Abstract
Strenuous exercise induces such inflammatory responses as leukocytosis (neutrophilia) and symptoms as delayed-onset muscle soreness and swelling. However, the association between inflammatory mediator cytokines and oxidative stress is not fully delineated. Herein, in addition to basic background information on cytokines, research findings on exertional effects on cytokine release and the underlying mechanisms and triggers are introduced. Then, the associations among cytokine responses, oxidative stress, and tissue damage are described not only in overloaded skeletal muscle, but also in other internal organs. Furthermore, we introduce preventive countermeasures against the exhaustive exercise-induced pathogenesis together with the possibility of antioxidant interventions.
1. Introduction
Exercise-induced leukocytosis and delayed-onset muscle soreness are well-known phenomena. Redness, swelling, fever, pain, and loss of function are characteristic of inflammation, which is accompanied by leukocyte infiltration, oxidative stress, and production of pro-inflammatory cytokines. Indeed, there has been a tremendous accumulation of research on exercise-induced oxidative stress together with inflammation including from our own research group [1]. However, what are the interactions of oxidative stress and inflammatory mediator cytokines? Also, which are more important for the pathogenic role in exercise-induced tissue damage and as sensitive biomarkers for assessing exercise effects? These are two important questions whose answers remain unknown. Herein, in addition to basic background information on cytokines, research findings on exertional effects on cytokine release and the underlying mechanisms and triggers are introduced. Furthermore, the associations among cytokine responses, oxidative stress and tissue damage are described in overloaded skeletal muscle and other internal organs. Finally, we introduce potential preventive countermeasures against pathogenesis together with the possibility of antioxidant interventions without harmful side-effects.
2. Background Knowledge of Cytokine Function and Release
Cytokines are a diverse family of intercellular signaling molecules that regulate inflammation and immune responses [2,3]. Cytokines are produced by a variety of cells and usually act in an autocrine or paracrine manner at very low concentrations in tissues. Localized inflammation is usually a physiological protective response to initial tissue injury [3]. However, an elevated response can result in cytokine release into the circulation, which becomes pathogenic, self-destructive, and sometimes fatal to the host [2,3]. Systemic inflammatory response syndrome (SIRS) is a condition that is described by the resultant systemic cytokine release (hypercytokinemia, known as cytokine storm). The overproduction of proinflammatory cytokines leads to multiple organ damage, associated with numerous serious acute insults such as severe trauma, thermal injury, ischemia-reperfusion injury, septic shock, and systemic infections [3,4].
Tumor necrosis factor (TNF)-α, interleukin (IL)-1β, IL-6, IL-12, and IL-17 are elevated in sepsis. These pro-inflammatory cytokines are considered the main cytokines in pathogenesis of sepsis. TNF-α appears to be the first cytokine released systemically and peaks within several hours after the onset of sepsis, followed shortly thereafter by peaks in IL-1 and then IL-6 [5,6]. These proinflammatory cytokines induce pyrogenesis and promote subsequent acute inflammatory responses such as leukocytosis (neutrophilia) by inducing granulocyte colony-stimulating factor (G-CSF) and chemokines (abbreviated from chemotactic cytokines) such as IL-8 and monocyte chemotactic protein (MCP)-1 [1,2,5,6].
There are also compensatory anti-inflammatory responses to re-establish homeostasis; some anti-inflammatory cytokines—including IL-1 receptor antagonist (IL-1ra), IL-4 and IL-10—are released into the circulation and dampen the proinflammatory cytokine cascade [5,6]. IL-12 is classified as a major immunomodulatory cytokine, activating cellular immunity. Bioactive IL-12 p70 is a heterodimer composed of two subunits: p35 and p40. The IL-12 p40 homodimer in the absence of p35 expression and free p40 monomer does not mediate IL-12 activity, but acts as an IL-12 antagonist and shares the activity with IL-6 and IL-23, the other neutrophil activating cytokines [2,3,6,7]. As such, some complexities exist in the cytokine network in itself for regulation of inflammation.
3. Cytokine Kinetics in Response to Exercise
Exhaustive endurance exercise induces leukocytosis mainly due to neutrophilia in the systemic circulation, muscle and internal organ damage, and immune suppression [1,8,9,10,11,12,13]. To determine the underlying mechanisms of these phenomena, much attention has been focused on cytokines released into the circulation following exercise. Indeed, many studies have consistently shown that IL-1ra, IL-6, IL-8, and IL-10 increase markedly following endurance exercise lasting longer than several hours, such as marathons and triathlons [5,6,8,10,14,15,16,17,18,19,20]. However, the response of these cytokines is not so significant during and after short-duration intensive exercise [6,7,20,21,22,23,24,25,26,27] and eccentric-contraction exercise [28,29,30,31,32]. These responses are not dependent on exercise-induced muscle damage but are related to exercise intensity (physiological load/stress) [5,6,23].
Indeed, it has been demonstrated that IL-6 response to endurance exercise depends on decreased cellular energy levels and increased heat stress and subsequently correlates with stress hormone responses; however, they are suppressed by increased energy supply [5,8,10,33,34,35,36] and prior body-cooling interventions [4,33]. Furthermore, IL-6 enhances utilization of energy substrates such as free fatty acids, which contribute to endurance performance [5,6,27,35], whilst also inducing neutrophil mobilization and activation together with the anti-inflammatory cytokine release of IL-1ra and IL-10 [1,5,6,13,18,19,36]. Here, IL-1ra is a natural antagonistic cytokine that competes with IL-1 for receptor binding without inducing signal transduction, whilst IL-10 is the most immunosuppressive cytokine. Endurance exercise also increases plasma levels of IL-4 and IL-12p40 (IL-12 antagonist), which might work to block cellular immune response, cause susceptibility to infections [9,10,19,34,35], and might promote inflammation as a component of IL-6 and IL-23 (neutrophil activator).
Chemokines regulate tissue infiltration of leukocytes. IL-8 is a potent neutrophil chemotactic and activation protein referred to as neutrophil activating peptide 1 (NAP-1). IL-8 is released into the circulation under prolonged, intense exercise conditions [5,6,13,18,19,34], whereas short-time intensive exercise also enhances plasma IL-8 concentration [6,23,33]. These findings suggest that not only the duration but also the intensity of exercise might be important for IL-8 release. MCP-1 facilitates infiltration and activation of monocytes and macrophages. We demonstrated that MCP-1 concentration increased significantly not only in plasma but also urine following a marathon race and immediately after short-duration intensive exercise [6,18]. Also, IL-6 and G-CSF are involved in neutrophil mobilization from bone marrow reserves to the circulation after exercise [6,7,13]. Although neutrophils are involved in exercise-induced muscle damage and inflammation, we recently demonstrated that neutrophils mobilized into muscle contribute to exacerbating muscle injury by upregulating proinflammatory cytokine expression through the induction of macrophage infiltration with MCP-1 [37,38]. From this point of view, leukocytosis (neutrophilia) and related variables noted above can be good predictive indicators of exhaustive exercise-induced muscle and other organ damage/dysfunction [1].
4. Examples of Countermeasures to Exercise-Induced Inflammation
Although pre-exercise cooling can effectively attenuate systemic inflammatory response to exhaustive exercise [4,25,33,39,40], post-exercise cooling may not have significant effects [31,33]. Fluid intake has been reported to prevent systemic induction of IL-6 and neutrophil activation markers [41], whilst intensive exercise in the menstruation phase of the menstruation cycle increases systemic inflammation [26]. During the day, evening exercise induces more marked IL-6 release than morning exercise [27]. From a nutraceutical perspective, curcumin ingestion has reduced muscle inflammation after downhill running in a mouse model of exercise-induced muscle damage [42], but the evidence is not sufficient for other antioxidant and/or anti-inflammatory substances for the prevention of muscle damage and inflammation. Intestinal permeability increases following exercise, and endotoxemia occurs, which induces systemic inflammation [4,39,43,44]. Therefore, other countermeasures such as intake of some functional foods for gut barrier protection, bioavailability, and distribution [3,45,46,47,48,49,50,51,52], and appropriate immune responsiveness should be examined in future studies.
5. Conclusions and Future Directions
Whilst IL-6 might be good for athletes in optimizing energy substrate utilization for endurance performance on one hand, it may compromise the immune status of the athlete on the other hand by inducing systemic inflammation and increasing susceptibility to infections (Figure 1). Accumulation of inflammatory cytokines, neutrophils, and macrophages within organs are involved in tissue damage/dysfunction of not only muscles but also of the kidney, liver, and intestines which have recently been investigated in reference to the pathogenesis of multiple organ failure in heat stroke and sepsis and their underlying mechanisms. It is possible that appropriate countermeasures such as exercising in cool environments, and ingesting sufficient energy and fluids together with some functional food(s) might help to maintain endurance performance without causing harmful side effects on health. However, insight into both what does and does not work in regard to exercise-induced inflammation drawn from experimental studies is not enough at present and accumulation of research findings is necessary regardless of positive or negative results. Also, a future focus of countermeasures research is required regarding how the intestines and their permeability may be preserved by functional foods. These countermeasures may not only help to reduce heat illness and promote recovery, but also lead to the introduction of new research findings for the defense to stress and infection.
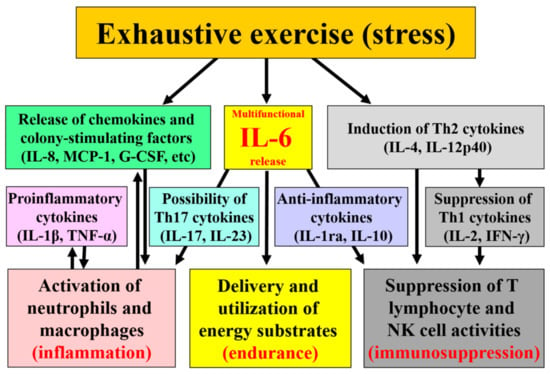
Figure 1.
Cytokines mediate exercise-induced inflammation, immunosuppression and energy metabolism.
Acknowledgments
This work was partly supported by a Grant-in-Aid for the Scientific Research (A) and the Strategic Research Foundation at Private Universities from the Ministry of Education, Culture, Sports, Science and Technology of Japan. I gratefully appreciate my colleagues and laboratory team for research development and progress. Also, I would like to thank Llion Roberts, Lecturer of Human Physiology, School of Allied Health Sciences & Menzies Health Institute Queensland, Griffith University, for English editing.
Author Contributions
Parts of these contents were presented in the Presidential Symposium in 13th International Society of Exercise and Immunology (ISEI) congress (Coimbra, Portugal, 2017). Based on them, K.S. made modifications for this special issue in the conception and design of the manuscript.
Conflicts of Interest
The author declares no conflict of interest.
References
- Suzuki, K. Exhaustive exercise-induced neutrophil-associated tissue damage and possibility of its prevention. J. Nanomed. Biother. Discov. 2017, 7, 156. [Google Scholar] [CrossRef]
- Hasegawa, H.; Mizoguchi, I.; Chiba, Y.; Ohashi, M.; Xu, M.; Yoshimoto, T. Expanding diversity in molecular structures and functions of the IL-6/IL-12 heterodimeric cytokine family. Front. Immunol. 2016, 7, 479. [Google Scholar] [CrossRef] [PubMed]
- Hung, Y.L.; Suzuki, K. The pattern recognition receptors and lipopolysaccharides (LPS)-induced systemic inflammation. Int. J. Res. Stud. Med. Health Sci. 2017, 2, 1–7. [Google Scholar]
- Lim, C.L.; Suzuki, K. Systemic inflammation mediates the effects of endotoxemia in the mechanisms of heat stroke. Biol. Med. 2017, 9, 376. [Google Scholar] [CrossRef]
- Pedersen, B.K.; Febbraio, M.A. Muscle as an endocrine organ: Focus on muscle-derived interleukin-6. Physiol. Rev. 2008, 88, 1379–1406. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Nakaji, S.; Yamada, M.; Totsuka, M.; Sato, K.; Sugawara, K. Systemic inflammatory response to exhaustive exercise: Cytokine kinetics. Exerc. Immunol. Rev. 2002, 8, 6–48. [Google Scholar] [PubMed]
- Yamada, M.; Suzuki, K.; Kudo, S.; Totsuka, M.; Nakaji, S.; Sugawara, K. Raised plasma G-CSF and IL-6 after exercise may play a role in neutrophil mobilization into the circulation. J. Appl. Physiol. 2002, 92, 1789–1794. [Google Scholar] [CrossRef] [PubMed]
- Nieman, D.C. Immune response to heavy exertion. J. Appl. Physiol. 1997, 82, 1385–1394. [Google Scholar] [CrossRef] [PubMed]
- Nieman, D.C. Is infection risk linked to exercise workload? Med. Sci. Sports Exerc. 2000, 32 (Suppl. 7), S406–S411. [Google Scholar] [CrossRef] [PubMed]
- Goh, J.M.; Lim, C.L.; Suzuki, K. Effects of endurance-, strength-, and concurrent training on cytokines and inflammation. In Concurrent Aerobic and Strength Training: Scientific Basics and Practical Applications; Schumann, M., Rønnestad, B., Eds.; Springer: Datenschutz, Germany, 2018; in press. [Google Scholar]
- Suzuki, K.; Naganuma, S.; Totsuka, M.; Suzuki, K.J.; Mochizuki, M.; Shiraishi, M.; Nakaji, S.; Sugawara, K. Effects of exhaustive endurance exercise and its one-week daily repetition on neutrophil count and functional status in untrained men. Int. J. Sports Med. 1996, 17, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Sato, H.; Kikuchi, T.; Abe, T.; Nakaji, S.; Sugawara, K.; Totsuka, M.; Sato, K.; Yamaya, K. Capacity of circulating neutrophils to produce reactive oxygen species after exhaustive exercise. J. Appl. Physiol. 1996, 81, 1213–1222. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Totsuka, M.; Nakaji, S.; Yamada, M.; Kudoh, S.; Liu, Q.; Sugawara, K.; Yamaya, K.; Sato, K. Endurance exercise causes interaction among stress hormones, cytokines, neutrophil dynamics, and muscle damage. J. Appl. Physiol. 1999, 87, 1360–1367. [Google Scholar] [CrossRef] [PubMed]
- Sugama, K.; Suzuki, K.; Yoshitani, K.; Shiraishi, K.; Kometani, T. Urinary excretion of cytokines versus their plasma levels after endurance exercise. Exerc. Immunol. Rev. 2013, 19, 29–48. [Google Scholar] [PubMed]
- Sugama, K.; Suzuki, K.; Yoshitani, K.; Shiraishi, K.; Kometani, T. IL-17, neutrophil activation and muscle damage following endurance exercise. Exerc. Immunol. Rev. 2012, 18, 116–127. [Google Scholar] [PubMed]
- Sugama, K.; Suzuki, K.; Yoshitani, K.; Shiraishi, K.; Miura, S.; Yoshioka, H.; Mori, Y.; Kometani, T. Changes of thioredoxin, oxidative stress markers, inflammation and muscle/renal damage following intensive endurance exercise. Exerc. Immunol. Rev. 2015, 21, 130–142. [Google Scholar] [PubMed]
- Suzuki, K.; Peake, J.; Nosaka, K.; Okutsu, M.; Abbiss, C.R.; Surriano, R.; Bishop, D.; Quod, M.J.; Lee, H.; Martin, D.T.; et al. Changes in markers of muscle damage, inflammation and HSP70 after an Ironman triathlon race. Eur. J. Appl. Physiol. 2006, 98, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Nakaji, S.; Yamada, M.; Liu, Q.; Kurakake, S.; Okamura, N.; Kumae, T.; Umeda, T.; Sugawara, K. Impact of a competitive marathon race on systemic cytokine and neutrophil responses. Med. Sci. Sports Exerc. 2003, 35, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Yamada, M.; Kurakake, S.; Okamura, N.; Yamaya, K.; Liu, Q.; Kudoh, S.; Kowatari, K.; Nakaji, S.; Sugawara, K. Circulating cytokines and hormones with immunosuppressive but neutrophil-priming potentials rise after endurance exercise in humans. Eur. J. Appl. Physiol. 2000, 81, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Peake, J.; Della Gatta, P.; Suzuki, K.; Nieman, D. Cytokine expression and secretion by skeletal muscle cells: Regulatory mechanisms and exercise effects. Exerc. Immunol. Rev. 2015, 21, 8–25. [Google Scholar] [PubMed]
- Suzuki, K.; Takahashi, M.; Li, C.Y.; Lin, S.P.; Tomari, M.; Shing, C.M.; Fang, S.H. The acute effects of green tea and carbohydrate co-ingestion on systemic inflammation and oxidative stress during sprint cycling. Appl. Physiol. Nutr. Metab. 2015, 40, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- Peake, J.; Peiffer, J.J.; Abbiss, C.R.; Nosaka, K.; Okutsu, M.; Laursen, P.B.; Suzuki, K. Body temperature and its effect on leukocyte mobilization, cytokines and markers of neutrophil activation during and after exercise. Eur. J. Appl. Physiol. 2008, 102, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Peake, J.; Wilson, G.; Hordern, M.; Suzuki, K.; Yamaya, K.; Nosaka, K.; Mackinnon, L.; Coombes, J.S. Changes in neutrophil surface receptor expression, degranulation, and respiratory burst activity after moderate- and high-intensity exercise. J. Appl. Physiol. 2004, 97, 612–618. [Google Scholar] [CrossRef] [PubMed]
- Hayashida, H.; Shimura, M.; Sugama, K.; Kanda, K.; Suzuki, K. Effects of the menstrual cycle and acute aerobic exercise on cytokine levels. J. Sports Med. Doping Stud. 2015, 6, 173. [Google Scholar] [CrossRef]
- Peake, J.; Peiffer, J.J.; Abbiss, C.R.; Nosaka, K.; Laursen, P.B.; Suzuki, K. Carbohydrate gel ingestion and immunoendocrine responses to cycling in temperate and hot conditions. Int. J. Sport Nutr. Exerc. Metab. 2008, 18, 229–246. [Google Scholar] [CrossRef] [PubMed]
- Hayashida, H.; Shimura, M.; Sugama, K.; Kanda, K.; Suzuki, K. Exercise-induced inflammation during different phases of the menstrual cycle. Physiother. Rehabil. 2016, 1, 121. [Google Scholar] [CrossRef]
- Kim, H.K.; Konishi, M.; Takahashi, M.; Tabata, H.; Endo, N.; Numao, S.; Lee, S.K.; Suzuki, K.; Kim, Y.H.; Sakamoto, S. Effects of acute endurance exercise performed in the morning and evening on inflammatory cytokine and metabolic hormone responses. PLoS ONE 2015, 10, e0137567. [Google Scholar] [CrossRef] [PubMed]
- Peake, J.; Nosaka, K.; Suzuki, K. Characterization of inflammatory responses to eccentric exercise in humans. Exerc. Immunol. Rev. 2005, 11, 64–85. [Google Scholar] [PubMed]
- Kanda, K.; Sugama, K.; Sakuma, J.; Kawakami, Y.; Suzuki, K. Evaluation of serum leaking enzymes and investigation into new biomarkers for exercise-induced muscle damage. Exerc. Immunol. Rev. 2014, 20, 39–54. [Google Scholar] [PubMed]
- Kanda, K.; Sugama, K.; Hayashida, H.; Sakuma, J.; Kawakami, Y.; Miura, S.; Yoshioka, H.; Mori, Y.; Suzuki, K. Eccentric exercise-induced delayed-onset muscle soreness and changes in markers of muscle damage and inflammation. Exerc. Immunol. Rev. 2013, 19, 74–87. [Google Scholar]
- Peake, J.M.; Roberts, L.A.; Figueiredo, V.C.; Egner, I.; Krog, S.; Aas, S.N.; Suzuki, K.; Markworth, J.F.; Coombes, J.S.; Cameron-Smith, D.; et al. The effects of cold water immersion and active recovery on inflammation and cell stress responses in human skeletal muscle after resistance exercise. J. Physiol. 2017, 595, 695–711. [Google Scholar] [CrossRef] [PubMed]
- Peake, J.M.; Suzuki, K.; Wilson, G.; Hordern, M.; Nosaka, K.; Mackinnon, L.; Coombs, J. Exercise-induced muscle damage, plasma cytokines, and markers of neutrophil activation. Med. Sci. Sports Exerc. 2005, 37, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Kondou, T.; Ogawa, K.; Terada, O.; Kim, K.; Okutsu, M.; Suzuki, K. Cytokine response to exercise during exposure to cold. Jpn. J. Complement. Altern. Med. 2009, 6, 89–95. [Google Scholar] [CrossRef]
- Suzuki, K.; Shiraishi, K.; Yoshitani, K.; Sugama, K.; Kometani, T. The effect of a sports drink based on highly branched cyclic dextrin on cytokine responses to exhaustive endurance exercise. J. Sports Med. Phys. Fit. 2014, 54, 622–630. [Google Scholar] [PubMed]
- Suzuki, K.; Nakaji, S.; Kurakake, S.; Totsuka, M.; Sato, K.; Kuriyama, T.; Fujimoto, H.; Shibusawa, K.; Machida, K.; Sugawara, K. Exhaustive exercise and type-1/type-2 cytokine balance in special focus on interleukin-12 p40/p70. Exerc. Immunol. Rev. 2003, 9, 48–57. [Google Scholar] [PubMed]
- Hashimoto, H.; Ishijima, T.; Hayashida, H.; Suzuki, K.; Higuchi, M. Menstrual cycle phase and carbohydrate ingestion alter immune response following endurance exercise and high intensity time trial performance test under hot conditions. J. Int. Soc. Sports Nutr. 2014, 11, 39. [Google Scholar] [CrossRef] [PubMed]
- Kawanishi, N.; Mizokami, T.; Niihara, H.; Yada, K.; Suzuki, K. Neutrophil depletion attenuates muscle injury after exhaustive exercise. Med. Sci. Sports Exerc. 2016, 48, 1917–1924. [Google Scholar] [CrossRef] [PubMed]
- Kawanishi, N.; Mizokami, T.; Niihara, H.; Yada, K.; Suzuki, K. Macrophage depletion by clodronate liposome attenuates muscle injury and inflammation following exhaustive exercise. Biochem. Biophys. Rep. 2016, 5, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.L.; Pyne, D.B.; Horn, P.; Kalz, A.; Saunders, P.; Peake, J.; Suzuki, K.; Wilson, G.; Mackinnon, L.T. The effects of increased endurance training load on biomarkers of heat tolerance during intense exercise in the heat. Appl. Physiol. Nutr. Metab. 2009, 34, 616–624. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, H.; Ishijima, T.; Suzuki, K.; Higuchi, M. The effect of the menstrual cycle and water consumption on physiological responses during prolonged exercise at moderate intensity in hot conditions. J. Sports Med. Phys. Fit. 2016, 56, 951–960. [Google Scholar] [PubMed]
- Suzuki, K.; Hashimoto, H.; Oh, T.; Ishijima, T.; Mitsuda, H.; Peake, J.M.; Sakamoto, S.; Muraoka, I.; Higuchi, M. The effects of sports drink osmolality on fluid intake and immunoendocrine responses to cycling in hot conditions. J. Nutr. Sci. Vitaminol. 2013, 59, 206–212. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kawanishi, N.; Kato, K.; Takahashi, M.; Mizokami, T.; Otsuka, Y.; Imaizumi, A.; Shiva, D.; Yano, H.; Suzuki, K. Curcumin attenuates oxidative stress following downhill running-induced muscle damage. Biochem. Biophys. Res. Commun. 2013, 441, 573–578. [Google Scholar] [CrossRef] [PubMed]
- Pires, W.; Veneroso, C.E.; Wanner, S.P.; Pacheco, D.A.S.; Vaz, G.C.; Amorim, F.T.; Tonoli, C.; Soares, D.D.; Coimbra, C.C. Association between exercise-induced hyperthermia and intestinal permeability: A systematic review. Sports Med. 2017, 47, 1389–1403. [Google Scholar] [CrossRef] [PubMed]
- Van Wijck, K.; Lenaerts, K.; Grootjans, J.; Wijnands, K.A.; Poeze, M.; van Loon, L.J.; Dejong, C.H.; Buurman, W.A. Physiology and pathophysiology of splanchnic hypoperfusion and intestinal injury during exercise: Strategies for evaluation and prevention. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G155–G168. [Google Scholar] [CrossRef] [PubMed]
- Peake, J.M.; Suzuki, K.; Coombes, J.S. The influence of antioxidant supplementation on markers of inflammation and the relationship to oxidative stress after exercise. J. Nutr. Biochem. 2007, 18, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Ohno, S.; Ohno, Y.; Suzuki, Y.; Miura, S.; Yoshioka, H.; Mori, Y.; Suzuki, K. Ingestion of Tabebuia avellanedae (Taheebo) inhibits production of reactive oxygen species from human peripheral blood neutrophils. Int. J. Food Sci. Nutr. Diet. 2015, S6, 1–4. [Google Scholar]
- Suzuki, K.; Ohno, S.; Suzuki, Y.; Ohno, Y.; Okuyama, R. Effect of green tea extract on reactive oxygen species produced by neutrophils from cancer patients. Anticancer Res. 2012, 32, 2369–2375. [Google Scholar] [PubMed]
- Takahashi, M.; Suzuki, K.; Kim, H.K.; Otsuka, Y.; Imaizumi, A.; Miyashita, M.; Sakamoto, S. Effects of curcumin supplementation on exercise-induced oxidative stress in humans. Int. J. Sports Med. 2014, 35, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Li, C.Y.; Suzuki, K.; Hung, Y.L.; Yang, M.S.; Yu, C.P.; Lin, S.P.; Hou, Y.C.; Fang, S.H. Aloe metabolites prevent LPS-induced sepsis and inflammatory response by inhibiting mitogen-activated protein kinase activation. Am. J. Chin. Med. 2017, 45, 847–861. [Google Scholar] [CrossRef] [PubMed]
- Hung, Y.L.; Fang, S.H.; Wang, S.C.; Cheng, W.C.; Liu, P.L.; Su, C.C.; Chen, C.S.; Huang, M.Y.; Hua, K.F.; Shen, K.H.; et al. Corylin protects LPS-induced sepsis and attenuates LPS-induced inflammatory response. Sci. Rep. 2017, 7, 46299. [Google Scholar] [CrossRef] [PubMed]
- Asadi, A.; Arazi, H.; Suzuki, K. Effects of β-hydroxy-β-methylbutyrate-free acid supplementation on strength, power and hormonal adaptations following resistance training. Nutrients 2017, 9, 1316. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Yada, K.; Lee, H.; Fukuda, Y.; Iida, A.; Suzuki, K. Taheebo polyphenols attenuate FFA-induced inflammation in murine and human macrophage cell lines as inhibitor of COX-2. Front. Nutr. 2017, 4, 63. [Google Scholar] [CrossRef] [PubMed]

© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).