Dibrov, E.; Hirono, S.; Zettler, M.E.; Pierce, G.N. A comparison of fish oil, flaxseed oil and hempseed oil supplementation on selected parameters of cardiovascular health in healthy volunteers. J. Am. Coll. Nutr. 2008, 27, 51–58. [Google Scholar] [CrossRef]
- Prociuk, M.A.; Edel, A.L.; Richard, M.N.; Gavel, N.T.; Ander, B.P.; Dupasquier, C.M.C.; Pierce, G.N. Cholesterol-induced stimulation of platelet aggregation is prevented by a hempseed-enriched diet. Can. J. Physiol. Pharmacol. 2008, 86, 153–159. [Google Scholar] [CrossRef]
- Smeriglio, A.; Galati, E.M.; Monforte, M.T.; Lanuzza, F.; D’Angelo, V.; Circosta, C. Polyphenolic compounds and antioxidant activity of cold-pressed seed oil from Finola cultivar of Cannabis sativa L. Phytother. Res. 2016, 30, 1298–1307. [Google Scholar] [CrossRef]
- Pollastroa, F.; Minassia, A.; Fresu, L.G. Cannabis phenolics and their bioactivities. Curr. Med. Chem. 2018, 25, 1160–1185. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Tang, J.; dos Santos Passos, C.; Nurisso, A.; Simões-Pires, C.A.; Ji, M.; Lou, H.; Fan, P. Characterization of lignanamides from hemp (Cannabis sativa L.) seed and their antioxidant and acetylcholinesterase inhibitory activities. J. Agric. Food Chem. 2015, 63, 10611–10619. [Google Scholar] [CrossRef] [PubMed]
- Legros, S.; Picault, S.; Cerruti, N. Factors affecting the yield of industrial hemp—Experimental results from France. In Hemp Industrial Production and Uses; Bouloc, P., Ed.; CABI: Wollingford, UK, 2013; pp. 72–97. [Google Scholar]
- Blade, S.F.; Ampong-Nyarko, K.; Przybylski, R. Fatty acid and tocopherol profiles of industrial hemp cultivars grown in the high latitude prairie region of Canada. J. Ind. Hemp 2005, 10, 33–43. [Google Scholar] [CrossRef]
- Vonapartis, E.; Aubin, M.P.; Seguin, P.; Mustafa, A.F.; Charron, J.B. Seed composition of ten industrial hemp cultivars approved for production in Canada. J. Food Comp. Anal. 2015, 39, 8–12. [Google Scholar] [CrossRef]
- Siano, F.; Moccia, S.; Picariello, G.; Russo, G.L.; Sorrentino, G.; Di Stasio, M.; Cara, F.L.; Volpe, M.G. Comparative study of chemical, biochemical characteristic and ATR-FTIR analysis of seeds, oil and flour of the edible Fedora cultivar hemp (Cannabis sativa L.). Molecules 2019, 24, 83. [Google Scholar] [CrossRef]
- Tang, K.; Struik, P.; Yin, C.X.; Thouminot, C.; Bjelková, M.; Stramkale, V.; Amaducci, S. Comparing hemp (Cannabis sativa L.) cultivars for dual-purpose production under contrasting environments. Ind. Crops Prod. 2016, 87, 33–44. [Google Scholar] [CrossRef]
- European Commission. Commission Regulation (EC) No. 809/2014 of 17 July 2014 laying down rules for the application of Regulation (EU) No 1306/2013 of the European Parliament and of the Council with regard to the integrated administration and control system, rural development measures and cross compliance. Off. J. Eur. Union 2014, L227, 69–124. [Google Scholar]
- AOAC International. Official Methods of Analysis, 20th ed.; Association of Official Agricultural Chemists: Rockville, MD, USA, 2016. [Google Scholar]
- AOAC Method 996-06. Fats (total, saturated and unsaturated) in foods. In AOAC Official Methods of Analysis; Association of Official Analytical Chemists Inc.: Gaithersburg, MD, USA, 2002. [Google Scholar]
- Irakli, M.; Chatzopoulou, P.; Kadoglidou, K.; Tsivelika, N. Optimization and development of a high-performance liquid chromatography method for the simultaneous determination of vitamin E and carotenoids in tomato fruits. J. Sep. Sci. 2016, 39, 3348–3356. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagents. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Skendi, A.; Irakli, M.; Chatzopoulou, P. Analysis of phenolic compounds in Greek plants of Lamiaceae family by HPLC. J. Appl. Res. Med. Arom. Plants 2017, 6, 62–69. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C.A. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Benzie, F.; Strain, J. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. Methods Enzymol. 1999, 299, 15–23. [Google Scholar] [PubMed]
- House, J.D.; Neufeld, J.; Leson, G. Evaluating the quality of protein from hemp seed (Cannabis sativa L.) products through the use of the protein digestibility-corrected amino acid score method. J. Agric. Food Chem. 2010, 58, 11801–11807. [Google Scholar] [CrossRef] [PubMed]
- Galasso, I.; Russo, R.; Mapelli, S.; Ponzoni, E.; Brambilla, I.M.; Battelli, G.; Reggiani, R. Variability in seed traits in a collection of Cannabis sativa L. genotypes. Front. Plant Sci. 2016, 7, 688–697. [Google Scholar] [CrossRef] [PubMed]
- Kinsella, J.; Lokesh, B.; Stone, R. Dietary n-3 polyunsaturated acids and amelioration of cardiovascular disease: Possible mechanisms. Am. J. Clin. Nutr. 1990, 52, 1–28. [Google Scholar] [CrossRef]
- Layne, K.S.; Goh, Y.K.; Jumpsen, J.; Ryan, R.A.; Chow, P.; Clandinin, M.T. Normal subjects consuming physiological levels of 18:3(n-3) and 20:5(n-3) from flaxseed or fish oils have characteristic differences in plasma lipid and lipoprotein fatty acid levels. J. Nutr. 1996, 126, 2130–2140. [Google Scholar] [CrossRef]
- Da Porto, C.; Decorti, D.; Natolino, A. Potential oil yield, fatty acid composition, and oxidation stability of the hempseed oil from four Cannabis sativa L. cultivars. J. Diet. Suppl. 2015, 12, 1–10. [Google Scholar] [CrossRef]
- Sies, H.; Murphy, M.E. Role of tocopherols in the protection of biological systems against oxidative damage. J. Photochem. Photobiol. B Biol. 1991, 8, 211–224. [Google Scholar] [CrossRef]
- Liang, J.; Aachary, A.A.; Thiyam-Holländer, U. Hemp seed oil: Minor components and oil quality. Lipid Technol. 2015, 27, 231–233. [Google Scholar] [CrossRef]
- Aladić, K.; Jokić, S.; Moslavac, T.; Tomas, S.; Vidović, J.; Vladić, S.; Šubarić, D. Cold pressing and supercritical CO2 extraction of hemp (Cannabis sativa) seed oil. Chem. Biochem. Eng. 2014, 28, 481–490. [Google Scholar] [CrossRef]
- Okarter, N.; Liu, C.; Sorrells, M.E.; Liu, R.H. Phytochemical content and antioxidant activity of six diverse varieties of whole wheat. Food Chem. 2010, 119, 249–257. [Google Scholar] [CrossRef]
- Arts, I.C.; Hollman, P.C. Polyphenols and disease risk in epidemiologic studies. Am. J. Clin. Nutr. 2005, 81, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Teh, S.S.; Birch, J. Physicochemical and quality characteristics of cold-pressed hemp, flax and canola seed oils. J. Food Comp. Anal. 2013, 30, 26–31. [Google Scholar] [CrossRef]
- Chen, T.; He, J.; Zhang, J.; Li, X.; Zhang, H.; Hao, J.; Li, L. The isolation and identification of two compounds with predominant radical scavenging activity in hempseed (seed of Cannabis sativa L.). Food Chem. 2012, 134, 1030–1037. [Google Scholar] [CrossRef]
- Pagnani, G.; Pellegrini, M.; Galieni, A.; D’Egidio, S.; Matteucci, F.; Ricci, A.; Stagnari, F.; Sergi, M.; Lo Sterzo, C.; Pisante, M.; et al. Plant growth-promoting rhizobacteria (PGPR) in Cannabis sativa ‘Finola’ cultivation: An alternative fertilization strategy to improve plant growth and quality characteristics. Ind. Crops Prod. 2018, 123, 75–83. [Google Scholar] [CrossRef]
- Heimler, D.; Vignolini, P.; Isolani, L.; Arfaioli, P.; Ghiselli, L.; Romani, A. Polyphenol content of modern and old varieties of Triticum aestivum L. and T. durum Desf. grains in two years of production. J. Agric. Food Chem. 2010, 58, 7329–7334. [Google Scholar] [CrossRef]
- Sakakibara, I.; Katsuhara, T.; Ikeya, Y.; Hayashi, K.; Mitsuhashi, H. Cannabisin A, an arylnaphthalene lignanamide from fruits of Cannabis sativa. Phytochemistry 1991, 30, 3013–3016. [Google Scholar] [CrossRef]
- Lesma, G.; Consonni, R.; Gambaro, V.; Remuzzi, C.; Roda, G.; Silvani, A.; Vece, V.; Visconti, G.L. Cannabinoid-free Cannabis sativa L. grown in the Po valley: Evaluation of fatty acid profile, antioxidant capacity and metabolic content. Nat. Prod. Res. 2014, 28, 1801–1807. [Google Scholar] [CrossRef]
- Wang, C.Z.; Ma, X.Q.; Yang, D.H.; Guo, Z.R.; Liu, G.R.; Zhao, G.X.; Tang, J.; Zhang, Y.N.; Ma, M.; Cai, S.Q.; et al. Production of enterodiol from defatted flax seeds through biotransformation by human intestinal bacteria. BMC Microbiol. 2010, 10, 115–123. [Google Scholar] [CrossRef]
- Pojić, M.; Mišan, A.; Sakač, M.; DapčevićHadnađev, T.; Šarić, B.; Milovanović, I.; Hadnađev, M. Characterization of byproducts originating from hemp oil processing. J. Agric. Food Chem. 2014, 62, 12436–12442. [Google Scholar] [CrossRef]
- Frassinetti, S.; Moccia, E.; Caltavuturo, L.; Gabriele, M.; Longo, V.; Bellani, L.; Giorgi, G.; Giorgetti, L. Nutraceutical potential of hemp (Cannabis sativa L.) seeds and sprouts. Food Chem. 2018, 262, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Jeong, H.-S.; Lee, J. Antioxidant activity of methanolic extracts from some grains consumed in Korea. Food Chem. 2007, 103, 130–138. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Abbreviations | Origin | Species |
|---|---|---|---|
| Bialobrzeskie | Bialo | Poland | monoecious |
| Felina 32 | Felina | France | monoecious |
| Tygra | Tygra | Poland | monoecious |
| Futura 75 | Futura | France | monoecious |
| Santhica 27 | Santhica | France | monoecious |
| Fedora 17 | Fedora | France | monoecious |
| Finola | Finola | Finland | dioecious |
| Month | 2016 | 2017 | 2018 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Tmin (°C) | Tmax (°C) | Rainfall (mm) | Tmin (°C) | Tmax (°C) | Rainfall (mm) | Tmin (°C) | Tmax (°C) | Rainfall (mm) | |
| April | 4 | 30 | 8.1 | 3 | 26 | 9.2 | 6 | 31 | 4.9 |
| May | 9 | 32 | 75.2 | 11 | 30 | 51.8 | 12 | 30 | 93.0 |
| June | 14 | 38 | 13.3 | 14 | 39 | 13.2 | 16 | 33 | 106.8 |
| July | 17 | 36 | 0.7 | 16 | 40 | 78.3 | 17 | 34 | 26.1 |
| August | 18 | 36 | 56.6 | 14 | 38 | 12.9 | 19 | 36 | 12.0 |
| September | 9 | 31 | 80.0 | 12 | 35 | 2.7 | 9 | 34 | 5.0 |
| Mean | 12 | 34 | 12 | 35 | 13 | 33 | |||
| Total | 233.9 | 168.1 | 247.8 | ||||||
| Parameter | Genotype (G) | Total Variance (%) | Year (Y) | Total Variance (%) | GxY | Total Variance (%) |
|---|---|---|---|---|---|---|
| Nutritional Components | ||||||
| Oil | 145.5*** | 58.8 | 92.5*** | 37.4 | 9.5*** | 3.9 |
| Protein | 38.4*** | 52.4 | 27.2*** | 37.0 | 7.8*** | 10.6 |
| Ash | 0.1*** | 9.6 | 0.3*** | 36.9 | 0.4*** | 53.6 |
| Carbohydrates | 30.4.4*** | 57.7 | 199.0*** | 37.7 | 24.5*** | 4.6 |
| Fatty Acids | ||||||
| Palmitic | 2.0* | 86.2 | 0.1*** | 6.0 | 0.0*** | 7.8 |
| Stearic | 0.3*** | 70.0 | 0.1*** | 25.7 | 0.0NS | 4.3 |
| Oleic | 39.0*** | 91.2 | 2.3*** | 5.5 | 1.4*** | 3.3 |
| Linoleic | 2.8*** | 42.0 | 3.2*** | 47.7 | 0.7*** | 10.3 |
| γ-linolenic | 3.8*** | 77.3 | 0.9*** | 18.0 | 0.2*** | 4.7 |
| α-linolenic | 13.0*** | 99.6 | 0.5*** | 3.3 | 0.9*** | 4.7 |
| ω-6/ω-3 ratio | 1.1NS | 85.3 | 0.0*** | 1.3 | 0.1*** | 13.4 |
| Tocopherols | ||||||
| γ-Τ | 9.1*** | 24.4 | 26.4*** | 71.0 | 1.7*** | 4.6 |
| δ-Τ | 0.0*** | 9.9 | 0.2*** | 85.9 | 0.0*** | 4.2 |
| Carotenoids | ||||||
| Lutein | 1.0*** | 43.7 | 0.0*** | 12.6 | 1.0*** | 43.7 |
| β-carotene | 0.3*** | 63.5 | 0.1*** | 28.3 | 0.0*** | 8.1 |
| Zeaxanthin | 0.0*** | 10.1 | 0.0*** | 64.9 | 0.0*** | 25.0 |
| Total | 2.0*** | 54.3 | 0.3*** | 7.4 | 1.4*** | 38.4 |
| Total Phenolic Content | 54,456.7*** | 27.3 | 117,289.3*** | 58.9 | 27,466.2*** | 13.8 |
| Phenolic Compounds | ||||||
| Protocatechuic acid | 0.3*** | 17.5 | 1.3*** | 66.6 | 0.3*** | 15.8 |
| p-hydroxybenzoic acid | 0.1*** | 2.9 | 5.4*** | 93.5 | 0.2*** | 3.7 |
| Cinnamic acid | 1.1*** | 2.9 | 29.1*** | 77.6 | 7.4*** | 19.6 |
| N-trans-caffeoyltyramine | 458.1*** | 6.5 | 5936.6*** | 84.0 | 674.7*** | 9.5 |
| Cannabisin A | 1738.1*** | 9.7 | 14,174.2*** | 79.1 | 1998.5*** | 11.2 |
| Antioxidant Activity | ||||||
| ABTS assay | 105,299.8*** | 26.3 | 272,547.7*** | 68.1 | 22,639.5*** | 5.7 |
| FRAP assay | 96,613.4*** | 33.7 | 169,163.2*** | 59.0 | 20,773.0*** | 7.3 |
| Genotype | Year | Fatty Acids (%) | ω-6/ω-3 | |||||
|---|---|---|---|---|---|---|---|---|
| Palmitic | Stearic | Oleic | Linoleic | α-Linolenic | γ-Linolenic | |||
| Bialo | 2016 | 8.5 ab | 2.4 ab | 15.3 b | 53.6 a | 10.5 b | 3.4 a | 5.5 a |
| Felina | 8.3 ab | 2.4 ab | 13.5 c | 53.7 a | 12.2 a | 2.9 b | 4.6 c | |
| Tygra | 8.2 ab | 2.7 a | 16.2 ab | 51.6 b | 11.7 a | 2.9 b | 4.7 bc | |
| Futura | 7.8 b | 2.5 ab | 16.7 a | 52.9 a | 10.8 b | 2.1 c | 5.1 ab | |
| Santhica | 8.9 a | 2.3 b | 12.4 d | 54.0 a | 12.1 a | 3.1 b | 4.7 bc | |
| Fedora | - | - | - | - | - | - | - | |
| Finola | - | - | - | - | - | - | - | |
| Bialo | 2017 | 8.6 B | 2.6 A | 15.8 B | 52.7 B | 11.4 D | 2.6 C | 4.9 BC |
| Felina | 7.9 C | 2.5 A | 13.6 C | 53.5 AB | 13.0 B | 2.6 C | 4.4 D | |
| Tygra | 8.5 B | 2.8 A | 17.6 A | 51.6 C | 10.6 E | 2.6 C | 5.5 A | |
| Futura | 8.5 B | 2.8 A | 17.9 A | 52.9 AB | 10.7 E | 1.9 D | 5.2 A | |
| Santhica | 9.1 A | 2.5 A | 12.6 D | 53.9 A | 12.1 C | 3.0 B | 4.7 C | |
| Fedora | 8.6 B | 2.6 A | 15.8 B | 52.9 AB | 11.2 D | 2.9 B | 5.0 AB | |
| Finola | 7.4 D | 2.1 B | 10.6 E | 53.8 A | 15.1 A | 4.5 A | 3.9 E | |
| Bialo | 2018 | 8.5 ab | 2.5 ab | 16.0 a | 53.5 b | 11.4 cd | 2.8 c | 5.0 b |
| Felina | 8.4 ab | 2.5 ab | 14.7 b | 54.2 a | 11.9 b | 2.9 bc | 4.9 bc | |
| Tygra | 8.3 b | 2.6 a | 15.4 ab | 53.7 b | 11.7 bc | 2.9 bc | 4.8 bc | |
| Futura | 7.8 c | 2.7 a | 16.0 a | 53.7 b | 12.1 b | 3.0 c | 4.7 c | |
| Santhica | 8.8 a | 2.3 b | 13.2 c | 54.2 a | 12.1 b | 3.2 b | 4.7 c | |
| Fedora | 8.7 ab | 2.6 a | 15.1 b | 53.7 b | 11.2 d | 3.1 bc | 5.1 a | |
| Finola | 7.1 d | 2.1 c | 10.3 d | 53.7 b | 15.3 a | 5.0 a | 3.9 d | |
| Genotype | Year | Tocopherols | Carotenoids | ||||
|---|---|---|---|---|---|---|---|
| γ-Τ | δ-Τ | Lutein | β-Carotene | Zeaxanthin | Total | ||
| Bialo | 2016 | 8.5 a | 0.8 c | 2.4 a | 0.2 d | 0.3 b | 2.9 b |
| Felina | 8.6 a | 1.0 a | 2.1 b | 0.6 b | 0.3 b | 3.0 b | |
| Tygra | 7.9 b | 0.9 ab | 1.8 c | 0.5 c | 0.3 b | 2.6 c | |
| Futura | 8.8 a | 0.9 b | 1.7 c | 0.2 d | 0.2 d | 2.1 d | |
| Santhica | 6.4 c | 0.8 c | 2.3 ab | 0.7 a | 0.4 a | 3.4 a | |
| Fedora | - | - | - | - | - | ||
| Finola | - | - | - | - | - | ||
| Bialo | 2017 | 6.3 D | 0.7 BC | 0.9 E | 0.2 E | 0.3 C | 1.4 E |
| Felina | 8.0 B | 0.8 A | 2.4 BC | 0.4 CD | 0.4 B | 3.2 C | |
| Tygra | 6.5 CD | 0.7 BC | 2.0 D | 0.6 A | 0.4 B | 3.0 C | |
| Futura | 8.9 A | 0.7 BC | 2.6 B | 0.3 D | 0.5 A | 3.4 B | |
| Santhica | 4.6 E | 0.6 C | 3.4 A | 0.5 B | 0.4 B | 4.3 A | |
| Fedora | 7.0 C | 0.8 A | 2.2 CD | 0.4 C | 0.3 C | 2.9 CD | |
| Finola | 8.4 B | 0.8 A | 2.2 CD | 0.2 E | 0.2 D | 2.6 D | |
| Bialo | 2018 | 5.2 cd | 0.6 c | 2.3 a | 0.5 cd | 0.3 a | 3.1 a |
| Felina | 4.8 d | 0.6 c | 2.0 bc | 0.6 bc | 0.3 ab | 3.0 ab | |
| Tygra | 5.6 c | 0.7 b | 1.7 de | 0.8 a | 0.2 bc | 2.7 b | |
| Futura | 6.5 b | 0.7 b | 1.5 e | 0.3 d | 0.2 c | 2.0 c | |
| Santhica | 4.9 d | 0.6 c | 2.3 a | 0.6 bc | 0.3 ab | 3.2 a | |
| Fedora | 5.4 cd | 0.6 c | 1.8 cd | 0.7 ab | 0.2 c | 2.8 b | |
| Finola | 11.3 a | 1.3 a | 2.2 ab | 0.3 d | 0.4 a | 2.9 b | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Irakli, M.; Tsaliki, E.; Kalivas, A.; Kleisiaris, F.; Sarrou, E.; Cook, C.M. Effect οf Genotype and Growing Year on the Nutritional, Phytochemical, and Antioxidant Properties of Industrial Hemp (Cannabis sativa L.) Seeds. Antioxidants 2019, 8, 491. https://doi.org/10.3390/antiox8100491
Irakli M, Tsaliki E, Kalivas A, Kleisiaris F, Sarrou E, Cook CM. Effect οf Genotype and Growing Year on the Nutritional, Phytochemical, and Antioxidant Properties of Industrial Hemp (Cannabis sativa L.) Seeds. Antioxidants. 2019; 8(10):491. https://doi.org/10.3390/antiox8100491
Chicago/Turabian StyleIrakli, Maria, Eleni Tsaliki, Apostolos Kalivas, Fotios Kleisiaris, Eirini Sarrou, and Catherine M Cook. 2019. "Effect οf Genotype and Growing Year on the Nutritional, Phytochemical, and Antioxidant Properties of Industrial Hemp (Cannabis sativa L.) Seeds" Antioxidants 8, no. 10: 491. https://doi.org/10.3390/antiox8100491
APA StyleIrakli, M., Tsaliki, E., Kalivas, A., Kleisiaris, F., Sarrou, E., & Cook, C. M. (2019). Effect οf Genotype and Growing Year on the Nutritional, Phytochemical, and Antioxidant Properties of Industrial Hemp (Cannabis sativa L.) Seeds. Antioxidants, 8(10), 491. https://doi.org/10.3390/antiox8100491