


Mitigating Doxorubicin-Induced Skeletal Muscle Toxicity: A Review of Oxidative Stress Mechanisms and the Therapeutic Role of Exercise

{kind=link}
{kind=link}
Abstract
1. Introduction
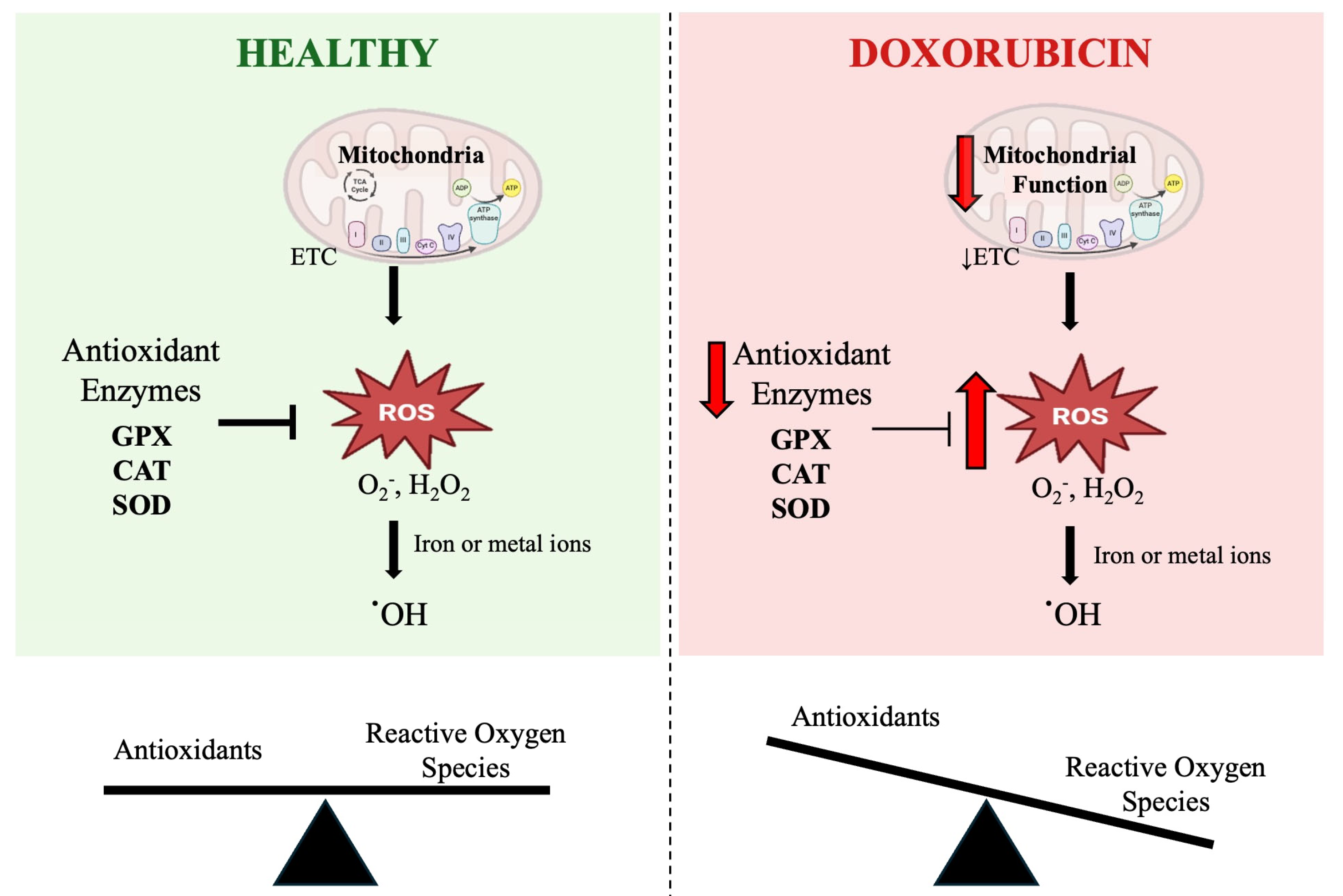
2. Reactive Oxygen Species and Oxidative Stress
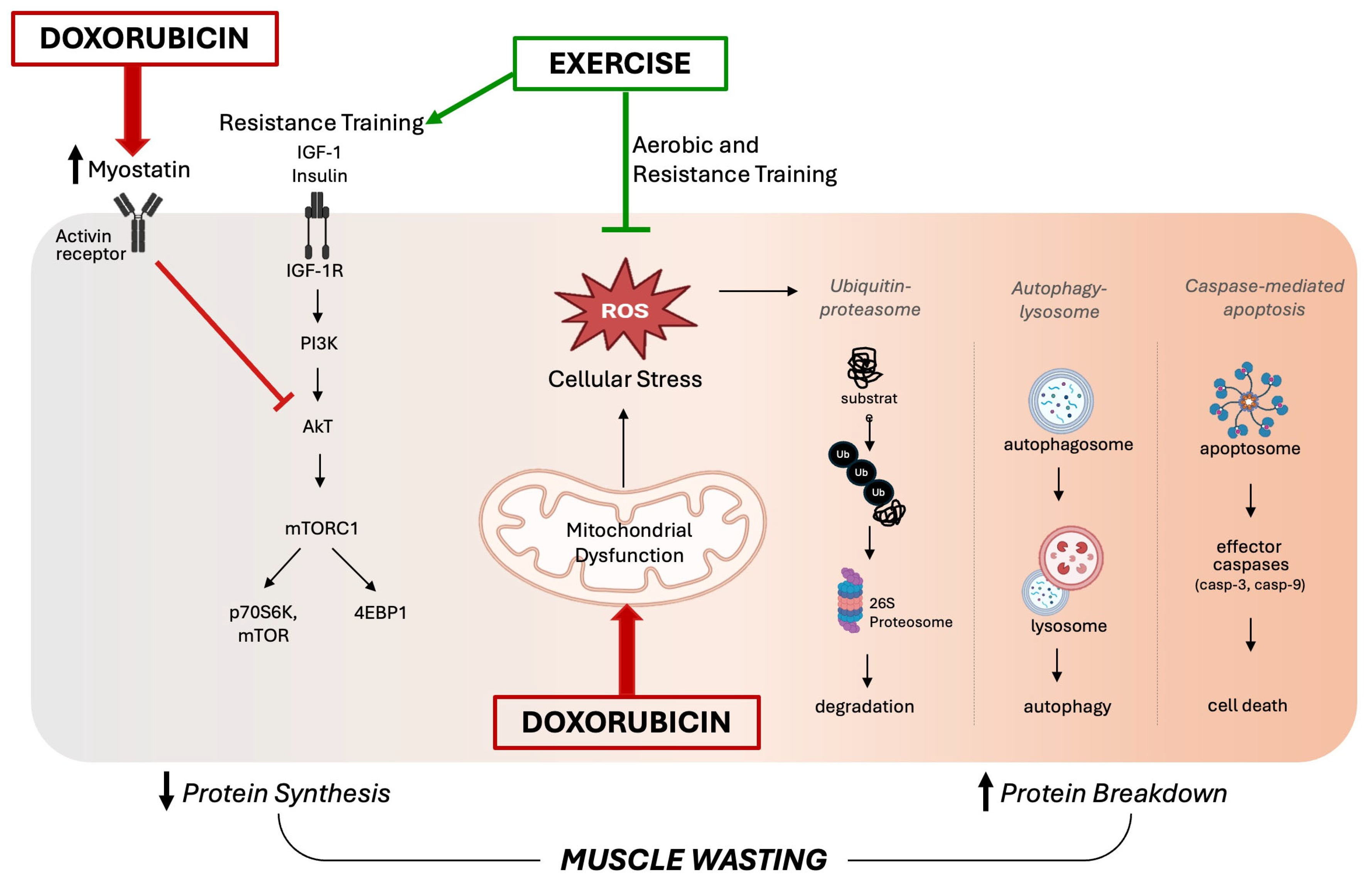
3. Role of Oxidative Stress in DOX-Induced Skeletal Muscle Wasting
4. Role of Exercise Against DOX-Induced Skeletal Muscle Wasting
5. Future Directions
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ahmad, F.B.; Anderson, R.N. The Leading Causes of Death in the US for 2020. Jama 2021, 325, 1829–1830. [Google Scholar] [CrossRef] [PubMed]
- National Cancer Institute. Cancer Statistics. Available online: https://www.cancer.gov/about-cancer/understanding/statistics (accessed on 7 January 2025).
- Johnson-Arbor, K.D. Ramin. Doxorubicin. Available online: https://www.ncbi.nlm.nih.gov/books/NBK459232/ (accessed on 20 January 2025).
- Alhowail, A.H.; Bloemer, J.; Majrashi, M.; Pinky, P.D.; Bhattacharya, S.; Yongli, Z.; Bhattacharya, D.; Eggert, M.; Woodie, L.; Buabeid, M.A.; et al. Doxorubicin-induced neurotoxicity is associated with acute alterations in synaptic plasticity, apoptosis, and lipid peroxidation. Toxicol. Mech. Methods 2019, 29, 457–466. [Google Scholar] [CrossRef] [PubMed]
- El-Moselhy, M.A.; El-Sheikh, A.A. Protective mechanisms of atorvastatin against doxorubicin-induced hepato-renal toxicity. Biomed. Pharmacother. 2014, 68, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Pfizer. Product Information—Adriamycin (Doxorubicin Hydrochloride). Available online: https://labeling.pfizer.com/ShowLabeling.aspx?id=12164 (accessed on 15 January 2025).
- Gilliam, L.A.; St Clair, D.K. Chemotherapy-induced weakness and fatigue in skeletal muscle: The role of oxidative stress. Antioxid. Redox Signal. 2011, 15, 2543–2563. [Google Scholar] [CrossRef] [PubMed]
- Doerr, V.; Montalvo, R.N.; Kwon, O.S.; Talbert, E.E.; Hain, B.A.; Houston, F.E.; Smuder, A.J. Prevention of Doxorubicin-Induced Autophagy Attenuates Oxidative Stress and Skeletal Muscle Dysfunction. Antioxidants 2020, 9, 263. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, Y.; Ghanefar, M.; Bayeva, M.; Wu, R.; Khechaduri, A.; Naga Prasad, S.V.; Mutharasan, R.K.; Naik, T.J.; Ardehali, H. Cardiotoxicity of doxorubicin is mediated through mitochondrial iron accumulation. J. Clin. Investig. 2014, 124, 617–630. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.J.; Martinez, P.F.; Pagan, L.U.; Damatto, R.L.; Cezar, M.D.M.; Lima, A.R.R.; Okoshi, K.; Okoshi, M.P. Skeletal muscle aging: Influence of oxidative stress and physical exercise. Oncotarget 2017, 8, 20428–20440. [Google Scholar] [CrossRef] [PubMed]
- Penna, F.; Ballarò, R.; Costelli, P. The Redox Balance: A Target for Interventions Against Muscle Wasting in Cancer Cachexia? Antioxid. Redox Signal. 2020, 33, 542–558. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Duarte, J.A.; Le Nguyen, B.; Hyatt, H. Endurance exercise protects skeletal muscle against both doxorubicin-induced and inactivity-induced muscle wasting. Pflugers Arch. 2019, 471, 441–453. [Google Scholar] [CrossRef] [PubMed]
- Simioni, C.; Zauli, G.; Martelli, A.M.; Vitale, M.; Sacchetti, G.; Gonelli, A.; Neri, L.M. Oxidative stress: Role of physical exercise and antioxidant nutraceuticals in adulthood and aging. Oncotarget 2018, 9, 17181–17198. [Google Scholar] [CrossRef] [PubMed]
- Songbo, M.; Lang, H.; Xinyong, C.; Bin, X.; Ping, Z.; Liang, S. Oxidative stress injury in doxorubicin-induced cardiotoxicity. Toxicol. Lett. 2019, 307, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Snezhkina, A.V.; Kudryavtseva, A.V.; Kardymon, O.L.; Savvateeva, M.V.; Melnikova, N.V.; Krasnov, G.S.; Dmitriev, A.A. ROS Generation and Antioxidant Defense Systems in Normal and Malignant Cells. Oxid. Med. Cell. Longev. 2019, 2019, 6175804. [Google Scholar] [CrossRef] [PubMed]
- Storz, P. Reactive oxygen species-mediated mitochondria-to-nucleus signaling: A key to aging and radical-caused diseases. Sci. STKE 2006, 2006, re3. [Google Scholar] [CrossRef] [PubMed]
- Smuder, A.J. Exercise stimulates beneficial adaptations to diminish doxorubicin-induced cellular toxicity. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2019, 317, R662–R672. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M.; Okutsu, M. Interleukin-1β triggers muscle-derived extracellular superoxide dismutase expression and protects muscles from doxorubicin-induced atrophy. J. Physiol. 2023, 601, 4699–4721. [Google Scholar] [CrossRef] [PubMed]
- Vitale, R.; Marzocco, S.; Popolo, A. Role of Oxidative Stress and Inflammation in Doxorubicin-Induced Cardiotoxicity: A Brief Account. Int. J. Mol. Sci. 2024, 25, 7477. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, C.; Santos, R.X.; Cardoso, S.; Correia, S.; Oliveira, P.J.; Santos, M.S.; Moreira, P.I. Doxorubicin: The good, the bad and the ugly effect. Curr. Med. Chem. 2009, 16, 3267–3285. [Google Scholar] [CrossRef] [PubMed]
- Boeno, F.P.; Patel, J.; Montalvo, R.N.; Lapierre-Nguyen, S.S.; Schreiber, C.M.; Smuder, A.J. Effects of Exercise Preconditioning on Doxorubicin-Induced Liver and Kidney Toxicity in Male and Female Rats. Int. J. Mol. Sci. 2023, 24, 10222. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Deminice, R.; Ozdemir, M.; Yoshihara, T.; Bomkamp, M.P.; Hyatt, H. Exercise-induced oxidative stress: Friend or foe? J. Sport Health Sci. 2020, 9, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Fearon, K.; Strasser, F.; Anker, S.D.; Bosaeus, I.; Bruera, E.; Fainsinger, R.L.; Jatoi, A.; Loprinzi, C.; MacDonald, N.; Mantovani, G.; et al. Definition and classification of cancer cachexia: An international consensus. Lancet Oncol. 2011, 12, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Ciciliot, S.; Rossi, A.C.; Dyar, K.A.; Blaauw, B.; Schiaffino, S. Muscle type and fiber type specificity in muscle wasting. Int. J. Biochem. Cell Biol. 2013, 45, 2191–2199. [Google Scholar] [CrossRef] [PubMed]
- Hiensch, A.E.; Bolam, K.A.; Mijwel, S.; Jeneson, J.A.L.; Huitema, A.D.R.; Kranenburg, O.; van der Wall, E.; Rundqvist, H.; Wengstrom, Y.; May, A.M. Doxorubicin-induced skeletal muscle atrophy: Elucidating the underlying molecular pathways. Acta Physiol. 2020, 229, e13400. [Google Scholar] [CrossRef] [PubMed]
- Huertas, A.M.; Morton, A.B.; Hinkey, J.M.; Ichinoseki-Sekine, N.; Smuder, A.J. Modification of Neuromuscular Junction Protein Expression by Exercise and Doxorubicin. Med. Sci. Sports Exerc. 2020, 52, 1477–1484. [Google Scholar] [CrossRef] [PubMed]
- Filho, A.; Stoyell-Conti, F.F.; Gauri, A.; Lima, L.S.S.; Bernardes, N.; Irigoyen, M.C.; Singal, P.; De Angelis, K.; Dias, D.D.S. Vitamin C protects against doxorubicin induced skeletal muscle atrophy: Role of oxidative stress. Can. J. Physiol. Pharmacol. 2025, 103, 6. [Google Scholar] [CrossRef] [PubMed]
- Kesik, V.; Honca, T.; Gulgun, M.; Uysal, B.; Kurt, Y.G.; Cayci, T.; Babacan, O.; Gocgeldi, E.; Korkmazer, N. Myostatin as a Marker for Doxorubicin Induced Cardiac Damage. Ann. Clin. Lab. Sci. 2016, 46, 26–31. [Google Scholar] [PubMed]
- McFarlane, C.; Plummer, E.; Thomas, M.; Hennebry, A.; Ashby, M.; Ling, N.; Smith, H.; Sharma, M.; Kambadur, R. Myostatin induces cachexia by activating the ubiquitin proteolytic system through an NF-kappaB-independent, FoxO1-dependent mechanism. J. Cell. Physiol. 2006, 209, 501–514. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.H.; Burns, S.P. Increasing Lean Mass and Strength: A Comparison of High Frequency Strength Training to Lower Frequency Strength Training. Int. J. Exerc. Sci. 2016, 9, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Vincent, H.K.; Powers, S.K.; Stewart, D.J.; Demirel, H.A.; Shanely, R.A.; Naito, H. Short-term exercise training improves diaphragm antioxidant capacity and endurance. Eur. J. Appl. Physiol. 2000, 81, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Carapeto, P.V.; Aguayo-Mazzucato, C. Effects of exercise on cellular and tissue aging. Aging (Albany NY) 2021, 13, 14522–14543. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, S.; Meng, X.; Zhao, X.; Wang, T.; Lei, Z.; Lehmann, H.I.; Li, G.; Alcaide, P.; Bei, Y.; et al. Exercise Inhibits Doxorubicin-Induced Cardiotoxicity via Regulating B Cells. Circ. Res. 2024, 134, 550–568. [Google Scholar] [CrossRef] [PubMed]
- Bredahl, E.C.; Pfannenstiel, K.B.; Quinn, C.J.; Hayward, R.; Hydock, D.S. Effects of Exercise on Doxorubicin-Induced Skeletal Muscle Dysfunction. Med. Sci. Sports Exerc. 2016, 48, 1468–1473. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Z.; Guo, H.Z.; Leng, J.Z.; Liang, Z.D.; Wang, J.T.; Luo, L.J.; Wang, S.Q.; Yuan, Y. Exercise preconditioning inhibits doxorubicin-induced cardiotoxicity via YAP/STAT3 signaling. Heliyon 2024, 10, e27035. [Google Scholar] [CrossRef] [PubMed]
- Zouhal, H.; Jayavel, A.; Parasuraman, K.; Hayes, L.D.; Tourny, C.; Rhibi, F.; Laher, I.; Abderrahman, A.B.; Hackney, A.C. Effects of Exercise Training on Anabolic and Catabolic Hormones with Advanced Age: A Systematic Review. Sports Med. 2022, 52, 1353–1368. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, M.; Bishop, N.C.; Stensel, D.J.; Lindley, M.R.; Mastana, S.S.; Nimmo, M.A. The anti-inflammatory effects of exercise: Mechanisms and implications for the prevention and treatment of disease. Nat. Rev. Immunol. 2011, 11, 607–615. [Google Scholar] [CrossRef] [PubMed]
- No, M.H.; Heo, J.W.; Yoo, S.Z.; Kim, C.J.; Park, D.H.; Kang, J.H.; Seo, D.Y.; Han, J.; Kwak, H.B. Effects of aging and exercise training on mitochondrial function and apoptosis in the rat heart. Pflügers Arch.-Eur. J. Physiol. 2020, 472, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Montalvo, R.N.; Doerr, V.; Min, K.; Szeto, H.H.; Smuder, A.J. Doxorubicin-induced oxidative stress differentially regulates proteolytic signaling in cardiac and skeletal muscle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2020, 318, R227–R233. [Google Scholar] [CrossRef] [PubMed]
- Kwon, I. Endurance exercise training reinforces muscular strength with improvements in mitochondrial oxidative capacity, lysosome reformation, and myogenic differentiation against doxorubicin-induced skeletal muscle wasting in mice. Phys. Act. Nutr. 2023, 27, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.J.; Martinez, P.F.; Campos, D.H.; Pagan, L.U.; Bonomo, C.; Lima, A.R.; Damatto, R.L.; Cezar, M.D.; Damatto, F.C.; Rosa, C.M.; et al. Beneficial Effects of Physical Exercise on Functional Capacity and Skeletal Muscle Oxidative Stress in Rats with Aortic Stenosis-Induced Heart Failure. Oxid. Med. Cell. Longev. 2016, 2016, 8695716. [Google Scholar] [CrossRef] [PubMed]
- Smuder, A.J.; Kavazis, A.N.; Min, K.; Powers, S.K. Exercise protects against doxorubicin-induced markers of autophagy signaling in skeletal muscle. J. Appl. Physiol. 2011, 111, 1190–1198. [Google Scholar] [CrossRef] [PubMed]
- Smuder, A.J.; Kavazis, A.N.; Min, K.; Powers, S.K. Exercise protects against doxorubicin-induced oxidative stress and proteolysis in skeletal muscle. J. Appl. Physiol. 2011, 110, 935–942. [Google Scholar] [CrossRef] [PubMed]
- Chow, L.S.; Gerszten, R.E.; Taylor, J.M.; Pedersen, B.K.; van Praag, H.; Trappe, S.; Febbraio, M.A.; Galis, Z.S.; Gao, Y.; Haus, J.M.; et al. Exerkines in health, resilience and disease. Nat Rev Endocrinol 2022, 18, 273–289. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Goldstein, E.; Schrager, M.; Ji, L.L. Exercise Training and Skeletal Muscle Antioxidant Enzymes: An Update. Antioxidants 2022, 12, 39. [Google Scholar] [CrossRef] [PubMed]
- Félix-Soriano, E.; Stanford, K.I. Exerkines and redox homeostasis. Redox Biol. 2023, 63, 102748. [Google Scholar] [CrossRef] [PubMed]
- Belhadjali, F.; Ghrir, S.; Ksia, F.; Limam, F.; Aouani, E.; Mokni, M. Protective effect of grape seed extract and exercise training on tissues toxicities in doxorubicin-treated healthy rat. Biomarkers 2023, 28, 544–554. [Google Scholar] [CrossRef] [PubMed]
- Monte, M.G.; Tonon, C.R.; Fujimori, A.S.; Ribeiro, A.P.D.; Zanati, S.G.; Okoshi, K.; Camacho, C.R.C.; Moretto, M.R.; de Paiva, S.A.R.; Zornoff, L.A.M.; et al. Omega-3 supplementation attenuates doxorubicin-induced cardiotoxicity but is not related to the ceramide pathway. Food Sci. Nutr. 2024, 12, 9198–9211. [Google Scholar] [CrossRef] [PubMed]
- Tarpey, M.D.; Amorese, A.J.; Balestrieri, N.P.; Fisher-Wellman, K.H.; Spangenburg, E.E. Doxorubicin causes lesions in the electron transport system of skeletal muscle mitochondria that are associated with a loss of contractile function. J. Biol. Chem. 2019, 294, 19709–19722. [Google Scholar] [CrossRef] [PubMed]
- Osama, E.; Khowailed, E.; Rashed, L.; Fawzy, A.; Hassan, R.M.; Harb, I.; Maher, M. Evaluation of skeletal muscle function in male rats with doxorubicin-induced myopathy following various exercise techniques: The significant role of glucose transporter 4. Pflügers Arch.-Eur. J. Physiol. 2024, 476, 797–808. [Google Scholar] [CrossRef] [PubMed]
- Lessard, S.J.; MacDonald, T.L.; Pathak, P.; Han, M.S.; Coffey, V.G.; Edge, J.; Rivas, D.A.; Hirshman, M.F.; Davis, R.J.; Goodyear, L.J. JNK regulates muscle remodeling via myostatin/SMAD inhibition. Nat. Commun. 2018, 9, 3030. [Google Scholar] [CrossRef] [PubMed]
- Krzysztofik, M.; Wilk, M.; Wojdała, G.; Gołaś, A. Maximizing Muscle Hypertrophy: A Systematic Review of Advanced Resistance Training Techniques and Methods. Int. J. Environ. Res. Public Health 2019, 16, 4879. [Google Scholar] [CrossRef] [PubMed]
- Bodine, S.C. The role of mTORC1 in the regulation of skeletal muscle mass. Fac. Rev. 2022, 11, 32. [Google Scholar] [CrossRef] [PubMed]
- Bredahl, E.C.; Busekrus, R.B.; Hydock, D.S. The combined effect of creatine and resistance training on doxorubicin-induced muscle dysfunction. Nutr. Cancer 2020, 72, 939–947. [Google Scholar] [CrossRef] [PubMed]
- Bredahl, E.C.; Sharif, S.; Siedlik, J.A.; Wagner, M.K.; Twaddell, M.D.; Tigner, A.T.; Dovgan, M.D.; Najdawi, W.O.; Hydock, D.S.; Eckerson, J.M.; et al. Resistance Training during Chemotherapy with Doxorubicin. Med. Sci. Sports Exerc. 2020, 52, 2529–2537. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Delafontaine, P. Mechanisms of IGF-1-Mediated Regulation of Skeletal Muscle Hypertrophy and Atrophy. Cells 2020, 9, 1970. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pigg, Q.W.; Harris, D.R.; Inoue, D.S.; Janini Gomes, M. Mitigating Doxorubicin-Induced Skeletal Muscle Toxicity: A Review of Oxidative Stress Mechanisms and the Therapeutic Role of Exercise. Antioxidants 2025, 14, 870. https://doi.org/10.3390/antiox14070870
Pigg QW, Harris DR, Inoue DS, Janini Gomes M. Mitigating Doxorubicin-Induced Skeletal Muscle Toxicity: A Review of Oxidative Stress Mechanisms and the Therapeutic Role of Exercise. Antioxidants. 2025; 14(7):870. https://doi.org/10.3390/antiox14070870
Chicago/Turabian StylePigg, Quinten W., Dillon R. Harris, Daniela Sayuri Inoue, and Mariana Janini Gomes. 2025. "Mitigating Doxorubicin-Induced Skeletal Muscle Toxicity: A Review of Oxidative Stress Mechanisms and the Therapeutic Role of Exercise" Antioxidants 14, no. 7: 870. https://doi.org/10.3390/antiox14070870
APA StylePigg, Q. W., Harris, D. R., Inoue, D. S., & Janini Gomes, M. (2025). Mitigating Doxorubicin-Induced Skeletal Muscle Toxicity: A Review of Oxidative Stress Mechanisms and the Therapeutic Role of Exercise. Antioxidants, 14(7), 870. https://doi.org/10.3390/antiox14070870