Carotenoids of Capsicum Fruits: Pigment Profile and Health-Promoting Functional Attributes

 ,
,
Abstract
:1. Introduction
2. The Capsicum Species
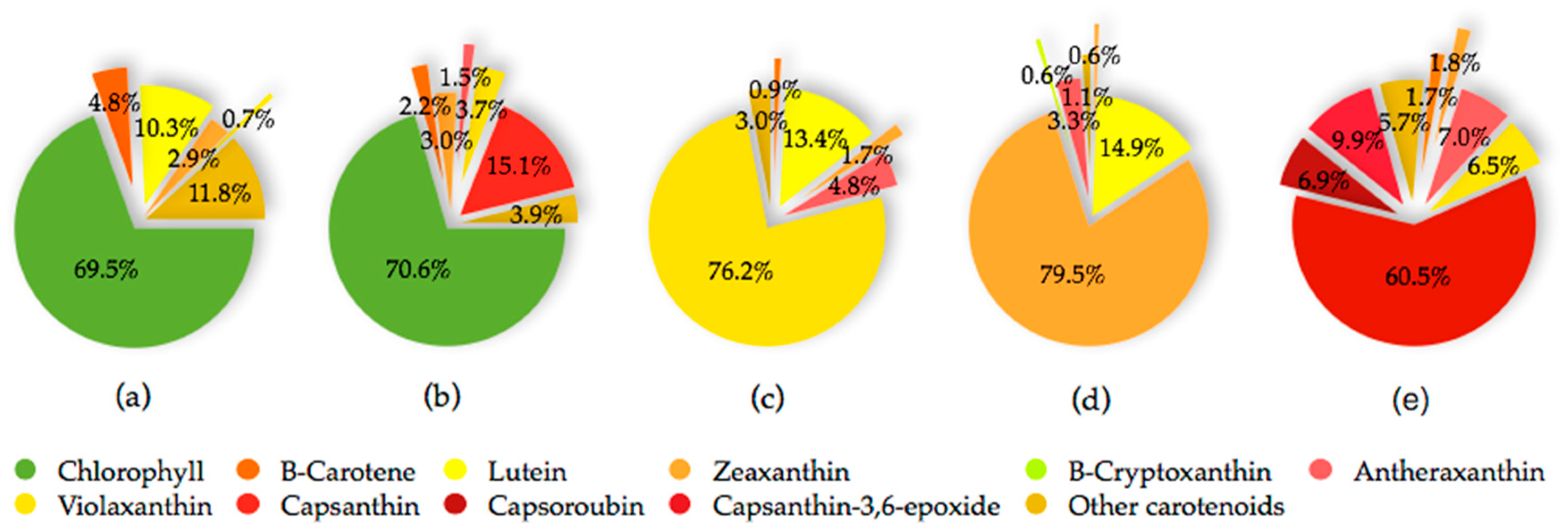
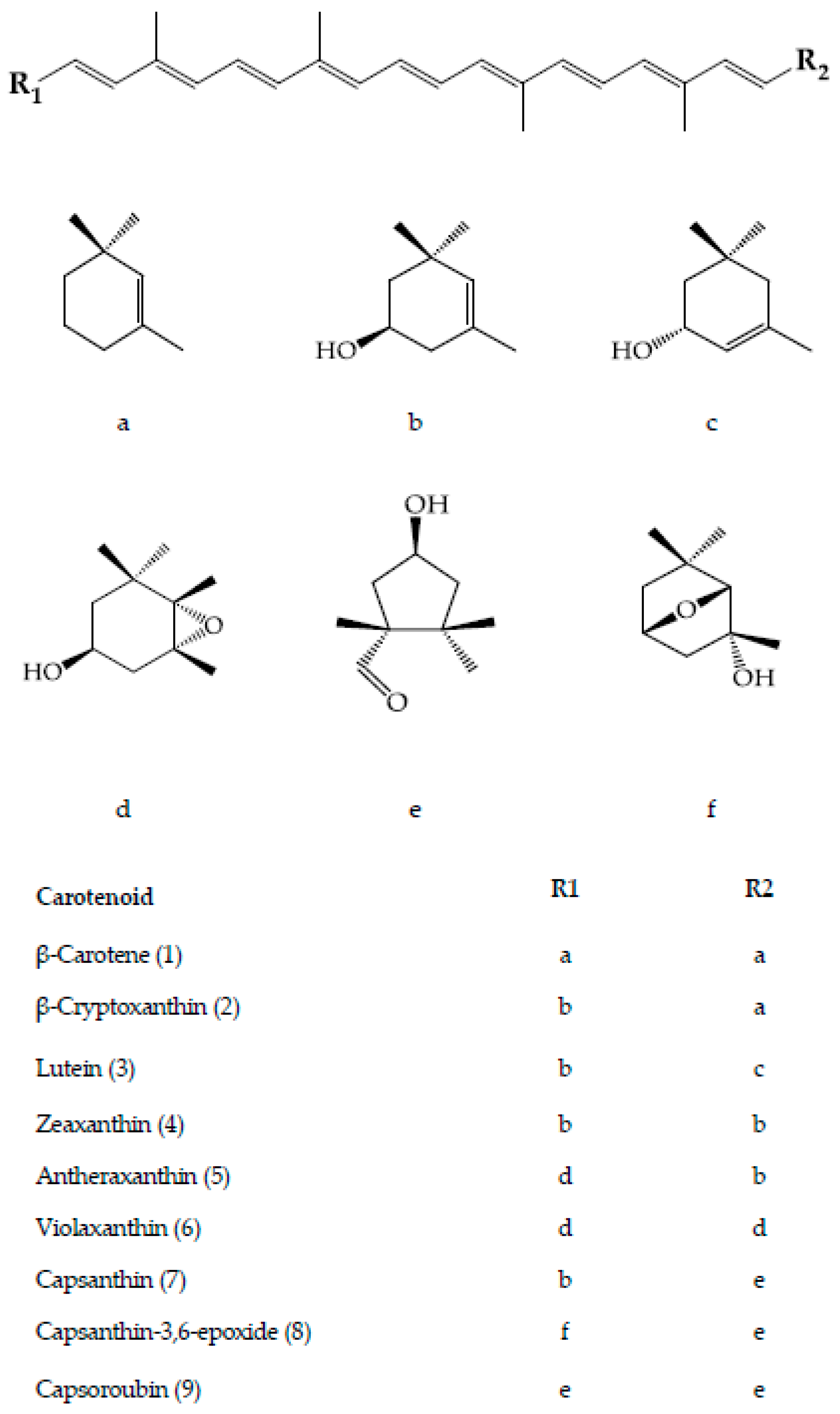
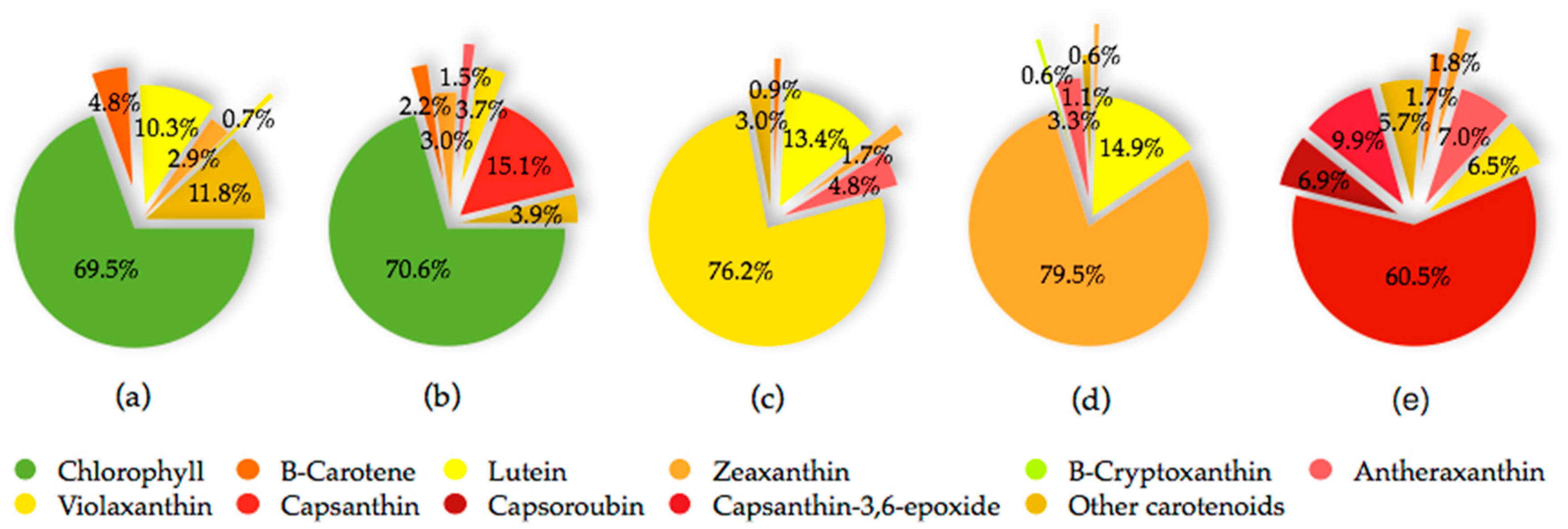
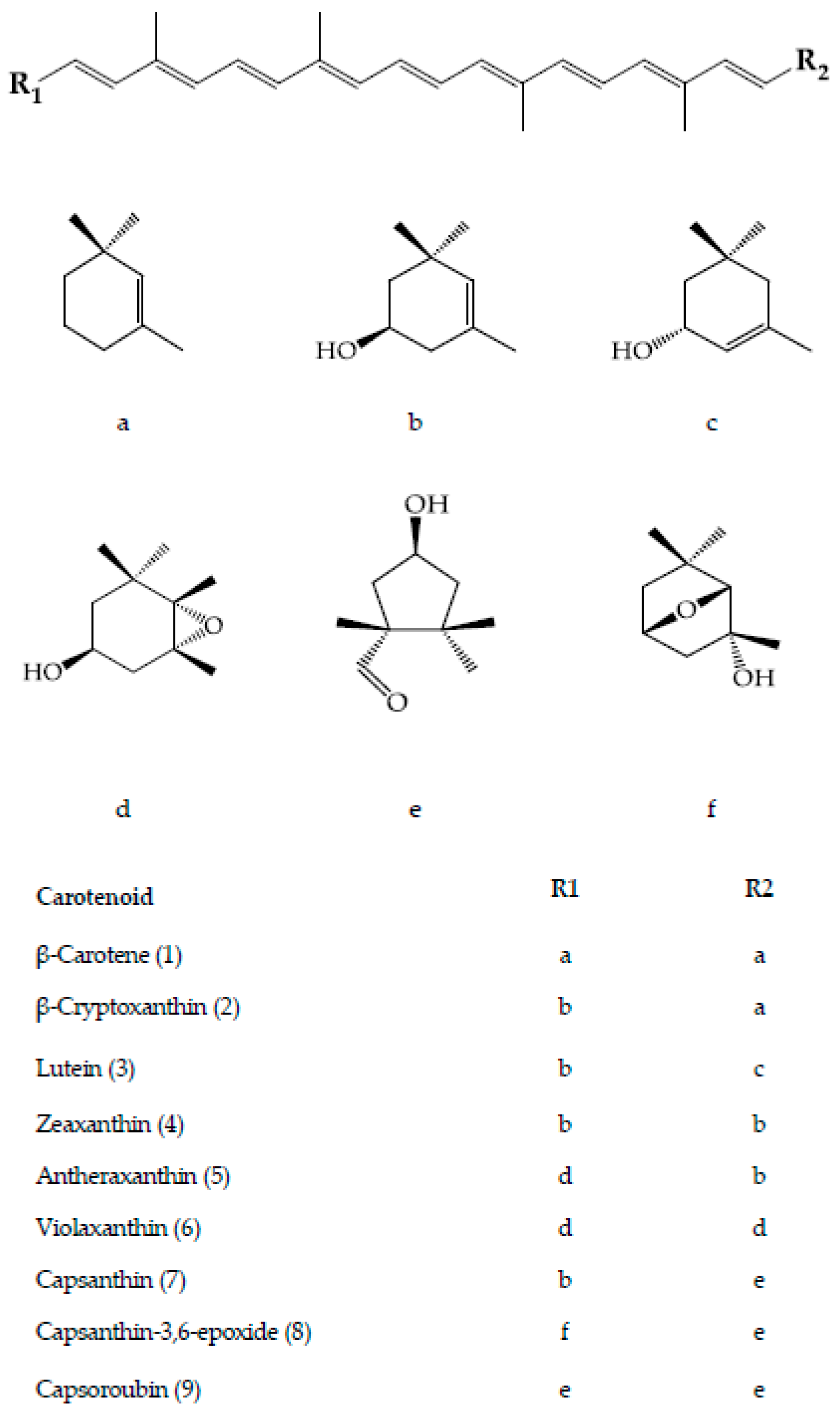
3. Carotenoids of Capsicum Species
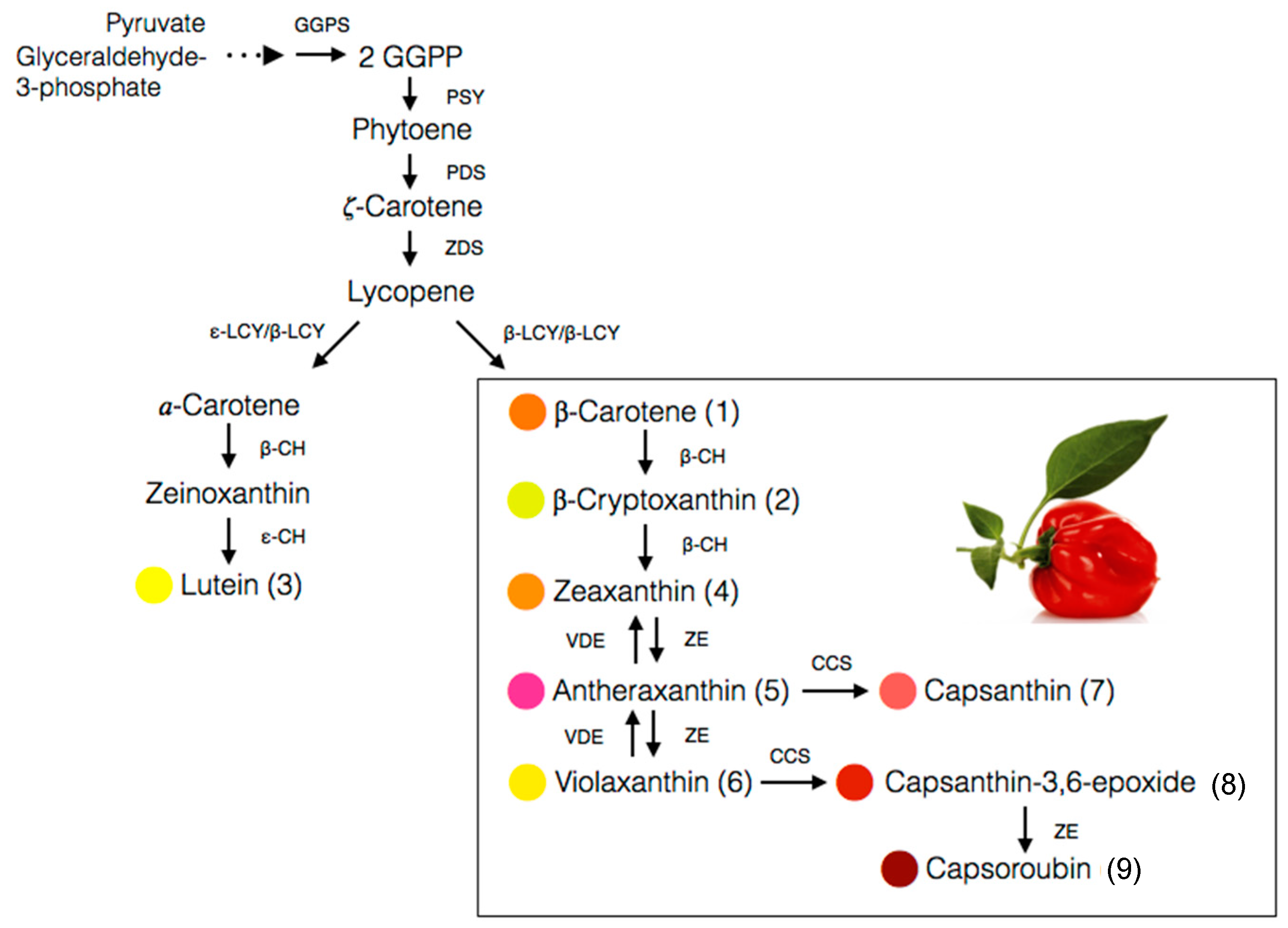
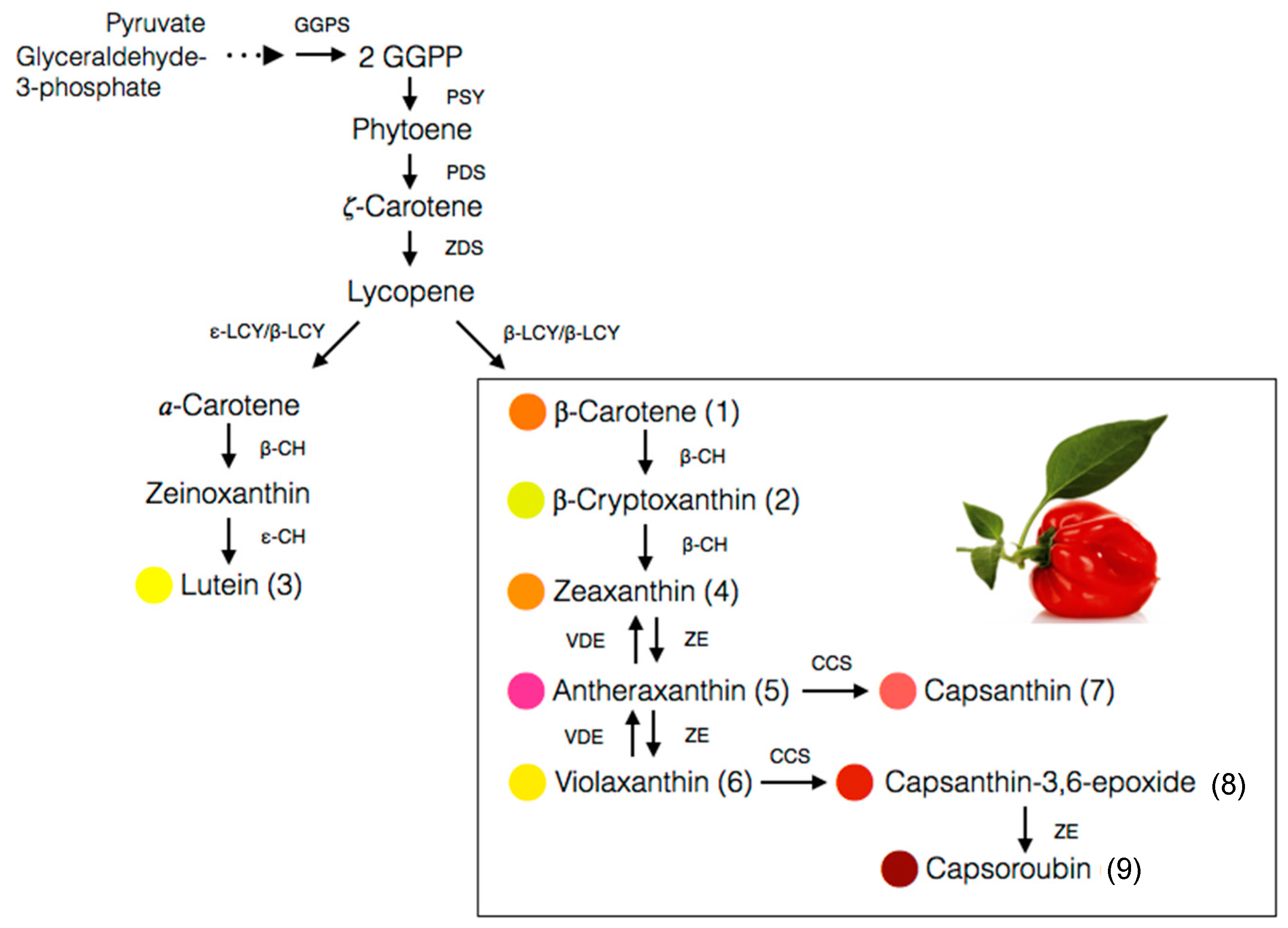
4. Biosynthesis of Carotenoids in Capsicum Species
5. Antioxidant Activities of Pepper’s Carotenoids
6. Health-Promoting Functional Attributes
6.1. Antidiabetic Potential
6.2. Antiadipogenic and Anti-Obesity
6.3. Skin Photoprotective
6.4. Macula Pigments
6.5. Antinociceptive/Analgesic and Anti-Inflammatory
6.6. Antihyperlipidemic and Cardioprotective
6.7. Hepatoprotective
6.8. Chemopreventive
6.9. Provitamin A Activity
7. Safety of Capsicum and Its Carotenoids
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pugliese, A.; Loizzo, M.R.; Tundis, R.; O’Callaghan, Y.; Galvin, K.; Menichini, F.; O’Brien, N. The effect of domestic processing on the content and bioaccessibility of carotenoids from chili peppers (Capsicum species). Food Chem. 2013, 141, 2606–2613. [Google Scholar] [CrossRef] [PubMed]
- Ludy, M.J.; Moore, G.E.; Mattes, R.D. The Effects of capsaicin and capsiate on energy balance: Critical review and meta-analyses of studies in humans. Chem. Sens. 2012, 37, 103–121. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ortega, M.; Ortiz-Moreno, A.; Hernández-Navarro, M.D.; Chamorro-Cevallos, G.; Dorantes-Alvarez, L.; Necoechea-Mondragón, H. Antioxidant, antinociceptive, and anti-Inflammatory effects of carotenoids extracted from dried pepper (Capsicum annuum L.). J. Biomed. Biotechnol. 2012, 2012, 524019. [Google Scholar] [CrossRef] [PubMed]
- Pugliese, A.; O’Callaghan, Y.; Tundis, R.; Galvin, K.; Menichini, F.; O’Brien, N.; Loizzo, M.R. In vitro investigation of the bioaccessibility of carotenoids from raw, frozen and boiled red chili peppers (Capsicum annuum). Eur. J. Nutr. 2014, 53, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Eroglu, A.; Harrison, E.H. Carotenoid metabolism in mammals, including man: Formation, occurrence, and function of apocarotenoids. J. Lipid Res. 2013, 54, 1719–1730. [Google Scholar] [CrossRef] [PubMed]
- Westphal, A.; Jena, V.B. Carotenoids: Properties, distribution, bioavailability, metabolism and health effects. Ernaehrungs Umschau Int. 2015, 11, 196–207. [Google Scholar] [CrossRef]
- Bhandari, S.R.; Jung, B.-D.; Baek, H.-Y.; Lee, Y.-S. Ripening-dependent changes in phytonutrients and antioxidant activity of red pepper (Capsicum annuum L.) fruits cultivated under open-field conditions. Hortic. Sci. 2013, 48, 1275–1282. [Google Scholar] [CrossRef]
- Zimmer, A.R.; Leonardi, B.; Miron, D.; Schapoval, E.; Oliveira, J.R.D.; Gosmann, G. Antioxidant and anti-inflammatory properties of Capsicum baccatum: From traditional use to scientific approach. J. Ethnopharmacol. 2012, 139, 228–233. [Google Scholar] [CrossRef]
- Arimboor, R.; Natarajan, R.B.; Menon, K.R.; Chandrasekhar, L.P.; Vidya Moorkoth, V. Red pepper (Capsicum annuum) carotenoids as a source of natural food colors: Analysis and stability—A review. J. Food Sci. Technol. 2015, 52, 1258–1271. [Google Scholar] [CrossRef]
- Chengaiah, B.; Rao, K.M.; Kumar, K.M.; Alagusundaram, M.; Chetty, C.M. Medicinal importance of natural dyes-a review. Int. J. PharmTech. Res. 2010, 2, 144–154. [Google Scholar]
- Nadeem, M.; Muhammad Anjum, F.; Rafiq Khan, M.; Saeed, M.; Riaz, A. Antioxidant potential of bell pepper (Capsicum annuum L.)—A review. Pak. J. Food Sci. 2011, 21, 45–51. [Google Scholar]
- Chen, Y.-H.; Zou, X.H.; Zheng, T.-Z.; Zhou, Q.; Qiu, H.; Chen, Y.-L.; He, M.; Du, J.; Lei, H.K.; Zhou, P. High spicy food intake and risk of cancer: A meta-analysis of case–control studies. Chin. Med. J. 2017, 130, 2241–2250. [Google Scholar] [CrossRef] [PubMed]
- Bosland, P.W.; Votava, E.J. Peppers: Vegetable and Spice Capsicum, 2nd ed.; CABI Publishing: Wallingford, UK, 2012; p. 248. ISBN 9781845938253. [Google Scholar]
- van Zonneveld, M.; Ramirez, M.; Williams, D.E.; Petz, M.; Meckelmann, S.; Avila, T.; Bejarano, C.; Ríos, L.; Peña, K.; Jäger, M.; et al. Screening genetic resources of Capsicum peppers in their primary center of diversity in Bolivia and Peru. PLoS ONE 2015, 10, e0134663. [Google Scholar] [CrossRef] [PubMed]
- Kraft, K.H.; Brown, C.H.; Nabhan, G.P.; Luedeling, E.; Ruiz, J.J.L.; d’Eeckenbrugge, G.C.; Hijmans, R.J.; Gepts, P. Multiple lines of evidence for the origin of domesticated chili pepper, Capsicum annuum, in Mexico. Proc. Natl. Acad. Sci. USA 2014, 111, 6165–6170. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Melendez, A.; Morrell, P.L.; Roose, M.L.; Kim, S.-C. Genetic diversity and structure of semiwild domesticated chiles (Capsicum annuum; Solanaceae) from Mexico. Am. J. Bot. 2009, 96, 1190–1202. [Google Scholar] [CrossRef] [PubMed]
- Sudré, C.P.; Gonçalves, L.S.; Rodrigues, R.; Do Amaral, T.; Riva-Souza, E.M.; Bento, C.D.S. Genetic variability in domesticated Capsicum spp as assessed by morphological and agronomic data in mixed statistical analysis. Genet. Mol. Res. 2010, 9, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Mori, M.; Ishikawa, K.; Takizawa, K.; Nunomura, O. Carotenoids composition in mature Capsicum annuum. Food Sci. Technol. Res. 2007, 17, 77–80. [Google Scholar] [CrossRef]
- Rodriguez-Uribe, L.; Hernandez, L.; Kilcrease, J.P.; Walker, S.; O’Connell, M.A. Capsaicinoid and carotenoid composition and genetic diversity of Kas I and Ccs in New Mexican Capsicum annuum landraces. Hortic. Sci. 2014, 49, 1370–1375. [Google Scholar] [CrossRef]
- Koeda, S.; Sato, K.; Tomi, K.; Tanaka, Y.; Takisawa, R.; Hosokawa, M.; Doi, M.; Nakazaki, T.; Kitajima, A. Analysis of non-pungency, aroma, and origin of a Capsicum chinense cultivar from a Caribbean Island. J. Jpn. Soc. Hortic. Sci. 2014, 83, 244–251. [Google Scholar] [CrossRef]
- Bosland, P.W.; Coon, D.; Reeves, G. ‘Trinidad moruga scorpion’ pepper is the world’s hottest measured chile pepper at more than two million scoville heat units. Hortic. Technol. 2012, 22, 534–538. [Google Scholar] [CrossRef]
- Vilarinho, L.B.B.; da Silva, D.J.H.; Greene, A.; Salazar, K.D.; Alves, C.; Eveleth, M.; Nichols, B.; Tehseen, S.; Khoury, J.K., Jr.; Johnson, J.V.; et al. Inheritance of fruit traits in Capsicum annuum: Heirloom cultivars as sources of quality parameters relating to pericarp shape, color, thickness, and total soluble solids. J. Am. Hortic. Sci. Res. 2015, 140, 597–604. [Google Scholar] [CrossRef]
- Velázquez-Ventura, J.C.; Márquez-Quiroz, C.; de la Cruz-Lázaro, E.; Osorio-Osorio, R.; Preciado-Rangel, P. Morphological variation of wild peppers (Capsicum spp.) from the state of Tabasco, Mexico. Emir. J. Food Agric. 2018, 30, 115–121. [Google Scholar] [CrossRef]
- The Biochemistry of Peppers. Available online: www.chemistryviews.org/details/ezine/6108461/The_Biochemistry_of_Peppers.html (accessed on 26 November 2018).
- Deli, J.; Molnár, P.; Matus, Z.; Tóth, G. Carotenoid composition in the fruits of red paprika (Capsicum annuum var. lycopersiciforme rubrum) during ripening; biosynthesis of carotenoids in red paprika. J. Agric. Food Chem. 2001, 49, 1517–1523. [Google Scholar] [CrossRef] [PubMed]
- Kiokias, S.; Proestos, C.; Varzakas, T. A review of structure, biosynthesis, absorption of carotenoids-analysis and properties of their common natural extracts. Curr. Res. Nutr. Food Sci. 2016, 4, 25–37. [Google Scholar] [CrossRef]
- Egea, I.; Barsan, C.; Bian, W.; Purgatto, E.; Latché, A.; Chervin, C.; Bouzayen, M.; Pech, J.-C. Chromoplast differentiation: Current status and perspectives. Plant Cell Physiol. 2010, 51, 1601–1611. [Google Scholar] [CrossRef] [PubMed]
- Gómez-García, M.R.; Ochoa-Alejo, N. Biochemistry and molecular biology of carotenoid biosynthesis in chili peppers (Capsicum spp.). Int. J. Mol. Sci. 2013, 14, 19025–19053. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.H.; Kim, J.B.; Park, J.S.; Lee, S.W.; Cho, K.J. A comparison of the carotenoid accumulation in Capsicum varieties that show different ripening colours: Deletion of the capsanthin-capsorubin synthase gene is not a prerequisite for the formation of a yellow pepper. J. Exp. Bot. 2007, 58, 3135–3144. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Burruezo, A.; Gonzalez-Mas Mdel, C.; Nuez, F. Carotenoid composition and vitamin A value in aji (Capsicum baccatum L.) and rocoto (C. pubescens R. and P.), 2 pepper species from the Andean region. J. Food Sci. 2010, 75, S446–S453. [Google Scholar] [CrossRef]
- Cervantes-Paz, B.; Yahia, E.M.; Ornelas-Paz, J.J.; Victoria-Campos, C.I.; Ibarra-Junquera, V.; Pérez-Martínez, J.D.; Escalante-Minakata, P. Antioxidant activity and content of chlorophylls and carotenoids in raw and heat-processed Jalapeño peppers at intermediate stages of ripening. Food Chem. 2014, 146, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-S.; An, C.G.; Park, J.-S.; Lim, Y.P.; Kim, S. Carotenoid profiling from 27 types of paprika (Capsicum annuum L.) with different colors, shapes, and cultivation methods. Food Chem. 2016, 201, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Wall, M.M.; Waddell, C.A.; Bosland, P.W. Variation in β-carotene and total carotenoid content in fruits of Capsicum. Hortic. Sci. 2001, 36, 746–749. [Google Scholar] [CrossRef]
- Umigai, N.; Murakami, K.; Shimizu, R.; Takeda, R.; Azuma, T. Safety evaluation and plasma carotenoid accumulation in healthy adult subjects after 12 weeks of paprika oleoresin supplementation. J. Oleo Sci. 2018, 67, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Schweiggert, U.; Kammerer, D.R.; Carle, R.; Schieber, A. Characterization of carotenoids and carotenoid esters in red pepper pods (Capsicum annuum L.) by high-performance liquid chromatography/atmospheric pressure chemical ionization mass spectrometry. Rapid Commun. Mass Spectrom. 2005, 19, 2617–2628. [Google Scholar] [CrossRef] [PubMed]
- Nishino, A.; Ichihara, T.; Takaha, T.; Kuriki, T.; Nihei, H.; Kawamoto, K.; Yasui, H.; Maoka, T. Accumulation of paprika carotenoids in human plasma and erythrocytes. J. Oleo Sci. 2015, 64, 1135–1142. [Google Scholar] [CrossRef]
- Giuffrida, D.; Dugo, P.; Torre, G.; Bignardi, C.; Cavazza, A.; Corradini, C.; Dugo, G. Characterization of 12 Capsicum varieties by evaluation of their carotenoid profile and pungency determination. Food Chem. 2013, 140, 794–802. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Uribe, L.; Guzman, I.; Rajapakse, W.; Richins, R.D.; O’Connell, M.A. Carotenoid accumulation in orange-pigmented Capsicum annuum fruit, regulated at multiple levels. J. Exp. Bot. 2012, 63, 517–526. [Google Scholar] [CrossRef]
- Muhammad Shah, S.N.; Tian, S.-L.; Gong, Z.-H.; Mohamed Hamid, A. Studies on metabolism of capsanthin and its regulation under different conditions in pepper fruits (Capsicum spp.). Ann. Res. Rev. Biol. 2014, 4, 1106–1120. [Google Scholar] [CrossRef]
- Pugliese, A.; O’Callaghan, Y.; Tundis, R.; Galvin, K.; Menichini, F.; O’Brien, N.; Loizzo, M.R. In vitro assessment of the bioaccessibility of carotenoids from sun-dried chilli peppers. Plant Foods Hum. Nutr. 2014, 69, 8–17. [Google Scholar] [CrossRef]
- Topuz, A.; Dincer, C.; Özdemir, K.S.; Feng, H.; Kushad, M. Influence of different drying methods on carotenoids and capsaicinoids of paprika (Cv., Jalapeno). Food Chem. 2011, 129, 860–865. [Google Scholar] [CrossRef] [PubMed]
- Guzman, I.; Hamby, S.; Romero, J.; Bosland, P.W.; O’Connell, M.A. Variability of carotenoid biosynthesis in orange colored Capsicum spp. Plant Sci. 2010, 179, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Acunha, T.D.S.; Crizel, R.L.; Tavares, I.B.; Barbieri, R.L.; de Pereira, C.M.P.; Cesar Valmor Rombaldi, C.V.; Chaves, F.C. Bioactive compound variability in a Brazilian Capsicum pepper collection. Crop Sci. 2017, 57, 1–13. [Google Scholar] [CrossRef]
- Carvalho, A.V.; de Andrade Mattietto, R.; de Oliveira Rios, A.; de Almeida Maciel, R.; Moresco, S.K.; de Souza Oliveira, T.C. Bioactive compounds and antioxidant activity of pepper (Capsicum sp.) genotypes. J. Food Sci. Technol. 2015, 52, 7457–7464. [Google Scholar] [CrossRef]
- Chávez-Mendoza, C.; Sanchez, E.; Muñoz-Marquez, E.; Sida-Arreola, J.P.; Flores-Cordova, M.A. Bioactive compounds and antioxidant activity in different grafted varieties of bell pepper. Antioxidants 2015, 4, 427–446. [Google Scholar] [CrossRef]
- Vinković, T.; Gluščić, V.; Mendaš, G.; Vinković, V.I.; Parađiković, N.; Tkalec, M.; Štolfa Camagajevac, S.I. Phytochemical composition of ground paprika from Eastern Danube region. Agriculture 2018, 24, 3–12. [Google Scholar] [CrossRef]
- Keyhaninejad, N.; Richins, R.D.; O’Connell, M.A. Carotenoid content in field-grown versus greenhouse-grown peppers: Different responses in leaf and fruit. Hort. Sci. 2012, 47, 852–855. [Google Scholar] [CrossRef]
- Zhang, Y.; Navarro, E.; Cánovas-Márquez, J.T.; Almagro, L.; Chen, H.; Chen, Y.Q.; Zhang, H.; Torres-Martínez, S.; Chen, W.; Garre, V. A new regulatory mechanism controlling carotenogenesis in the fungus Mucor circinelloides as a target to generate β-carotene over-producing strains by genetic engineering. Microb. Cell Fact. 2016, 15, 99. [Google Scholar] [CrossRef] [PubMed]
- Perucka, I.; Materska, M. Antioxidant vitamin contents of Capsicum annuum fruit extracts as affected by processing and varietal factors. Acta Sci. Pol. Technol. Aliment 2007, 6, 67–74. [Google Scholar]
- Menichini, F.; Tundis, R.; Bonesi, M.; Loizzo, M.R.; Conforti, F.; Statti, G.; de Cindio, B.; Houghton, P.J.; Menichini, F. The influence of fruit ripening on the phytochemical content and biological activity of Capsicum chinense Jacq. cv Habanero. Food Chem. 2009, 114, 553–560. [Google Scholar] [CrossRef]
- Bunea, A.; Socaciu, C.; Pintea, A. Xanthophyll esters in fruits and vegetables. Not. Bot. Hortic. Agrobot. 2014, 42, 310–324. [Google Scholar] [CrossRef]
- Khoo, H.; Prasad, K.N.; Kong, K.; Jiang, Y.; Ismail, A. Carotenoids and their isomers: Color pigments in fruits and vegetables. Molecules 2011, 16, 1710–1738. [Google Scholar] [CrossRef] [PubMed]
- Giuffrida, D.; Zoccali, B.; Giofrè, S.V.; Dugo, P.; Mondello, L. Apocarotenoids determination in Capsicum chinense Jacq. cv. Habanero, by supercritical fluid chromatography-triple-quadrupole/mass spectrometry. Food Chem. 2017, 231, 316–323. [Google Scholar] [CrossRef]
- Maoka, T.; Fujiwara, Y.; Hashimoto, K.; Akimoto, N. Isolation of a series of apocarotenoids from the fruits of the red paprika Capsicum annuum L. J. Agric. Food Chem. 2001, 49, 1601–1606. [Google Scholar] [CrossRef]
- Maoka, T.; Akimoto, T.N.; Fujiwara, Y.; Hashimoto, K. Structure of new carotenoids with the 6-oxo-κ end group from the fruits of paprika, Capsicum annuum. J. Nat. Prod. 2004, 67, 115–117. [Google Scholar] [CrossRef]
- Abdel-Aal, E.M.; Akhtar, H.; Zaheer, K.; Ali, R. Dietary sources of lutein and zeaxanthin carotenoids and their role in eye health. Nutrients 2013, 5, 1169–1185. [Google Scholar] [CrossRef]
- Jahns, P.; Holzwarth, A.R. The role of the xanthophyll cycle and of lutein in photoprotection of photosystem II. Biochim. Biophys. Acta 2012, 1817, 182–193. [Google Scholar] [CrossRef] [Green Version]
- Murillo, E.; Nagy, V.; Agócs, A.; Deli, J. Carotenoids with κ-end group. In Carotenoids: Food Sources, Production and Health Benefits; Yamaguchi, M., Ed.; Nova Science Publishers Inc.: New York, NY, USA, 2013; pp. 49–77. ISBN 978-1-62808-622-5. [Google Scholar]
- Tian, S.; Li, L.; Shah, S.N.M.; Gong, Z. The relationship between red fruit colour formation and key genes of capsanthin biosynthesis pathway in Capsicum annuum. Biol. Plant. 2015, 59, 507–513. [Google Scholar] [CrossRef]
- Mercy, E.R.; David, U. Potential health benefits of conventional nutrients and phytochemicals of Capsicum peppers. Pharm. Pharmacol. Int. J. 2018, 6, 62–69. [Google Scholar] [CrossRef]
- Abbeddou, S.; Petrakis, C.; Pérez-Galvez, A.; Kefalas, P.; Hornero-Mendez, D. Effect of simulated thermo-degradation on the carotenoids, tocopherols and antioxidant properties of tomato and paprika oleoresins. J. Oil Chem. Soc. 2013, 90, 1697–1703. [Google Scholar] [CrossRef]
- González-Ponce, H.A.; Rincón-Sánchez, A.R.; Jaramillo-Juárez, F.; Moshage, H. Natural dietary pigments: Potential mediators against hepatic damage induced by over-the-counter non-steroidal anti-inflammatory and analgesic drugs. Nutrients 2018, 10, 117. [Google Scholar] [CrossRef]
- Cervantes-Paz, B.; Yahia, E.M.; Ornelas-Paz, J.d.J.; Gardea-Béjar, A.A.; Ibarra-Junquera, V.; Pérez-Martínez, J.D. Effect of heat processing on the profile of pigments and antioxidant capacity of green and red jalapeño peppers. J. Agric. Food Chem. 2012, 60, 10822–10833. [Google Scholar] [CrossRef]
- Bae, H.; Jayaprakasha, G.K.; Jifon, J.; Patil, B.S. Variation of antioxidant activity and the levels of bioactive compounds in lipophilic and hydrophilic extracts from hot pepper (Capsicum spp.) cultivars. Food Chem. 2012, 134, 1912–1918. [Google Scholar] [CrossRef]
- Le Grandois, J.; Guffond, D.; Hamon, E.; Marchioni, E.; Werner, D. Combined microplate-ABTS and HPLC-ABTS analysis of tomato and pepper extracts reveals synergetic and antagonist effects of their lipophilic antioxidative components. Food Chem. 2007, 223, 62–71. [Google Scholar] [CrossRef]
- Chuah, A.M.; Lee, Y.-C.; Yamaguchi, T.; Takamura, H.; Yin, L.-J.; Matoba, T. Effect of cooking on the antioxidant properties of coloured peppers. Food Chem. 2008, 111, 20–28. [Google Scholar] [CrossRef]
- Ornelas-Paz, J.D.J.; Cira-Chávez, L.A.; Gardea-Béjar, A.A.; Guevara-Arauza, J.C.; Ahumada, D.R.S.; Reyes-Hernández, J.; Ruiz-Cruz, S. Effect of heat treatment on the content of some bioactive compounds and free radical-scavenging activity in pungent and non-pungent peppers. J. Food Res. Int. 2013, 50, 519–525. [Google Scholar] [CrossRef]
- Sayin, K.; Arslan, D. Antioxidant properties, ascorbic acid and total carotenoid value of sweet and hot red pepper paste: A traditional food in Turkish diet. Int. J. Nutr. Food Eng. 2015, 9, 834–837. [Google Scholar] [CrossRef]
- Blanco-Ríos, A.K.; Medina-Juárez, L.A.; González-Aguilar, G.A.; Gámez-Meza, N. Antioxidant activity of the phenolic and oily fractions of different sweet bell peppers. J. Mex. Chem. Soc. 2013, 57, 137–143. [Google Scholar] [CrossRef]
- Abourashed, E.A. Bioavailability of plant-derived antioxidants. Antioxidants 2013, 2, 309–325. [Google Scholar] [CrossRef]
- Pérez-Gálvez, A.; Mínguez-Mosquera, M.I. Structure-reactivity relationship in the oxidation of carotenoid pigments of the pepper (Capsicum annuum L.). J. Agric. Food Chem. 2001, 49, 4864–4869. [Google Scholar] [CrossRef]
- Maoka, T.; Goto, Y.; Isobe, K.; Fujiwara, Y.; Hashimoto, K.; Mochida, K. Antioxidative activity of capsorubin and related compounds from paprika (Capsicum annuum). J. Oleo Sci. 2001, 50, 663–665. [Google Scholar] [CrossRef]
- Pérez-Gálvez, A.; Mínguez-Mosquera, M.I. Degradation of non-esterified and esterified xanthophylls by free radicals. Biochim. Biophys. 2002, 1569, 31–34. [Google Scholar] [CrossRef]
- Lv, J.; Qi, L.; Yu, C.; Yang, L.; Guo, Y.; Chen, Y.; Bian, Z.; Sun, D.; Du, J.; Ge, P.; et al. Consumption of spicy foods and total and cause specific mortality: Population based cohort study. BMJ 2015, 351, 3942. [Google Scholar] [CrossRef]
- Chopan, M.; Littenberg, B. The association of hot red chili pepper consumption and mortality: A large population-based cohort study. PLoS ONE 2017. [Google Scholar] [CrossRef]
- Fiedor, J.; Burda, K. Potential role of carotenoids as antioxidants in human health and disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef]
- Kim, S.; Ha, T.Y.; Hwang, I.K. Analysis, bioavailability, and potential healthy effects of capsanthin, natural red pigment from Capsicum spp. Food Rev. Int. 2009, 25, 198–213. [Google Scholar] [CrossRef]
- Perera, C.O.; Yen, G.M. Functional properties of carotenoids in human health. Int. J. Food Prop. 2007, 10, 201–230. [Google Scholar] [CrossRef]
- Tundis, R.; Loizzo, M.R.; Menichini, F.; Bonesi, M.; Conforti, F.; Statti, G.; De Luca, D.; de Cindio, B.; Menichini, F. Comparative study on the chemical composition, antioxidant properties and hypoglycaemic activities of two Capsicum annuum L. cultivars (acuminatum small and cerasiferum). Plant Foods Hum. Nutr. 2011, 66, 261–269. [Google Scholar] [CrossRef]
- Tundis, R.; Loizzo, M.R.; Menichini, F.; Bonesi, M.; Conforti, F.; De Luca, D.; Menichini, F. Air-dried Capsicum annuum var. acuminatum medium and big: Determination of bioactive constituents, antioxidant activity and carbohydrate-hydrolyzing enzymes inhibition. Food Res. Int. 2012, 45, 170–176. [Google Scholar] [CrossRef]
- Tundis, R.; Menichini, F.; Bonesi, M.; Conforti, F.; Statti, G.; Menichini, F.; Loizzo, M.R. Antioxidant and hypoglycaemic activities and their relationship to phytochemicals in Capsicum annuum cultivars during fruit development. LWT Food Sci. Technol. 2013, 53, 370–377. [Google Scholar] [CrossRef]
- Maeda, H.; Saito, S.; Nakamura, N.; Maoka, T. Paprika pigments attenuate obesity-induced inflammation in 3T3-L1 adipocytes. ISRN Inflamm. 2013. [Google Scholar] [CrossRef]
- Jo, S.J.; Kim, J.W.; Choi, H.O.; Kim, J.H.; Kim, H.J.; Woo, S.H.; Han, B.H. Capsanthin inhibits both adipogenesis in 3T3-L1 preadipocytes and weight gain in high-fat diet-induced obese mice. Biomol. Ther. 2017, 25, 329–336. [Google Scholar] [CrossRef]
- Kou, L.; Du, M.; Zhang, C.; Dai, Z.; Li, X.; Zhang, B. The hypoglycemic, hypolipidemic and anti-diabetic nephritic activities of zeaxanthin in diet-streptozotocin-induced diabetic Sprague Dawley rats. Appl. Biochem. Biotechnol. 2017, 182, 944–955. [Google Scholar] [CrossRef] [PubMed]
- Fernández-García, E.; Carvajal-Lérida, I.; Pérez-Gálvez, A. Carotenoids exclusively synthesized in red pepper (capsanthin and capsorubin) protect human dermal fibroblasts against UVB induced DNA damage. Photochem. Photobiol. Sci. 2016, 15, 1204–1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishino, A.; Sugimoto, K.; Sambe, H.; Ichihara, T.; Takaha, T.; Kuriki, T. Effects of dietary paprika xanthophylls on ultraviolet light-induced skin damage: A double-blind placebo-controlled study. J. Oleo Sci. 2018, 67, 863–869. [Google Scholar] [CrossRef] [PubMed]
- Boiko, Y.A.; Kravchenko, I.A.; Shandra, A.A.; Boiko, I.A. Extraction, identification and anti-inflammatory activity of carotenoids out of Capsicum Anuum L. J. Herbmed. Pharmacol. 2017, 6, 10–15. [Google Scholar]
- Kwon, M.-J.; Song, Y.-S.; Choi, M.-S.; Song, Y.-K. Red pepper attenuates cholesteryl ester transfer protein activity and atherosclerosis in cholesterol-fed rabbits. Clin. Chim. Acta 2003, 332, 37–44. [Google Scholar] [CrossRef]
- Aizawa, K.; Inakuma, T. Dietary capsanthin, the main carotenoid in paprika (Capsicum annuum), alters plasma high-density lipoprotein-cholesterol levels and hepatic gene expression in rats. Br. J. Nutr. 2009, 102, 1760–1766. [Google Scholar] [CrossRef]
- Priya, A.S.; Anitha, A. In vivo hepatoprotective effect of ethanolic extract of Capsicum annuum L. red variety against paracetamol induced hepatotoxicity in male wistar albino rats. Eur. J. Biomed. Pharm. Sci. 2017, 4, 326–332. [Google Scholar]
- Kim, J.-S.; Lee, W.-M.; Rhee, H.C.; Kim, S. Red paprika (Capsicum annuum L.) and its main carotenoids, capsanthin and β-carotene, prevent hydrogen peroxide-induced inhibition of gap-junction intercellular communication. Chem. Biol. Interact. 2016, 254, 146–155. [Google Scholar] [CrossRef]
- Molnár, J.; Gyémánt, N.; Mucsi, I.; Molnár, A.; Szabó, M.; Körtvélyesi, T.; Varga, A.; Molnár, P.; Tóth, G. Modulation of multidrug resistance and apoptosis of cancer cells by selected carotenoids. In Vivo 2004, 18, 237–244. [Google Scholar]
- Burrows, T.L.; Warren, J.M.; Colyvas, K.; Garg, M.L.; Collins, C.E. Validation of overweight children’s fruit and vegetable intake using plasma carotenoids. Obesity 2009, 17, 162–168. [Google Scholar] [CrossRef]
- Canas, J.A.; Damaso, L.; Altomare, A.; Killen, K.; Hossain, J.; Balagopal, P. Insulin resistance and adiposity in relation to serum b-Carotene levels. J. Paediatr. 2012, 161, 58–64. [Google Scholar] [CrossRef]
- Fernández-García, E. Skin protection against UV light by dietary antioxidants. Food Funct. 2014, 5, 1994–2003. [Google Scholar] [CrossRef] [PubMed]
- Stahl, W.; Sies, H. β-Carotene and other carotenoids in protection from sunlight. Am. J. Clin. Nutr. 2012, 96, 1179S–1184S. [Google Scholar] [CrossRef] [PubMed]
- Palombo, P.; Fabrizi, G.; Ruocco, V.; Ruocco, E.; Fluhr, J.; Roberts, R.; Morganti, P. Beneficial long-term effects of combined oral/topical antioxidant treatment with the carotenoids lutein and zeaxanthin on human skin: A double-blind, placebo-controlled study. Skin Pharmacol. Physiol. 2007, 20, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Yan, S.F.; Huang, Y.M.; Lu, X.R.; Qian, F.; Pang, H.L.; Xu, X.R.; Zou, Z.Y.; Dong, P.C.; Xiao, X.; et al. Effect of lutein and zeaxanthin on macular pigment and visual function in patients with early age-related macular degeneration. Ophthalmology 2012, 119, 2290–2297. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarieh, M.; Sacu, S.; Wedrich, A. The role of the carotenoids, lutein and zeaxanthin, in protecting against age-related macular degeneration: A review based on controversial evidence. Nutr. J. 2003, 2, 20. [Google Scholar] [CrossRef]
- Connolly, E.E.; Beatty, S.; Loughman, J.; Howard, A.N.; Louw, M.S.; Nolan, J.M. Supplementation with all three macular carotenoids: Response, stability, and safety. Investig. Ophthalmol. Vis. Sci. 2011, 52, 9207–9217. [Google Scholar] [CrossRef]
- Wu, J.; Cho, E.; Willett, W.C.; Sastry, S.M.; Schaumberg, D.A. Intakes of lutein, zeaxanthin, and other carotenoids and age-related macular degeneration during 2 decades of prospective follow-up. JAMA Ophthalmol. 2015, 133, 1415–1424. [Google Scholar] [CrossRef]
- Wong, W.L.; Su, X.; Li, X.; Cheung, C.M.G.; Klein, R.; Cheng, C.-Y.; Wong, T.Y. Global prevalence of age-related macular degeneration and disease burden projection for 2020 and 2040: A systematic review and meta-analysis. Lancet Glob. Health 2014. [Google Scholar] [CrossRef]
- Voutilainen, S.; Nurmi, T.; Mursu, J.; Rissanen, T.H. Carotenoids and cardiovascular health. Am. J. Clin. Nutr. 2006, 83, 1265–1271. [Google Scholar] [CrossRef]
- Ciccone, M.M.; Cortese, F.; Gesualdo, M.; Carbonara, S.; Zito, A.; Ricci, G.; De Pascalis, F.; Scicchitano, P.; Riccioni, G. Dietary intake of carotenoids and their antioxidant and anti-inflammatory effects in cardiovascular care. Mediat. Inflamm. 2013. [Google Scholar] [CrossRef]
- Di Pietro, N.; Di Tomo, P.; Pandolfi, A. Carotenoids in cardiovascular disease prevention. JSM Atheroscler. 2016, 1, 1002. [Google Scholar]
- Karathanasis, S.K.; Freeman, L.A.; Gordon, S.M.; Remaley, A.T. The changing face of HDL and the best way to measure it. Clin. Chem. 2017, 63, 196–210. [Google Scholar] [CrossRef]
- Murillo, A.G.; DiMarco, D.M.; Luz Fernandez, M. The potential of non-provitamin A carotenoids for the prevention and treatment of non-alcoholic fatty liver disease. Biology 2016, 5, 42. [Google Scholar] [CrossRef]
- Ni, Y.; Zhuge, F.; Nagashimada, M.; Ota, T. Novel action of carotenoids on non-alcoholic fatty liver disease: Macrophage polarization and liver homeostasis. Nutrients 2016, 8, 391. [Google Scholar] [CrossRef]
- Cao, Y.; Wang, C.; Liu, J.; Liu, Z.; Ling, W.; Chen, Y. Greater serum carotenoid levels associated with lower prevalence of nonalcoholic fatty liver disease in Chinese adults. Sci. Rep. 2015, 5, 12951. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, B.; Sahin, K.; Bilen, H.; Bahcecioglu, I.H.; Bilir, B.; Ashraf, S.; Halazun, K.J.; Kucuk, O. Carotenoids and non-alcoholic fatty liver disease. Hepatobiliary Surg. Nutr. 2015, 4, 161–171. [Google Scholar] [CrossRef]
- Bhattacharya, S. Anticarcinogenic property of medicinal plants: Involvement of antioxidant role. In Medicinal Plants as Antioxidant Agents: Understanding Their Mechanism of Action and Therapeutic Efficacy; Capasso, A., Ed.; Research Signpost: Kerala, India, 2012; pp. 83–96. ISBN 978-81-308-0509-2. [Google Scholar]
- Tanaka, T.; Shnimizu, M.; Moriwaki, H. Cancer chemoprevention by carotenoids. Molecules 2012, 17, 3202–3242. [Google Scholar] [CrossRef]
- Nishino, H.; Murakoshi, M.; Ii, T.; Takemura, M.; Kuchide, M.; Kanazawa, M.; Mou, X.Y.; Wada, S.; Masuda, M.; Ohsaka, Y.; et al. Carotenoids in cancer chemoprevention. Cancer Metastasis Rev. 2002, 21, 257–264. [Google Scholar] [CrossRef]
- De Flora, S.; Ferguson, L.R. Overview of mechanisms of cancer chemopreventive agents. Mutat. Res. 2005, 591, 8–15. [Google Scholar] [CrossRef]
- Maoka, T.; Mochida, K.; Kozuka, M.; Ito, Y.; Fujiwara, Y.; Hashimoto, K.; Enjo, F.; Ogata, M.; Nobukuni, Y.; Tokuda, H.; et al. Cancer chemopreventive activity of carotenoids in the fruits of red paprika Capsicum annuum L. Cancer Lett. 2001, 172, 103–109. [Google Scholar] [CrossRef]
- Huang, X.; Gao, Y.; Zhi, X.; Ta, N.; Jiang, H.; Zheng, J. Association between vitamin A, retinol and carotenoid intake and pancreatic cancer risk: Evidence from epidemiologic studies. Sci. Rep. 2016. [Google Scholar] [CrossRef]
- Grune, T.; Lietz, G.; Palou, A.; Ross, A.C.; Stahl, W.; Tang, G.; Thurnham, D.; Yin, S.; Biesalski, H.K. β-Carotene as an important vitamin A source for humans. Nutrition 2010, 140, 2268S–2285S. [Google Scholar] [CrossRef]
- Global Prevalence of Vitamin A Deficiency in Populations at Risk 1995–2005. WHO Global Database on Vitamin A Deficiency. 2009. Available online: www.who.int/vmnis/database/vitamina/x/en/ (accessed on 26 November 2018).
- Weber, D.; Grune, T. The contribution of beta-carotene to vitamin A supply of humans. Mol. Nutr. Food Res. 2012, 56, 251–258. [Google Scholar] [CrossRef]
- Tomlekova, N.B.; White, P.J.; Thompson, J.A.; Penchev, E.A.; Nielen, S. Mutation increasing β-carotene concentrations does not adversely affect concentrations of essential mineral elements in pepper fruit. PLoS ONE 2017, 12, e0172180. [Google Scholar] [CrossRef]
- Burri, B.J. Beta-cryptoxanthin as a source of vitamin A. J. Sci. Food Agric. 2015, 95, 1786–1794. [Google Scholar] [CrossRef]
- Marin, A.; Ferreres, F.; Tomas-Barberaan, F.A.; Gil, M.I. Characterization and quantitation of antioxidant constituents of sweet pepper (Capsicum annuum L.). J. Agric. Food Chem. 2004, 52, 3861–3869. [Google Scholar] [CrossRef]
- Blumhoff, R. Vitamin A and carotenoid toxicity. Food Nutr. Bull. 2001, 22, 320–334. [Google Scholar] [CrossRef]
- Vitamin, A.U.S. Department of Health and Human Services. Available online: https://ods.od.nih.gov/factsheets/VitaminA-Consumer/ (accessed on 9 September 2018).
- Parvez, G.M.M. Current advances in pharmacological activity and toxic effects of various Capsicum species. Int. J. Pharm. Sci. Res. 2017, 8, 1900–1912. [Google Scholar] [CrossRef]
- Wahyuni, Y.; Ballester, A.-R.; Sudarmonowati, E.; Bino, R.J.; Bovy, A.G. Metabolite biodiversity in pepper (Capsicum) fruits of thirty-two diverse accessions: Variation in health-related compounds and implications for breeding. Phytochemistry 2011, 72, 1358–1370. [Google Scholar] [CrossRef]
- Saleh, B.K.; Omer, A.; Teweldemedhin, B. Medicinal uses and health benefits of chili pepper (Capsicum spp.): A review. MOJ Food Process Technol. 2018, 6, 325–328. [Google Scholar] [CrossRef]
- EFSA ANS Panel (EFSA Panel on Food Additives and Nutrient Sources added to Food). Scientific Opinion on the re-evaluation of paprika extract (E 160c) as a food additive. EFSA J. 2015, 13, 4320. [Google Scholar] [CrossRef]
- Bendich, A. The safety of β-carotene. Nutr. Cancer 2009, 11, 207–214. [Google Scholar] [CrossRef]
- Toti, E.; Chen, C.O.; Palmery, M.; Valencia, D.B.; Peluso, I. Non-Provitamin A and Provitamin A Carotenoids as Immunomodulators: Recommended Dietary Allowance, Therapeutic Index, or Personalized Nutrition? Oxid. Med. Cell. Longev. 2018, 2018, 4637861. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Cultivar | Colour | Carotenoid Concentration | Reference | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | ||||
| Red Mountain | Red | 31.8 ± 4.54 | 4.6 ± 0.71 | 2.73 ± 0.77 | 0.59 ± 0.16 | ND | 3.61 ± 0.44 | ND | ND | 43.32 ± 6.88 | ||
| Magnifico | Red | 28.33 ± 2.83 | 4.48 ± 0.78 | 2.06 ± 0.06 | 0.4 ± 0.04 | ND | ND | ND | ND | 35.99 ± 3.51 | ||
| Nagano | Red | 22.88 ± 1.65 | 4.01 ± 0.32 | 2.01 ± 0.05 | 0.31 ± 0.02 | ND | ND | ND | ND | 29.82 ± 2.01 | ||
| aC. annuum | Preludium | Red | 29.56 ± 0.67 | 2.67 ± 0.09 | 1.94 ± 0.17 | 0.33 ± 0.02 | ND | ND | ND | ND | 35.05 ± 0.90 | [32] |
| Adami Red | Red | 28.62 ± 0.99 | 2.59 ± 0.17 | 1.88 ± 0.18 | 0.32 ± 0.02 | ND | ND | ND | ND | 33.94 ± 1.38 | ||
| Raon Red | Red | 13.26 ± 5.14 | 1.74 ± 0.82 | 0.83 ± 0.17 | 0.13 ± 0.07 | ND | ND | ND | ND | 13.51 ± 1.51 | ||
| Red | Red | 21.55 ± 5.93 | 3.09 ± 0.97 | 1.13 ± 0.21 | 0.30 ± 0.09 | ND | ND | ND | ND | 29.11 ± 1.52 | ||
| RD-Glory | Red | 3.98 ± 0.47 | 0.95 ± 0.09 | 0.86 ± 0.02 | 0.47 ± 0.24 | ND | 24.05 ± 0.16 | ND | ND | 37.25 ± 0.63 | ||
| Mazzona | Orange | ND | ND | 1.23 ± 0.01 | 1.12 ± 0.04 | 28.39 ± 1.00 | 151.39 ± 4.85 | 6.19 ± 0.09 | 0.29 ± 0.00 | 190.43 ± 5.66 | ||
| Orange Glory | Orange | ND | ND | 0.98 ± 0.09 | 1.09 ± 0.08 | 15.77 ± 1.52 | 145.92 ± 13.17 | 5.52 ± 2.00 | 0.26 ± 0.04 | 171.95 ± 17.13 | ||
| Orange Star | Orange | ND | ND | 0.62 ± 0.05 | 0.64 ± 0.08 | 25.27 ± 3.21 | 140.05 ± 14.48 | 4.47 ± 1.59 | 0.22 ± 0.10 | 172.77 ± 19.71 | ||
| aC. annuum | Raon Orange | Orange | ND | ND | 0.76 ± 0.01 | 0.55 ± 0.01 | 22.24 ± 0.51 | 88.80 ± 1.06 | 0.89 ± 0.07 | ND | 115.01 ± 1.46 | [32] |
| Mini Goggal Orange | Orange | ND | ND | 0.63 ± 0.01 | 0.64 ± 0.01 | 17.32 ± 0.48 | 89.89 ± 2.89 | 1.87 ± 0.77 | ND | 111.83 ± 4.25 | ||
| Orange | Orange | ND | ND | 0.75 ± 0.03 | 0.61 ± 0.01 | 25.10 ± 0.39 | 115.53 ± 1.11 | 3.06 ± 0.32 | ND | 146.93 ± 1.93 | ||
| OE-Glory | Orange | ND | ND | 0.66 ± 0.16 | 0.48 ± 0.11 | 19.31 ± 4.00 | 85.06 ± 19.08 | 2.00 ± 0.49 | 0.28 ± 0.07 | 109.69 ± 24.32 | ||
| Jorrit | Yellow | ND | ND | 0.15 ± 0.04 | 0.03 ± 0.01 | 8.75 ± 2.29 | 0.66 ± 0.31 | 0.81 ± 0.25 | 0.14 ± 0.03 | 11.38 ± 3.14 | ||
| Coletti | Yellow | ND | ND | 0.21 ± 0.01 | 0.05 ± 0.00 | 13.83 ± 0.94 | 0.71 ± 0.06 | 1.48 ± 0.06 | 0.72 ± 0.30 | 18.2 ± 1.46 | ||
| Sven | Yellow | ND | ND | 0.15 ± 0.01 | 0.05 ± 0.01 | 13.16 ± 1.07 | 0.63 ± 0.04 | 1.06 ± 0.20 | 0.20 ± 0.01 | 16.21 ± 1.42 | ||
| aC. annuum | Atalante | Yellow | ND | ND | 0.17 ± 0.00 | 0.03 ± 0.00 | 11.97 ± 0.56 | 0.45 ± 0.09 | ND | 1.05 ± 0.57 | 15.31 ± 1.37 | [32] |
| Raon Yellow | Yellow | ND | ND | 0.30 ± 0.00 | 0.43 ± 0.04 | 21.08 ± 0.84 | 2.22 ± 0.18 | 1.35 ± 0.41 | 2.85 ± 1.34 | 29.70 ± 0.67 | ||
| Yellow | Yellow | ND | ND | 0.29 ± 0.06 | 0.29 ± 0.00 | 18.32 ± 6.79 | 1.27 ± 0.08 | ND | 0.85 ± 0.09 | 22.32 ± 7.23 | ||
| YW-Glory | Yellow | ND | ND | 0.27 ± 0.03 | 0.27 ± 0.07 | 15.17 ± 1.25 | 1.19 ± 0.52 | ND | 0.55 ± 0.21 | 18.45 ± 1.02 | ||
| b,cC. annuum | Pimenta PMO | Red | NA | NA | b 4442.72 ± 1.0 | b111.12 ± 0.19 | b 195.75 ± 0.25 | b 460.03 ± 3.13 | NA | NA | c 1064.35 ± 19.38 | [44] |
| Pimenta Amarela | Yellow | NA | NA | ND | ND | b 312.79 ± 0.12 | b 8.78 ± 0.03 | NA | NA | c91.26 ± 8.59 | ||
| b,cC. baccatum var. umbilicatum | Pimenta Chumbinho Baião | Red | NA | NA | b 454.08 ± 0.20 | b 1456.24 ± 0.80 | b 139.85 ± 0.14 | b 1291.29 ± 0.40 | NA | NA | c 580.98 ± 51.91 | [44] |
| Pimenta Biquinho | Orange | NA | NA | ND | ND | b 687.71 ± 0.66 | b 25.56 ± 0.02 | NA | NA | c 208.45 ± 12.65 | ||
| b,cC. chinense | Pimenta Curuçazinho | Yellow | NA | NA | b 33.48 ± 1.06 | ND | b 89.91 ± 0.65 | b 355.68 ± 0.35 | NA | NA | c 73.80 ± 2.93 | [44] |
| Pimenta Murupi | Yellow | NA | NA | ND | ND | b 262.96 ± 0.08 | b 20.81 ± 5.99 | NA | NA | c 79.56 ± 7.12 | ||
| Functional Property | Type of Study | Biological/Pharmacological/ Clinical Activity | Species/Variety/ Cultivar | Extract/Fraction/ Carotenoid | Carotenoid Content/Purity | Dose | Effects/Identified Mechanism | Reference |
|---|---|---|---|---|---|---|---|---|
| Hypoglycaemic | In vitro | α-Amylase inhibitory | C. chinense cv. Habanero | Lipophilic hexane fraction from ethanol extract of immature (I) and mature (M) fruits. | 62.7 ± 5.5 (I) and 362 ± 7.8 (M) mg β-carotene eq./100 mg FW | IC50, 9.88 (I) and 29.58 (M) µg/mL | A selective α-amylase inhibitory activity. No α-glucosidase inhibitory activity. | [50] |
| C. annuum var. acuminatum small | Lipophilic hexane fraction of ethanol extract from deseeded air-dried mature fruits. | Not determined | IC50, 6.9 µg/mL | A selective α-amylase inhibitory activity. Inactive as α-glucosidase inhibitor. | [79] | |||
| C. annuum var. cerasiferum | Lipophilic hexane fraction of ethanol extract from deseeded air-dried mature fruits. | Not determined | IC50, 20.1 µg/mL | A selective α-amylase inhibitory activity. No α-glucosidase inhibitory activity. | [79] | |||
| C. annuum var. acuminatum big | Lipophilic fraction of ethanol extract from deseeded air-dried immature fruits. | Not determined | IC50, 8.7 µg/mL | A selective α-amylase inhibitory activity. Inactive as α-glucosidase inhibitor. | [80] | |||
| C. annuum cv. Fiesta, Acuminatum, Orange Thai and Cayenne Golden | Lipophilic hexane fraction of ethanol extract from deseeded air-dried immature (I) fruits. | Not determined | IC50, ranged from 9.1 to 28.6 µg/mL (I) | A selective α-amylase inhibitory activity, Fiesta > Cayenne Golden > Acuminatum > Orange Thai. No α-glucosidase inhibitory activity. | [81] | |||
| Anti-obesity | In vitro | Antiadipogenic on murine 3T3-L1 pre-adipocytes | Not determined | Capsanthin purified from commercialised red pepper powder. | Capsanthin, 100% | IC50, 2.5 ± 0.45 µM | The activities resulted from potent adrenoceptor-β2-agonistic which is linked to the activation of hormone sensitive lipase. | [83] |
| Lipolytic in differentiated 3T3-L1 adipocytes | ED50, 0.872 ± 0.06 µM | |||||||
| In vivo | Inhibition of weight gain in high-fat-diet-induced obese female C57BL/6C mice | Not determined | Capsanthin purified from commercialised red pepper powder. | Capsanthin, 100% | 1, 5, and 10 µmol | A dose-dependent enhancement of locomotive activity associated with excessive production of ATP with progressive weight loss. | [83] | |
| Skin photoprotective | In vitro | Anti UVB-induced cytotoxicity on normal human dermal fibroblasts | Not determined | Purified capsanthin, capsorubin and lutein from commercialised paprika oleoresin. | Capsanthin, 100% Capsorubin, 100% Lutein, 100% | 1 µM | Protective effects by producing significant decrease in the formation of UVB-induced DNA strand break and counteracting caspase-3 cleavage. | [85] |
| Clinical | Anti-UV-induced skin damage in a double-blind placebo-controlled study (Japanese male and female, aged 30 to 50 years with skin phototype II). | Not determined | A commercial paprika-xanthophyll preparation (PapriX- oil). | 333 mg of PapriX-oil gelatine capsule (l 9 mg total xanthophylls, 5 mg capsanthin, and 0.5 mg cryptoxanthin) | One capsule orally with a meal every evening for 5 weeks. | Suppression of UV-induced erythema and pigmentation by the xanthophylls’ strong singlet oxygen quenching activity that counteracted UV-induced photooxidative stress and acute inflammation response. | [86] | |
| Anti-Inflammatory | In vitro | Anti-inflammatory in obesity-induced inflammation in 3T3-L1 adipocytes co-cultured with RAW264.7 macrophages. | Not determined | Purified paprika pigments from commercialised paprika carotenoids. | Capsanthin, 44.3% Capsorubin, 12.8% Capsanthin analog | 15, 30, 60 µg/mL | Attenuation of inflammation in the 3T3-L1 adipocytes by dose dependant suppression of adipocytokine mRNA gene expression for IL-6, TNF-α, MCP-1 and resistin, and significant (p < 0.05) deceased versus control in nitric oxide release. | [82] |
| In vivo | Anti-inflammatory in carrageenan-induced mice paw oedema. | C. annuum var. guajillo (Guajilo 15660) | Petroleum ether fraction of acetone extract from dried fruits. | β-carotene, 10.01% β-cryptoxanthin, 11.96% Violaxanthin, 49.06% | 5, 20 and 80 mg/kg | Significant (p < 0.05) reduction of oedema at 5 hr time point, comparable to indomethacine (7 mg/kg) | [3] | |
| Anti-inflammatory in adjuvant-induced mice paw oedema. | C. annuum (Ukrainian cayenne bitter pepper) | Petroleum ether fraction of acetone extract from air-dried fruits. | Carotenoid extract containing 69.3% yellow and 30.7% red fractions rich in capsanthin, lutein and β-carotene. | Topical application of ointment containing 2 mg extract/g daily for 20 days. | Reduction of serum cholinesterase activity by 1.3 times and double decrease in the serum seromucoid concentration that indicated good inhibitory activity. | [87] | ||
| Anti-Hyperlipidemic | In vivo | Inhibition of CETP activity and anti-atherosclerotic in cholesterol-fed male New Zealand white rabbits. | Not determined | Mixture of red pepper powder, Purina lab Chow (Purina Chemical, Korea) and 1% cholesterol. | Red pepper powder, 1% | 100 g supplement/day for 12 weeks | Significant (p < 0.05) lowered in total cholesterol, triglyceride, LDL-C, VLDL-C, and VLDL-TG levels, atherogenic index and CETP activity whereas higher HDL-C level than in the control group during the experimental period. | [88] |
| Promotion of plasma LDL-C levels and hepatic gene expression in young male Wistar rats. | Not determined | Purified capsanthin from non-acylated capsanthin powder. | Capsanthin, 100% | 0·25 and 0·49 mmol/kg diet ad libitum for 2 weeks | A dose-dependent increment of HDL-C associated with up-regulation of mRNA for apoA5 and LCAT. | [89] | ||
| Hepatoprotective | In vivo | Liver protectant against paracetamol induced hepatotoxicity in male albino Wistar rats. | C. annuum | Ethanol extract of dried red fruit powder. | Lycopene, 4.69 ± 0.01 mg/g tissue | 250 and 500 mg/kg body weight | Improvement of liver functions as indicated by significant decreased in liver weight and lipid levels while increased in glycogen and glycoprotein in liver tissue as well as decreased in serum liver markers (AST, ALT ALP) and bilirubin, (p < 0.001) versus untreated rats. | [90] |
| Chemopreventive | In vitro | Protective activity against H2O2-induced inhibition of GJIC in WB-F344 rat liver epithelial cells. | C. annuum | Diethyl ether fraction of the red fruits (RPE), capsanthin (CST) and β-carotene (BCT). | RPE CST, 100% BCT, 100% | RPE, 10 and 25 µg/mL CST, 0.5 and 1.0 µM BCT, 0.1 and 0.5 µM | RPE, CST and BCT prevented GJIC inhibition by blocking the generation and actions of ROS and enhancing Cx43 mRNA expression, protein levels and the activity of ERK1/2, p38 and JNK MAP kinases. | [91] |
| MDR-efflux protein inhibitory in human MDR-1 gene-transfected L1210 mouse lymphoma cells and human breast cancer cells MDA-MB-231 (HTB-26). | C. annuum | Purified capsanthin and capsorubin from red paprika. | Capsanthin, 100% Capsorubin, 100% | 4 µg/mL | Capsanthin and capsorubin enhanced rhodamine 123 accumulation 30-fold relative to nontreated lymphoma cells that suggested their MDR reversal effect on the cells. | [92] | ||
| Apoptosis induction in human breast cancer cells MDA- MB-231 (HTB-26). | C. annuum | Purified capsanthin from red paprika. | Capsanthin, 100% | 2 µg/mL | Capsanthin induced apoptosis in both tumour cells comparable to that of the positive control, M627 (50 µg/mL). | [92] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohd Hassan, N.; Yusof, N.A.; Yahaya, A.F.; Mohd Rozali, N.N.; Othman, R. Carotenoids of Capsicum Fruits: Pigment Profile and Health-Promoting Functional Attributes. Antioxidants 2019, 8, 469. https://doi.org/10.3390/antiox8100469
Mohd Hassan N, Yusof NA, Yahaya AF, Mohd Rozali NN, Othman R. Carotenoids of Capsicum Fruits: Pigment Profile and Health-Promoting Functional Attributes. Antioxidants. 2019; 8(10):469. https://doi.org/10.3390/antiox8100469
Chicago/Turabian StyleMohd Hassan, Norazian, Nurul Asyiqin Yusof, Amirah Fareeza Yahaya, Nurul Nasyitah Mohd Rozali, and Rashidi Othman. 2019. "Carotenoids of Capsicum Fruits: Pigment Profile and Health-Promoting Functional Attributes" Antioxidants 8, no. 10: 469. https://doi.org/10.3390/antiox8100469
APA StyleMohd Hassan, N., Yusof, N. A., Yahaya, A. F., Mohd Rozali, N. N., & Othman, R. (2019). Carotenoids of Capsicum Fruits: Pigment Profile and Health-Promoting Functional Attributes. Antioxidants, 8(10), 469. https://doi.org/10.3390/antiox8100469