Therapeutic Agents with AHR Inhibiting and NRF2 Activating Activity for Managing Chloracne


1
Department of Dermatology, Kyushu University, Maidashi 3-1-1, Higashiku, Fukuoka 812-8582, Japan
2
Research and Clinical Center for Yusho and Dioxin, Kyushu University, Fukuoka 812-8582, Japan
3
Division of Skin Surface Sensing, Kyushu University, Fukuoka 812-8582, Japan
*
Author to whom correspondence should be addressed.
Antioxidants 2018, 7(7), 90; https://doi.org/10.3390/antiox7070090
Submission received: 13 June 2018
/
Revised: 2 July 2018
/
Accepted: 11 July 2018
/
Published: 13 July 2018
(This article belongs to the Special Issue Nrf2 in Dermatological Pathologies)
{kind=link}
Abstract
:Chloracne is the major skin symptom caused by dioxin intoxication. Dioxin activates the aryl hydrocarbon receptor (AHR)–cytochrome p450 1A1 (CYP1A1) system, generates oxidative stress, and induces hyperkeratinization of keratinocytes and sebocytes leading to chloracne. Nuclear factor-erythroid 2-related factor-2 (NRF2) is a master switch that induces the expression of various antioxidative enzymes, such as heme oxygenase-1. Cinnamaldehyde is an antioxidant phytochemical that inhibits AHR–CYP1A1 signaling and activates the NRF2–antioxidative axis. The cinnamaldehyde-containing Kampo herbal medicine Keishibukuryogan is capable of improving chloracne in Yusho patients who are highly contaminated with dioxin. Agents with dual functions in promoting AHR–CYP1A1 inhibition and NRF2 activation may be useful for managing dioxin-related health hazards.
1. Introduction
Health problems associated with environmental pollutants are an important issue. Environmental polycyclic aromatic hydrocarbons such as 2,3,7,8-tetrachlorodibenzo-p-dioxin, polychlorinated dibenzofuran, and benzo(a)pyrene (BaP) are high-affinity ligands for the aryl hydrocarbon receptor (AHR) or dioxin receptor [1,2,3,4]. These chemical compounds strongly activate AHR, generate reactive oxygen species (ROS), and induce the production of inflammatory cytokines in various tissues including skin [1,2,3,4]. To maintain cellular homeostasis, excessive production of ROS should be neutralized or minimized by cellular antioxidants, including antioxidative enzymes such as heme oxygenase-1 (HMOX1) and NAD(P)H:quinone oxidoreductase 1 (NQO1) [5,6]. The induction of these antioxidative enzymes is upregulated by nuclear factor-erythroid 2-related factor-2 (NRF2), which is a master transcription factor for antioxidant signaling [3,5,6].
Exposure to high concentrations of dioxin induces various acute and chronic health hazards including general fatigue, and neurological (numbness or pain in the limbs), respiratory (cough and sputa), and dermatological symptoms [7,8,9]. In addition, high-dose dioxin intoxication increases the prevalence of cardiovascular diseases, hyperlipidemia, thyroid diseases, diabetes, liver dysfunction, and chronic bronchitis [7,10,11]. Moreover, blood concentrations of dioxins are correlated with conditions, such as general fatigue, increased blood sugar, and hyperlipidemia [12]. The increased rate of mortality associated with liver and lung cancers is an additional important issue in dioxin intoxication [13,14].
Among the cutaneous symptoms caused by dioxin, chloracne is one of the major ones, causing significant deterioration in the quality of daily life [15,16,17,18]. Chloracne has a characteristic skin distribution, frequently affecting the retroauricular and malar areas of the face, ear lobes, and groin, whereas the nose and perioral area are typically spared [16,17,18,19,20]. The severity of chloracne is also correlated with the blood dioxin level [16].
The pathology of chloracne is characterized by hyperkeratinization of the interfollicular epidermis, hyperproliferation and hyperkeratinization of hair follicle cells, gradual loss of sebocytes with shrinkage of sebaceous glands, and infundibular dilatation, eventually leading to comedo formation [3,16,17,18,21,22]. AHR is abundantly expressed in epidermal keratinocytes and sebocytes [3,21]. Moreover, highly lipophilic dioxins appear to accumulate in, and are excreted via, sebaceous glands and sebum [19,23,24], which facilitates dioxin excretion from the intoxicated body [25]. The high concentration of dioxin in the sebum may explain why chloracne frequently develops in individuals with high-dose dioxin intoxication.
In accordance with the histopathology of chloracne, agonistic ligation of AHR accelerates epidermal terminal differentiation and keratinization [26,27,28]. Upon AHR stimulation, the proliferation and lipid synthesis of sebocytes are impaired, probably due to the switching of sebocytes toward keratinocyte-like differentiation [21,29,30]. In this review, we focus on the AHR signaling related to chloracne and highlight its potential treatment with an NRF2 agonist.
2. AHR Signaling in Keratinocytes and Sebocytes
As a chemical sensor, AHR is constitutively expressed in the tissues, separating the inside and outside of the body, including the epidermis and pilosebaceous units [2,3,31]. Dioxins activate AHR and induce its cytoplasmic-to-nuclear translocation. Nuclear AHR binds to its specific DNA recognition site, namely, the xenobiotic responsive element and upregulates the transcription of responsive genes, such as cytochrome p450 1A1 (CYP1A1) in keratinocytes and sebocytes [2,3,31]. CYP1A1 is a xenobiotic-metabolizing enzyme and metabolizes dioxin [1]. As dioxin is very stable and persistent, the metabolizing process by CYP1A1 generates high levels of ROS (Figure 1). In CYP1A1-deficient conditions, ROS production is profoundly attenuated [4,32]. ROS-mediated oxidative stress induces DNA damage and upregulates the production of inflammatory cytokines and chemokines, such as IL-8 and CCL2 from keratinocytes [1,4,33].
In addition to generating oxidative stress, persistent activation of AHR by dioxin accelerates the terminal differentiation of keratinocytes and epidermal hyperkeratosis [27,28]. This effect is mediated by coordinated upregulation of the gene expression of epidermal terminal differentiation molecules, such as filaggrin and proline-rich small proteins (Figure 1) [26,34].
Upon AHR activation by dioxin, sebocytes lose their specific features for sebaceous differentiation, including lipogenesis, keratin 7 expression, and epithelial membrane antigen expression [21]. Instead, AHR ligation converts sebocytes towards keratinocytic differentiation, upregulating keratin 10 and peroxisome proliferator-activated receptor-δ [21]. These findings have been corroborated by ex vivo sebaceous gland cultures where it has been shown that dioxin induces the shrinkage and disappearance of sebaceous glands [21]. These keratinocytic and sebocytic alterations by dioxin coincide with the pathological features of chloracne [18,22].
3. Role of NRF2 in Neutralizing AHR-Mediated Oxidative Stress
Under unstimulated conditions, NRF2 resides in the cytoplasm, but upon activation, it translocates to the nucleus. The antioxidative enzymes downstream of NRF2 include HMOX1, NQO1, glutathione S-transferase, UDP-glucuronosyltransferases, epoxide hydrolase, glutathione reductase, thioredoxin reductase, catalase, and superoxide dismutase. NRF2 also activates the transcription of genes encoding non-enzymatic antioxidative proteins, such as thioredoxin and ferritin [6].
Dioxin induces AHR-mediated ROS production [26,35]. The oxidative stress reciprocally activates the NRF2–antioxidative pathway in order to neutralize excessive ROS generation [36]. However, dioxin is structurally stable and is very difficult to degrade. Therefore, dioxin is capable of activating AHR for a long period. Therefore, persistent activation of the AHR–oxidative pathway by chemically stable dioxin may overwhelm NRF2–antioxidative signaling, leaving the cell in a ROS-rich milieu.
A variety of salubrious, antioxidative plants and herbs utilize NRF2 to exert antioxidative activity. For example, phytoextracts from artichoke in Mediterranean countries, cactus Opuntia ficus-indica in Mexico, and the Asian herb, Houttuynia cordata, inhibit BaP/AHR-mediated oxidative stress via NRF2 activation [37,38,39]. Moreover, NRF2-mediated antioxidative activity is capable of alleviating ROS production induced by tumor necrosis factor-α [37,38,39]. These results highlight that exogenous NRF2 agonists can antagonize dioxin–AHR–ROS signaling.
4. Therapeutic Potential of Cinnamomum cassia-Containing Kampo Herbal Medicine for Chloracne
As antioxidant phytoextracts are potent inhibitors of AHR-mediated oxidative stress, we screened phytoextracts that inhibit the AHR–CYP1A1 pathway and activate the NRF2–antioxidative pathway. Cinnamomum cassia extract and its major constituent cinnamaldehyde have dual activity [40]. Both C. cassia extract and cinnamaldehyde attenuate the AHR–CYP1A1 axis and inhibit oxidative stress [40]. Many Japanese Kampo herbal medicines contain varying doses of C. cassia extract. Among them, Keishibukuryogan is the strongest inhibitor of AHR–CYP1A1 signaling [40]. In addition, both C. cassia extract and cinnamaldehyde activate the NRF2–HMOX1 antioxidative system and inhibit AHR-mediated ROS production (Figure 1) [40].
We conducted a clinical trial of the oral administration of Keishibukuryogan to treat Yusho patients who had been intoxicated with high concentrations of polychlorinated dibenzofurans after they ate a contaminated rice bran oil in 1968 [8,9]. Their mean blood concentrations of polychlorinated dibenzofurans still remained more than 10 times higher than that of normal individuals 30 and 40 years after the accident [41,42]. They suffer from chloracne, general fatigue, numbness and paresthesia of the extremities, cough, and expectoration of sputum [9]. After 3 months of oral administration, Keishibukuryogan significantly attenuated the symptoms of chloracne, general fatigue, and cough, and expectoration of sputum. It also tended to reduce symptoms of numbness and paresthesia of the extremities [9].
Perillaldehyde is another useful phytochemical [33]. It is a flavoring ingredient found in Perilla frutescens (shiso in Japanese), which adds spiciness and a citrus taste to food. Like cinnamaldehyde, perillaldehyde inhibits AHR–CYP1A1 signaling and activates the NRF2–HMOX1 antioxidative axis [33]. Unfortunately, P. frutescens extract-containing Kampo medicines do not exhibit similar activities, suggesting that dried P. frutescens extract may lose the flavoring of perillaldehyde during the extraction process. However, consuming fresh P. frutescens in meals on a daily basis may be helpful in managing chloracne.
5. Conclusions
Chloracne is a devastating skin symptom induced by exposure to high concentrations of dioxins and other hazardous compounds. These environmental pollutants bind to and activate AHR and generate abundant ROS. They also accelerate the terminal differentiation and keratinization of keratinocytes and sebocytes. As dioxin is stable and resistant to metabolization, persistent activation of AHR results in exaggerated oxidative stress and unopposed hyperkeratinization. These features probably explain the pathogenesis of chloracne.
Cinnamaldehyde and perillaldehyde are potent phytochemicals that inhibit the AHR–CYP1A1 pathway and activate the NRF2–antioxidative axis [33,40]. Given that cinnamaldehyde-containing herbal medicine improves the clinical symptoms of patients with dioxin intoxication, agents with dual functions in promoting AHR–CYP1A1 inhibition and NRF2 activation are potential candidates for managing dioxin hazards. Since C. cassia and P. frutescens are inexpensive and popular plants in Asia, their daily ingestion may be a suitable approach for defending against the health hazards of people living in areas contaminated with high levels of dioxins.
Funding
This work was partly supported by a grant (H30-Shokuhin-Shitei-005) for Research on Food Safety from The Ministry of Health, Labour and Welfare, Japan.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- Denison, M.S.; Soshilov, A.A.; He, G.; DeGroot, D.E.; Zhao, B. Exactly the same but different: Promiscuity and diversity in the molecular mechanisms of action of the aryl hydrocarbon (dioxin) receptor. Toxicol. Sci. 2011, 124, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Esser, C.; Bargen, I.; Weighardt, H.; Haarmann-Stemmann, T.; Krutmann, J. Functions of the aryl hydrocarbon receptor in the skin. Semin. Immunopathol. 2013, 35, 677–691. [Google Scholar] [CrossRef] [PubMed]
- Furue, M.; Takahara, M.; Nakahara, T.; Uchi, H. Role of AhR/ARNT system in skin homeostasis. Arch. Dermatol. Res. 2014, 306, 769–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuji, G.; Takahara, M.; Uchi, H.; Takeuchi, S.; Mitoma, C.; Moroi, Y.; Furue, M. An environmental contaminant, benzo(a)pyrene, induces oxidative stress-mediated interleukin-8 production in human keratinocytes via the aryl hydrocarbon receptor signaling pathway. J. Dermatol. Sci. 2011, 62, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Furue, M.; Uchi, H.; Mitoma, C.; Hashimoto-Hachiya, A.; Chiba, T.; Ito, T.; Nakahara, T.; Tsuji, G. Antioxidants for healthy skin: The emerging role of aryl hydrocarbon receptors and nuclear factor-erythroid 2-related factor-2. Nutrients 2017, 9, 223. [Google Scholar] [CrossRef] [PubMed]
- Gęgotek, A.; Skrzydlewska, E. The role of transcription factor Nrf2 in skin cells metabolism. Arch. Dermatol. Res. 2015, 307, 385–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akahane, M.; Matsumoto, S.; Kanagawa, Y.; Mitoma, C.; Uchi, H.; Yoshimura, T.; Furue, M.; Imamura, T. Long-term health effects of PCBs and related compounds: A comparative analysis of patients suffering from Yusho and the general population. Arch. Environ. Contam. Toxicol. 2018, 74, 203–217. [Google Scholar] [CrossRef] [PubMed]
- Mitoma, C.; Uchi, H.; Tsukimori, K.; Yamada, H.; Akahane, M.; Imamura, T.; Utani, A.; Furue, M. Yusho and its latest findings-A review in studies conducted by the Yusho Group. Environ. Int. 2015, 82, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Mitoma, C.; Uchi, H.; Tsukimori, K.; Todaka, T.; Kajiwara, J.; Shimose, T.; Akahane, M.; Imamura, T.; Furue, M. Current state of Yusho and prospects for therapeutic strategies. Environ. Sci. Pollut. Res. Int. 2017. [Google Scholar] [CrossRef]
- Geusau, A.; Abraham, K.; Geissler, K.; Sator, M.O.; Stingl, G.; Tschachler, E. Severe 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) intoxication: clinical and laboratory effects. Environ. Health Perspect. 2001, 109, 865–869. [Google Scholar] [CrossRef] [PubMed]
- Mohsenzadeh, M.S.; Zanjani, B.R.; Karimi, G. Mechanisms of 2,3,7,8-tetrachlorodibenzo-p-dioxin-induced cardiovascular toxicity: An overview. Chem. Biol. Interact. 2018, 282, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kanagawa, Y.; Matsumoto, S.; Koike, S.; Tajima, B.; Fukiwake, N.; Shibata, S.; Uchi, H.; Furue, M.; Imamura, T. Association of clinical findings in Yusho patients with serum concentrations of polychlorinated biphenyls, polychlorinated quarterphenyls and 2,3,4,7,8-pentachlorodibenzofuran more than 30 years after the poisoning event. Environ. Health 2008, 7, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onozuka, D.; Yoshimura, T.; Kaneko, S.; Furue, M. Mortality after exposure to polychlorinated biphenyls and polychlorinated dibenzofurans: A 40-year follow-up study of Yusho patients. Am. J. Epidemiol. 2009, 169, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Li, M.C.; Chen, P.C.; Tsai, P.C.; Furue, M.; Onozuka, D.; Hagihara, A.; Uchi, H.; Yoshimura, T.; Guo, Y.L. Mortality after exposure to polychlorinated biphenyls and polychlorinated dibenzofurans: A meta-analysis of two highly exposed cohorts. Int. J. Cancer 2015, 137, 1427–1432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caputo, R.; Monti, M.; Ermacora, E.; Carminati, G.; Gelmetti, C.; Gianotti, R.; Gianni, E.; Puccinelli, V. Cutaneous manifestations of tetrachlorodibenzo-p-dioxin in children and adolescents. Follow-up 10 years after the Seveso, Italy, accident. J. Am. Acad. Dermatol. 1988, 19, 812–819. [Google Scholar] [CrossRef]
- Mitoma, C.; Mine, Y.; Utani, A.; Imafuku, S.; Muto, M.; Akimoto, T.; Kanekura, T.; Furue, M.; Uchi, H. Current skin symptoms of Yusho patients exposed to high levels of 2,3,4,7,8-pentachlorinated dibenzofuran and polychlorinated biphenyls in 1968. Chemosphere 2015, 137, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Saurat, J.H.; Kaya, G.; Saxer-Sekulic, N.; Pardo, B.; Becker, M.; Fontao, L.; Mottu, F.; Carraux, P.; Pham, X.C.; Barde, C.; et al. The cutaneous lesions of dioxin exposure: Lessons from the poisoning of Victor Yushchenko. Toxicol. Sci. 2012, 125, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Suskind, R.R. Chloracne, “the hallmark of dioxin intoxication”. Scand. J. Work Environ. Health 1985, 11, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Sorg, O.; Zennegg, M.; Schmid, P.; Fedosyuk, R.; Valikhnovskyi, R.; Gaide, O.; Kniazevych, V.; Saurat, J.H. 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) poisoning in Victor Yushchenko: identification and measurement of TCDD metabolites. Lancet 2009, 374, 1179–1185. [Google Scholar] [CrossRef]
- Tindall, J.P. Chloracne and chloracnegens. J. Am. Acad. Dermatol. 1985, 13, 539–558. [Google Scholar] [CrossRef]
- Ju, Q.; Fimmel, S.; Hinz, N.; Stahlmann, R.; Xia, L.; Zouboulis, C.C. 2,3,7,8-Tetrachlorodibenzo-p-dioxin alters sebaceous gland cell differentiation in vitro. Exp. Dermatol. 2011, 20, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Panteleyev, A.A.; Bickers, D.R. Dioxin-induced chloracne—Reconstructing the cellular and molecular mechanisms of a classic environmental disease. Exp. Dermatol. 2006, 15, 705–730. [Google Scholar] [CrossRef] [PubMed]
- Iida, T.; Hirakawa, H.; Matsueda, T.; Takenaka, S.; Yu, M.L.; Guo, Y.L. Recent trend of polychlorinated dibenzo-p-dioxins and their related compounds in the blood and sebum of Yusho and Yu Cheng patients. Chemosphere 1999, 38, 981–993. [Google Scholar] [CrossRef]
- Morokuma, S.; Tsukimori, K.; Hori, T.; Kato, K.; Furue, M. The vernix caseosa is the main site of dioxin excretion in the human foetus. Sci. Rep. 2017, 7, 739. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, S.; Akahane, M.; Kanagawa, Y.; Kajiwara, J.; Todaka, T.; Yasukawa, F.; Uchi, H.; Furue, M.; Imamura, T. Individuals’ half-lives for 2,3,4,7,8-penta-chlorodibenzofuran (PeCDF) in blood: Correlation with clinical manifestations and laboratory results in subjects with Yusho. Chemosphere 2013, 92, 772–777. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, L.H.; Sutter, C.H.; Leon Carrion, S.; Tran, Q.T.; Bodreddigari, S.; Kensicki, E.; Mohney, R.P.; Sutter, T.R. 2,3,7,8-Tetrachlorodibenzo-p-dioxin-mediated production of reactive oxygen species is an essential step in the mechanism of action to accelerate human keratinocyte differentiation. Toxicol. Sci. 2013, 132, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Loertscher, J.A.; Sattler, C.A.; Allen-Hoffmann, B.L. 2,3,7,8-Tetrachlorodibenzo-p-dioxin alters the differentiation pattern of human keratinocytes in organotypic culture. Toxicol. Appl. Pharmacol. 2001, 175, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Loertscher, J.A.; Lin, T.M.; Peterson, R.E.; Allen-Hoffmann, B.L. In utero exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin causes accelerated terminal differentiation in fetal mouse skin. Toxicol. Sci. 2002, 68, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Wang, D.; Yu, Q.; Li, L.; Mo, X.; Pan, Z.; Zouboulis, C.C.; Peng, L.; Xia, L.; Ju, Q. Aryl hydrocarbon receptor negatively regulates lipid synthesis and involves in cell differentiation of SZ95 sebocytes in vitro. Chem. Biol. Interact. 2016, 258, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wu, J.; Song, J.; Liang, P.; Zheng, K.; Xiao, G.; Liu, L.; Zouboulis, C.C.; Lei, T. Particulate matter 2.5 regulates lipid synthesis and inflammatory cytokine production in human SZ95 sebocytes. Int. J. Mol. Med. 2017, 40, 1029–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stockinger, B.; Di Meglio, P.; Gialitakis, M.; Duarte, J.H. The aryl hydrocarbon receptor: Multitasking in the immune system. Annu. Rev. Immunol. 2014, 32, 403–432. [Google Scholar] [CrossRef] [PubMed]
- Anandasadagopan, S.K.; Singh, N.M.; Raza, H.; Bansal, S.; Selvaraj, V.; Singh, S.; Chowdhury, A.R.; Leu, N.A.; Avadhani, N.G. β-Naphthoflavone-induced mitochondrial respiratory damage in Cyp1 knockout mouse and in cell culture systems: Attenuation by resveratrol treatment. Oxid. Med. Cell Longev. 2017, 2017, 5213186. [Google Scholar] [CrossRef] [PubMed]
- Fuyuno, Y.; Uchi, H.; Yasumatsu, M.; Morino-Koga, S.; Tanaka, Y.; Mitoma, C.; Furue, M. Perillaldehyde inhibits AHR signaling and activates NRF2 antioxidant pathway in human keratinocytes. Oxid. Med. Cell Longev. 2018, 2018, 9524657. [Google Scholar] [CrossRef] [PubMed]
- Furue, M.; Tsuji, G.; Mitoma, C.; Nakahara, T.; Chiba, T.; Morino-Koga, S.; Uchi, H. Gene regulation of filaggrin and other skin barrier proteins via aryl hydrocarbon receptor. J. Dermatol. Sci. 2015, 80, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Nie, X.; Liang, L.; Xi, H.; Jiang, S.; Jiang, J.; Tang, C.; Liu, X.; Liu, S.; Wan, C.; Zhao, J.; et al. 2,3,7,8-Tetrachlorodibenzo-p-dioxin induces premature senescence of astrocytes via WNT/β-catenin signaling and ROS production. J. Appl. Toxicol. 2015, 35, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; He, X.; Szklarz, G.D.; Bi, Y.; Rojanasakul, Y.; Ma, Q. The aryl hydrocarbon receptor interacts with nuclear factor erythroid 2-related factor 2 to mediate induction of NAD(P)H:quinoneoxidoreductase 1 by 2,3,7,8-tetrachlorodibenzo-p-dioxin. Arch. Biochem. Biophys. 2013, 537, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Doi, K.; Mitoma, C.; Nakahara, T.; Uchi, H.; Hashimoto-Hachiya, A.; Takahara, M.; Tsuji, G.; Nakahara, M.; Furue, M. Antioxidant Houttuynia cordata extract upregulates filaggrin expression in an aryl hydrocarbon-dependent manner. Fukuoka Igaku Zasshi 2014, 105, 205–213. [Google Scholar] [PubMed]
- Nakahara, T.; Mitoma, C.; Hashimoto-Hachiya, A.; Takahara, M.; Tsuji, G.; Uchi, H.; Yan, X.; Hachisuka, J.; Chiba, T.; Esaki, H.; et al. Antioxidant Opuntia ficus-indica extract activates AHR-NRF2 signaling and upregulates filaggrin and loricrin expression in human keratinocytes. J. Med. Food 2015, 18, 1143–1149. [Google Scholar] [CrossRef] [PubMed]
- Takei, K.; Hashimoto-Hachiya, A.; Takahara, M.; Tsuji, G.; Nakahara, T.; Furue, M. Cynaropicrin attenuates UVB-induced oxidative stress via the AhR-Nrf2-Nqo1 pathway. Toxicol. Lett. 2015, 234, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Uchi, H.; Yasumatsu, M.; Morino-Koga, S.; Mitoma, C.; Furue, M. Inhibition of aryl hydrocarbon receptor signaling and induction of NRF2-mediated antioxidant activity by cinnamaldehyde in human keratinocytes. J. Dermatol. Sci. 2017, 85, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Todaka, T.; Hirakawa, H.; Hori, T.; Tobiishi, K.; Iida, T.; Furue, M. Concentrations of polychlorinated dibenzo-p-dioxins, polychlorinated dibenzofurans, and non-ortho and mono-ortho polychlorinated biphenyls in blood of Yusho patients. Chemosphere 2007, 66, 1983–1989. [Google Scholar] [CrossRef] [PubMed]
- Todaka, T.; Honda, A.; Imaji, M.; Takao, Y.; Mitoma, C.; Furue, M. Effect of colestimide on the concentrations of polychlorinated dibenzo-p-dioxins, polychlorinated dizenzofurans, and polychlorinated biphenyls in blood of Yusho patients. Environ. Health 2016, 15, 63. [Google Scholar] [CrossRef] [PubMed]
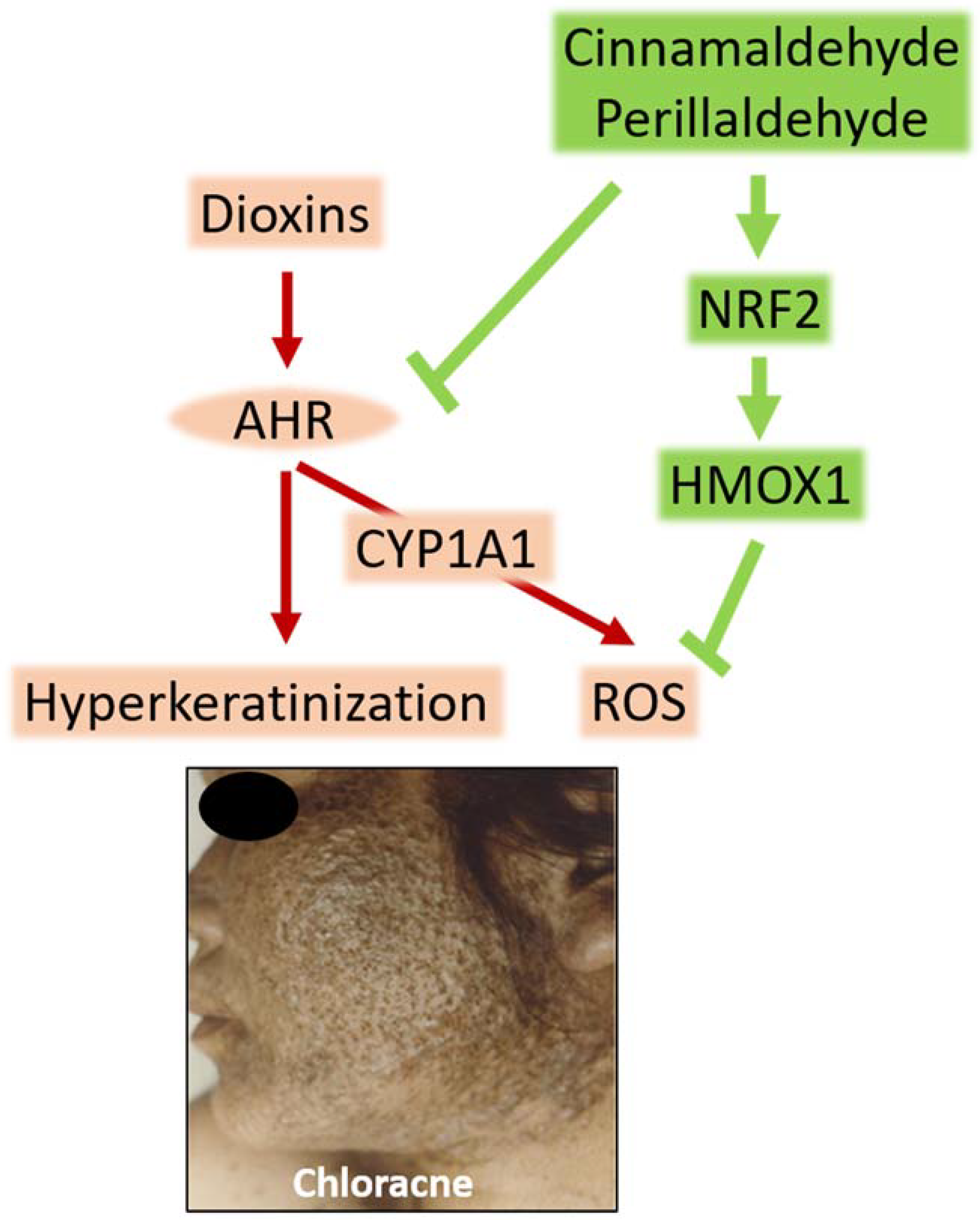
Figure 1.
Dioxins activate the aryl hydrocarbon receptor (AHR), upregulate the expression of cytochrome P450 1A1 (CYP1A1), and generate reactive oxygen species (ROS) in keratinocytes and sebocytes. The ligation of AHR by dioxins also accelerates terminal differentiation. Oxidative stress and hyperkeratinization are probably responsible for chloracne. Cinnamaldehyde (a functional component of C. cassia) and perillaldehyde (a functional component of P. frutescens) are potent inhibitors of AHR–CYP1A1 signaling. On the other hand, they activate nuclear factor-erythroid 2-related factor-2 (NRF2). NRF2 is a master switch for the cellular antioxidative system. The activation of NRF2 upregulates various antioxidative enzymes, such as heme oxygenase-1 (HMOX1), and neutralizes ROS. These natural phytochemicals are useful for managing chloracne.
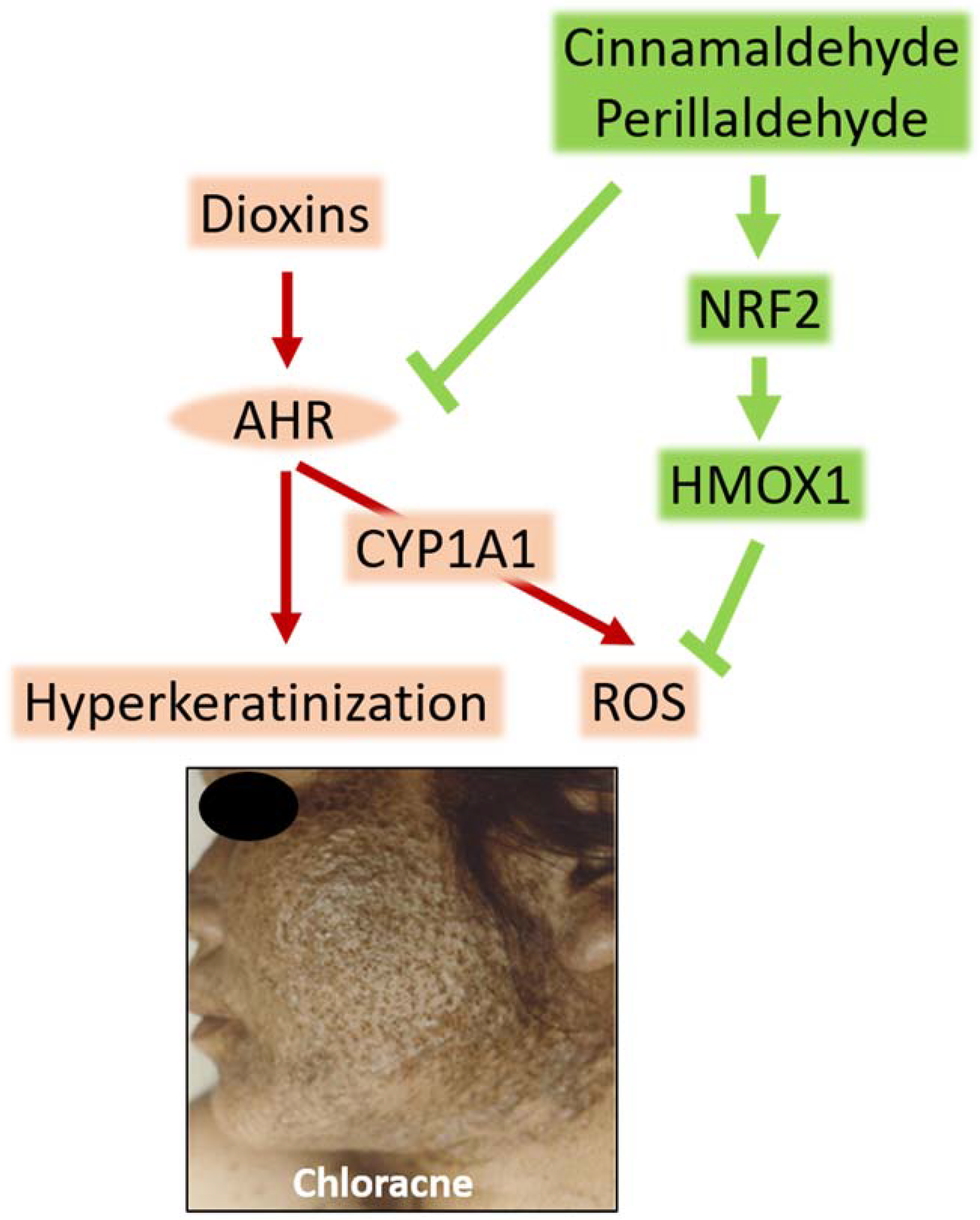
Figure 1.
Dioxins activate the aryl hydrocarbon receptor (AHR), upregulate the expression of cytochrome P450 1A1 (CYP1A1), and generate reactive oxygen species (ROS) in keratinocytes and sebocytes. The ligation of AHR by dioxins also accelerates terminal differentiation. Oxidative stress and hyperkeratinization are probably responsible for chloracne. Cinnamaldehyde (a functional component of C. cassia) and perillaldehyde (a functional component of P. frutescens) are potent inhibitors of AHR–CYP1A1 signaling. On the other hand, they activate nuclear factor-erythroid 2-related factor-2 (NRF2). NRF2 is a master switch for the cellular antioxidative system. The activation of NRF2 upregulates various antioxidative enzymes, such as heme oxygenase-1 (HMOX1), and neutralizes ROS. These natural phytochemicals are useful for managing chloracne.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Furue, M.; Fuyuno, Y.; Mitoma, C.; Uchi, H.; Tsuji, G. Therapeutic Agents with AHR Inhibiting and NRF2 Activating Activity for Managing Chloracne. Antioxidants 2018, 7, 90. https://doi.org/10.3390/antiox7070090
AMA Style
Furue M, Fuyuno Y, Mitoma C, Uchi H, Tsuji G. Therapeutic Agents with AHR Inhibiting and NRF2 Activating Activity for Managing Chloracne. Antioxidants. 2018; 7(7):90. https://doi.org/10.3390/antiox7070090
Chicago/Turabian StyleFurue, Masutaka, Yoko Fuyuno, Chikage Mitoma, Hiroshi Uchi, and Gaku Tsuji. 2018. "Therapeutic Agents with AHR Inhibiting and NRF2 Activating Activity for Managing Chloracne" Antioxidants 7, no. 7: 90. https://doi.org/10.3390/antiox7070090
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.