
Singlet Oxygen in Food: A Review on Its Formation, Oxidative Damages, Quenchers, and Applications in Preservation


Abstract
1. Introduction
2. Literature Selection and Scope
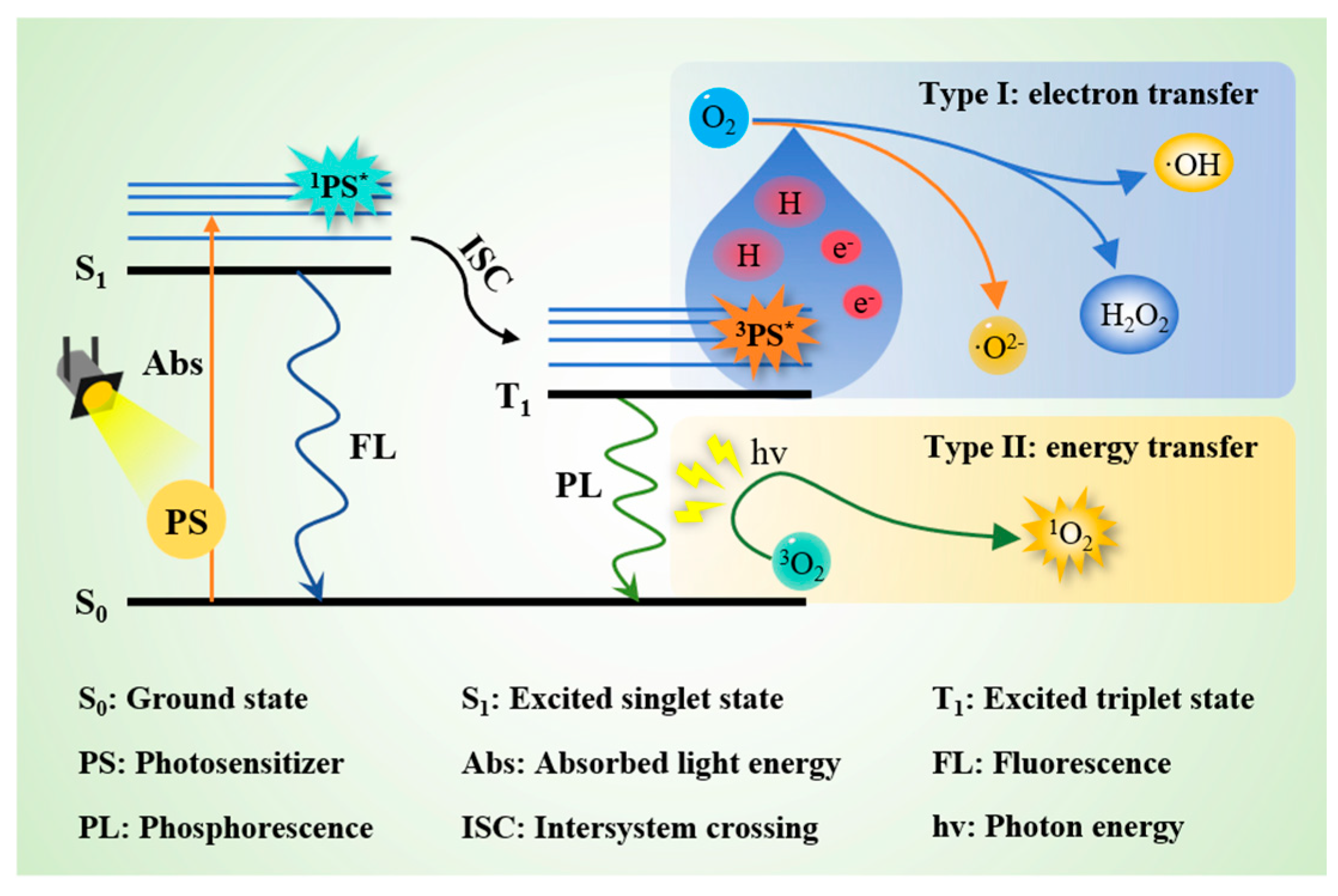
3. Formation Mechanisms of Singlet Oxygen in Nature Food
3.1. Formation Mechanisms of Singlet Oxygen in Vegetables
3.2. Formation Mechanism of Singlet Oxygen in Meats
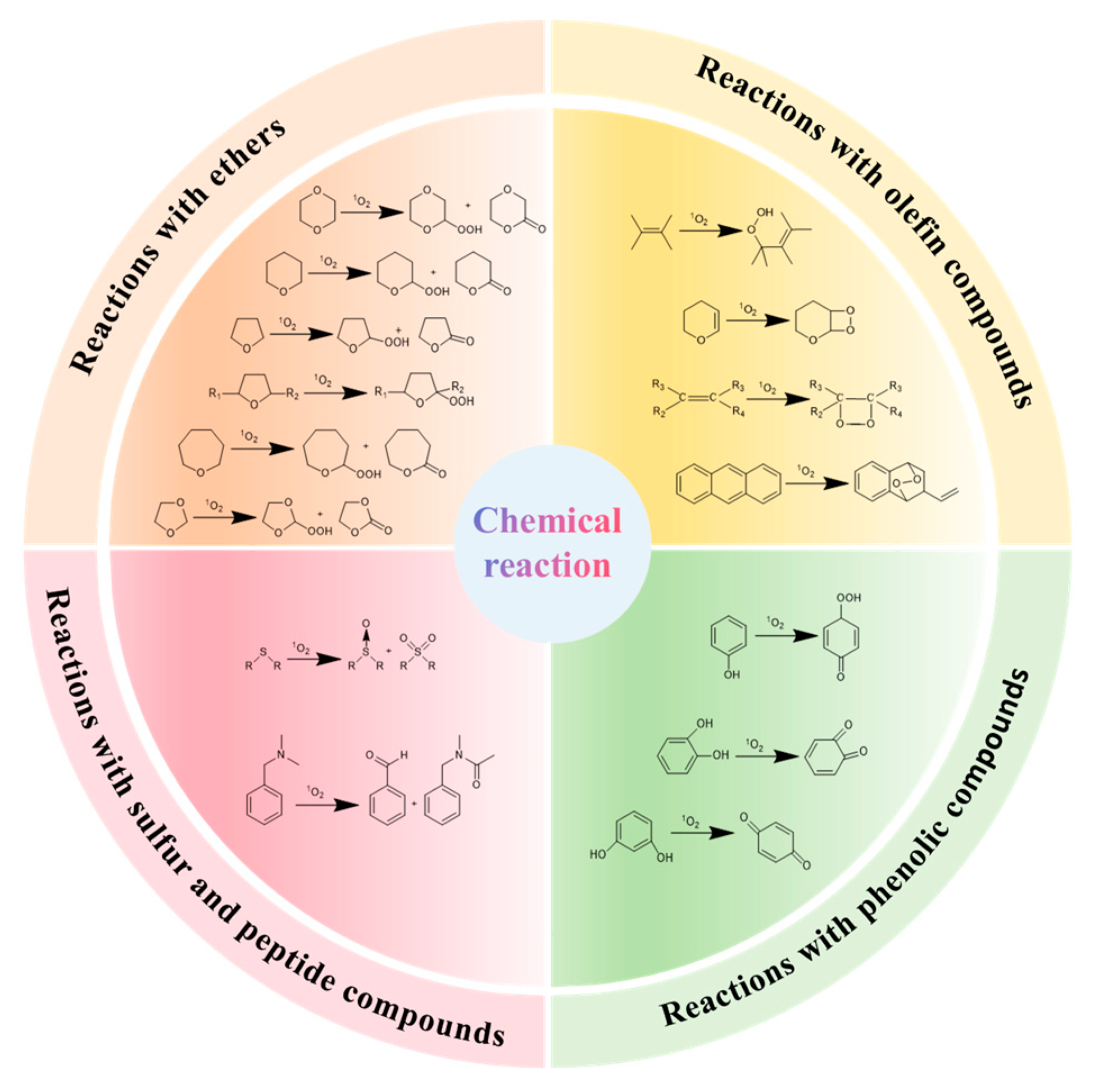
4. Damages of Singlet Oxygen on Food
4.1. Damages of Singlet Oxygen on Protein
4.2. Damages of Singlet Oxygen on Lipid
4.3. Damages of Singlet Oxygen on DNA
5. Self-Protection Mechanisms in Foods Against Singlet Oxygen
5.1. Roles of Singlet Oxygen in Signaling
5.2. Quenchers of Singlet Oxygen in Food
6. Roles of Singlet Oxygen in Food Preservation
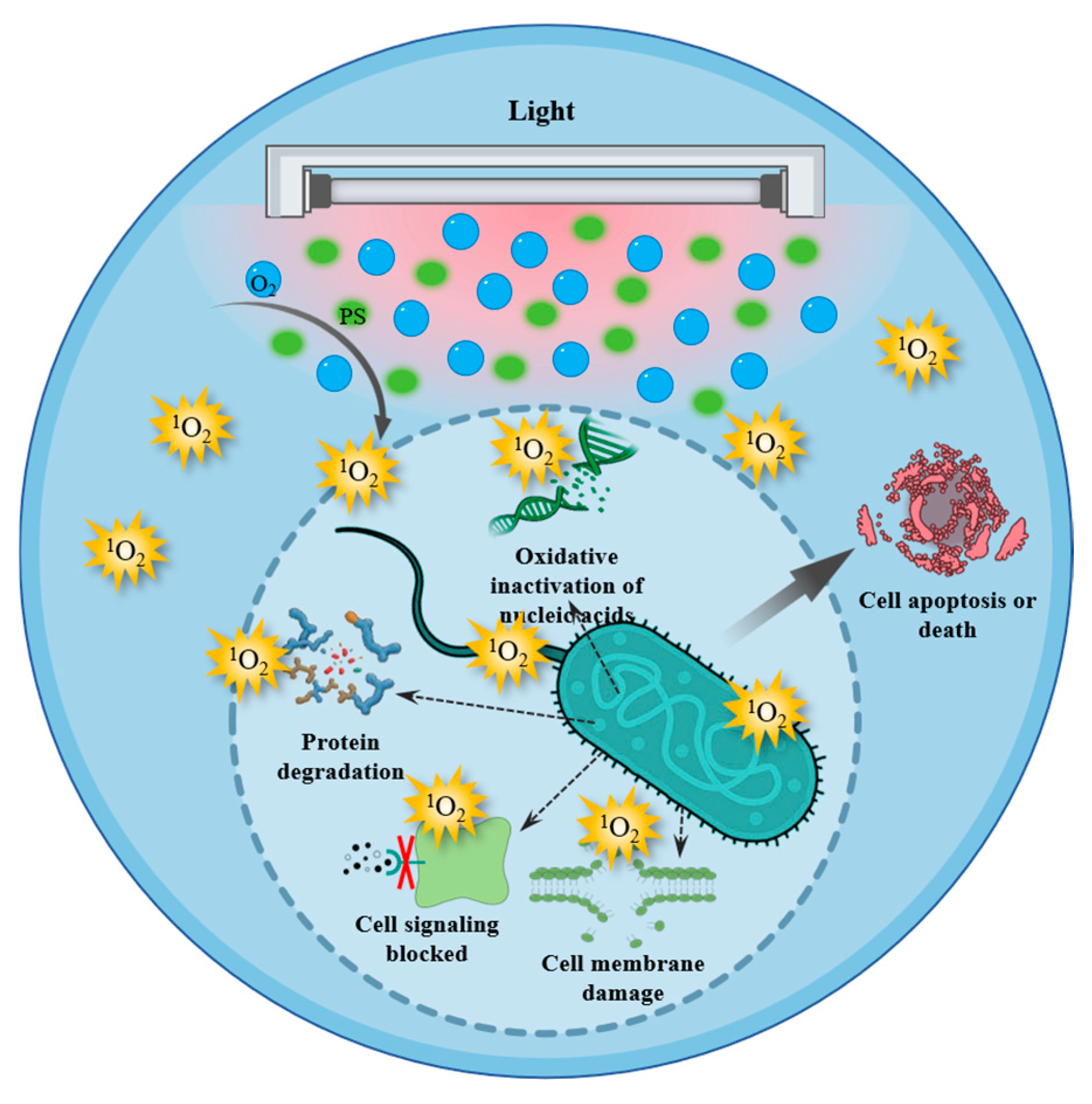
6.1. Antimicrobial Mechanisms of Singlet Oxygen
6.2. Application of Singlet Oxygen in Food Preservation
6.2.1. Photodynamic Technology

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Food Type | Food | Photosensitizer | Light | Wavelength, Energy Density/Irradiance, Time | ΦΔ | Detection Method for ΦΔ | Sensory Quality of Food | References |
|---|---|---|---|---|---|---|---|---|
| Marine products | Oyster | Curcumin | Blue light | 455–460 nm, 9.36 J/cm2, 30 min | 0.11 | UV-Vis/TRF/SSF | PDT treatment can effectively delay the color change of oysters, while maintaining higher hardness and elasticity, a more complete muscle fiber structure, and inhibiting the recovery and proliferation of bacteria, thus enabling oysters to maintain good quality and appearance even after 10 days of storage. | [95,104,117] |
| Tunny | Riboflavin | Blue light | 455 nm, 5.2 mW/cm2, 40 min | 0.54 | NIR | PDT treatment did not affect the contents of TP and TVBN, indicating that the protein quality was retained. With the increase in riboflavin concentration, the number of Salmonella gradually decreased, and the high dose of radiation accelerated the lipid oxidation of tuna. | [4,118] | |
| Fruit and vegetable | Fresh-cut hami melon | Curcumin | Blue light | 460 nm, -, 60 min | 0.11 | UV-Vis/TRF/SSF | This method can significantly reduce the microbial count in the sliced cantaloupe, while also effectively delaying browning and preserving the brightness, hardness, moisture, and soluble solid content of the fruit. Additionally, it ensures that the cantaloupe maintains a good sensory quality even after 9 days of storage. | [94,104,117] |
| Fruit and vegetable | Potatoes | Curcumin | Blue light | 420 nm, 0.7 kJ/cm2, 20 min | 0.11 | UV-Vis/TRF/SSF | The treatment successfully inactivated 2.43 log CFU mL−1 E. coli and 3.18 log CFU mL−1 Staphylococcus aureus. Concurrently, it minimized the loss of phenols and flavonoids and improved the total antioxidant capacity. After being stored for 8 d, the color, elasticity, and chewiness of the potatoes did not change significantly. | [104,117,119] |
| Blueberry | Riboflavin | Blue light | 405 nm, 4.2 mW/cm2, 30 min | 0.54 | NIR | The addition of 0.1% riboflavin or Rose Bengal as a singlet oxygen booster could result in a significant reduction in Tulane virus, with decreases of 0.51 and 1.01 log, respectively. | [4,103,120] | |
| Rose Bengal | 0.76 | |||||||
| Meat product | Beef | Curcumin | Blue light | 450 nm, 55 mW/cm2, 4.8 min | 0.11 | UV-Vis/TRF/SSF | In the sample of beef, chicken, and pork, Staphylococcus aureus count was reduced by 1.5, 1.4, and 0.6 lg mL−1, respectively, without altering their nutritional properties. | [104,117,121] |
| Chicken | ||||||||
| pork | ||||||||
| Dairy product | Milk | Methylene blue | Xenon lamp | 664 nm, 10 mW/cm2, 15 min | 0.57 | FPM | Phthalocyanine-mediated PDT could reduce the number of Staphylococcus aureus in milk by more than 5 logarithmic levels, while the same dosage of methylene blue only achieved a reduction of approximately 1 log. | [105] |
| Silicon (IV) phthalocyanine derivative | 678 nm, 10 mW/cm2, 15 min | 0.59 | ||||||
| Dairy product | Cheese | Riboflavin | Blue light | 460–470 nm; 1 mW/cm2, 7 d | 0.54 | NIR | Treatment at 4 °C can effectively inactivate Listeria monocytogenes and Pseudomonas fluorescens in cheese, with an inactivation amount of 5.14 log CFU/g, and no significant changes occur in sensory quality and color. | [4,28] |
6.2.2. Integration of Packaging Technology and Photodynamic Technology
7. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 1O2 | Singlet oxygen |
| ROS | Reactive oxygen species |
| EPR | Electron paramagnetic resonance |
| Chl | Chlorophyll |
| LHCII | Light-harvesting antenna complexes |
| PSII | Photosystem II |
| 3Chl* | Chlorophyll triplet state |
| ISC | Intersystem crossing |
| 1Chl* | Singlet excited chlorophyll |
| CR | Charge recombination |
| NIR | Near-infrared luminescence method |
| PpIX | Protoporphyrin IX |
| FMN | Flavin mononucleotide |
| FAD | Flavin guanine dinucleotide |
| PUFA | Polyunsaturated fatty acids |
| MDA | Malondialdehyde |
| 4-HNE | 4-hydroxy-2-nonenal |
| BHA | Butylated hydroxyanisole |
| BHT | Butylated hydroxytoluene |
| TBHQ | Tert-butyl-hydroquinone |
| PDT | Photodynamic technology |
| S1 | Excited singlet state |
| T1 | Excited triplet state |
| 3PS* | Tri-excitation photosensitizer |
| GQDs | Graphene quantum dots |
| UV-Vis/TRF/SSF | Ultraviolet-visible spectroscopy with time-resolved/steady-state fluorescence techniques |
| FPM | Fluorescent probe method |
References
- Unsihuay, D.; Su, P.; Hu, H.; Qiu, J.; Kuang, S.; Li, Y.; Sun, X.; Dey, S.K.; Laskin, J. Inside Cover: Imaging and Analysis of Isomeric Unsaturated Lipids through Online Photochemical Derivatization of Carbon–Carbon Double Bonds (Angew. Chem. Int. Ed. 14/2021). Angew. Chem. Int. Ed. 2021, 60, 7450. [Google Scholar] [CrossRef]
- Sari, Y.P.; Santoso, U.; Supriyadi; Raharjo, S. Determination of Singlet Oxygen Quenching Rate and Mechanism of γ-Oryzanol. Heliyon 2021, 7, e07065. [Google Scholar] [CrossRef] [PubMed]
- Fujii, J.; Soma, Y.; Matsuda, Y. Biological Action of Singlet Molecular Oxygen from the Standpoint of Cell Signaling, Injury and Death. Molecules 2023, 28, 4085. [Google Scholar] [CrossRef]
- do Prado-Silva, L.; Brancini, G.T.P.; Braga, G.Ú.L.; Liao, X.; Ding, T.; Sant’Ana, A.S. Antimicrobial Photodynamic Treatment (APDT) as an Innovative Technology to Control Spoilage and Pathogenic Microorganisms in Agri-Food Products: An Updated Review. Food Control. 2022, 132, 108527. [Google Scholar] [CrossRef]
- Temba, B.A.; Fletcher, M.T.; Fox, G.P.; Harvey, J.; Okoth, S.A.; Sultanbawa, Y. Curcumin-Based Photosensitization Inactivates Aspergillus Flavus and Reduces Aflatoxin B1 in Maize Kernels. Food Microbiol. 2019, 82, 82–88. [Google Scholar] [CrossRef]
- Gorji, M.E.; Li, D. Photoinactivation of Bacteriophage MS2, Tulane Virus and Vibrio Parahaemolyticus in Oysters by Microencapsulated Rose Bengal. Food Qual. Saf. 2022, 6, fyac017. [Google Scholar] [CrossRef]
- Munir, Z.; Banche, G.; Cavallo, L.; Mandras, N.; Roana, J.; Pertusio, R.; Ficiarà, E.; Cavalli, R.; Guiot, C. Exploitation of the Antibacterial Properties of Photoactivated Curcumin as ‘Green’ Tool for Food Preservation. Int. J. Mol. Sci. 2022, 23, 2600. [Google Scholar] [CrossRef]
- Steen, C.J.; Niklas, J.; Poluektov, O.G.; Schaller, R.D.; Fleming, G.R.; Utschig, L.M. EPR Spin-Trapping for Monitoring Temporal Dynamics of Singlet Oxygen during Photoprotection in Photosynthesis. Biochemistry 2024, 63, 1214–1224. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, L.; Pan, Y.; Qin, R.; Wang, W.; Zhou, M.; Zhang, Y. Detection Methodologies and Mechanisms of Reactive Oxygen Species Generated in Fenton/Fenton-like Processes. Sep. Purif. Technol. 2025, 355, 129578. [Google Scholar] [CrossRef]
- Nardi, G.; Manet, I.; Monti, S.; Miranda, M.A.; Lhiaubet-Vallet, V. Scope and Limitations of the TEMPO/EPR Method for Singlet Oxygen Detection: The Misleading Role of Electron Transfer. Free Radic. Biol. Med. 2014, 77, 64–70. [Google Scholar] [CrossRef]
- Zaghdoudi, K.; Ngomo, O.; Vanderesse, R.; Arnoux, P.; Myrzakhmetov, B.; Frochot, C.; Guiavarc’h, Y. Extraction, Identification and Photo-Physical Characterization of Persimmon (Diospyros kaki L.) Carotenoids. Foods 2017, 6, 4. [Google Scholar] [CrossRef] [PubMed]
- Edge, R.; Truscott, T.G. Singlet Oxygen and Free Radical Reactions of Retinoids and Carotenoids—A Review. Antioxidants 2018, 7, 5. [Google Scholar] [CrossRef]
- Izuhara, T.; Kaihatsu, I.; Jimbo, H.; Takaichi, S.; Nishiyama, Y. Elevated Levels of Specific Carotenoids During Acclimation to Strong Light Protect the Repair of Photosystem II in Synechocystis Sp. PCC 6803. Front. Plant Sci. 2020, 11, 543740. [Google Scholar] [CrossRef]
- Balevičius, V.; Duffy, C.D.P. Excitation Quenching in Chlorophyll-Carotenoid Antenna Systems: “coherent” or “Incoherent”. Photosynth. Res. 2020, 144, 301–315. [Google Scholar] [CrossRef] [PubMed]
- Mattila, H.; Mishra, S.; Tyystjärvi, T.; Tyystjärvi, E. Singlet Oxygen Production by Photosystem II Is Caused by Misses of the Oxygen Evolving Complex. New Phytol. 2023, 237, 113–125. [Google Scholar] [CrossRef]
- Belyaeva, N.E.; Bulychev, A.A.; Paschenko, V.Z.; Klementiev, K.E.; Ermachenko, P.A.; Konyukhov, I.V.; Riznichenko, G.Y.; Rubin, A.B. Dynamics of In Vivo Membrane Processes in Algal Thylakoids as Analyzed from Chlorophyll Fluorescence Induction Using the Photosystem II and Thylakoid Models. Biophysics 2022, 67, 708–725. [Google Scholar] [CrossRef]
- Rodoni, L.M.; Lemoine, M.L. Shedding Light about the Use of Photosensitizers and Photodynamic Treatments during Postharvest of Fruit and Vegetables: Current and Future State. Postharvest Biol. Technol. 2023, 204, 112463. [Google Scholar] [CrossRef]
- Suzuki, N. Fine Tuning of ROS, Redox and Energy Regulatory Systems Associated with the Functions of Chloroplasts and Mitochondria in Plants under Heat Stress. Int. J. Mol. Sci. 2023, 24, 1356. [Google Scholar] [CrossRef]
- Jan, S.; Singh, R.; Bhardwaj, R.; Ahmad, P.; Kapoor, D. Plant Growth Regulators: A Sustainable Approach to Combat Pesticide Toxicity. 3 Biotech. 2020, 10, 466. [Google Scholar] [CrossRef]
- Prasad, A.; Ferretti, U.; Sedlářová, M.; Pospíšil, P. Singlet Oxygen Production in Chlamydomonas Reinhardtii under Heat Stress OPEN. Sci. Rep. 2016, 6, 20094. [Google Scholar] [CrossRef]
- Vicidomini, C.; Palumbo, R.; Moccia, M.; Roviello, G.N. Oxidative Processes and Xenobiotic Metabolism in Plants: Mechanisms of Defense and Potential Therapeutic Implications. J. Xenobiot. 2024, 14, 1541–1569. [Google Scholar] [CrossRef] [PubMed]
- Hajimohammadi, M.; Sheikh Mahboobi, F.; Wu, H. Suppressing Effect of Flavonoid Compounds on Lipids Photooxidation of Sheep Red Blood Cells and Oleic Acid Photooxidation. Food Sci. 2024, 12, 10405–10411. [Google Scholar] [CrossRef]
- SAJGO, M. The Photooxidation of Myoglobin. Acta Physiol. Acad. Sci. Hung. 1961, 18, 279–281. [Google Scholar]
- Lepeshkevich, S.V.; Stasheuski, A.S.; Parkhats, M.V.; Galievsky, V.A.; Dzhagarov, B.M. Does Photodissociation of Molecular Oxygen from Myoglobin and Hemoglobin Yield Singlet Oxygen? J. Photochem. Photobiol. B 2013, 120, 130–141. [Google Scholar] [CrossRef]
- Vikas, V.; Yang, W.; Wilson, B.C.; Zhu, T.C.; Hadfield, R.H. Analysis of Singlet Oxygen Luminescence Generated By Protoporphyrin IX. Antioxidants 2025, 14, 176. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T.; Hara, K.; Honda, N.; Okazaki, S.; Hazama, H.; Awazu, K. Determination and Analysis of Singlet Oxygen Quantum Yields of Talaporfin Sodium, Protoporphyrin IX, and Lipidated Protoporphyrin IX Using near-Infrared Luminescence Spectroscopy. Lasers Med. Sci. 2020, 35, 1289–1297. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, B.B.; Sestito, J.M.B.; Bezerra, R.A.D.; de Oliveira, G.L.M.; da Silva Júnior, R.C.; Machado, R.R.B.; Nakamura, C.V.; Alfieri, A.A.; dos Santos Pozza, M.S. Reduction of Staphylococcus Aureus in Vitro and in Milk by Photodynamic Inactivation Using Riboflavin and Curcumin as Photosensitizers: Cell Damage and Effects on Product Quality. J. Photochem. Photobiol. A Chem. 2024, 446, 115120. [Google Scholar] [CrossRef]
- Hadi, J.; Wu, S.; Brightwell, G. Antimicrobial Blue Light versus Pathogenic Bacteria: Mechanism, Application in the Food Industry, Hurdle Technologies and Potential Resistance. Foods 2020, 9, 1895. [Google Scholar] [CrossRef]
- Suwannasom, N.; Kao, I.; Pruß, A.; Georgieva, R.; Bäumler, H. Riboflavin: The Health Benefits of a Forgotten Natural Vitamin. Int. J. Mol. Sci. 2020, 21, 950. [Google Scholar] [CrossRef]
- Westberg, M.; Etzerodt, M.; Ogilby, P.R. Rational Design of Genetically Encoded Singlet Oxygen Photosensitizing Proteins. Curr. Opin. Struct. Biol. 2019, 57, 56–62. [Google Scholar] [CrossRef]
- Felčíková, K.; Hovan, A.; Polák, M.; Loginov, D.S.; Holotová, V.; Díaz, C.; Kožár, T.; Lee, O.S.; Varhač, R.; Novák, P.; et al. Design of AsLOV2 Domain as a Carrier of Light-induced Dissociable FMN Photosensitizer. Protein Sci. 2024, 33, e4921. [Google Scholar] [CrossRef] [PubMed]
- Petrenčáková, M.; Filandr, F.; Hovan, A.; Yassaghi, G.; Man, P.; Kožár, T.; Schwer, M.S.; Jancura, D.; Plückthun, A.; Novák, P.; et al. Photoinduced Damage of AsLOV2 Domain Is Accompanied by Increased Singlet Oxygen Production Due to Flavin Dissociation. Sci. Rep. 2020, 10, 4119. [Google Scholar] [CrossRef] [PubMed]
- Insińska-Rak, M.; Henke, P.; Breitenbach, T.; Ogilby, P.R. Singlet Oxygen Quenching by Riboflavin. J. Photochem. Photobiol. A Chem. 2024, 446, 115108. [Google Scholar] [CrossRef]
- Zhoua, Q.; Zhang, X.; Zhou, C. Transformation of Amino Acid Tyrosine in Chromophoric Organic Matter Solutions: Generation of Peroxide and Change of Bioavailability. Chemosphere 2020, 245, 125662. [Google Scholar] [CrossRef]
- Wang, Y.; Lin, Y.; He, S.; Wu, S.; Yang, C. Singlet Oxygen: Properties, Generation, Detection, and Environmental Applications. J. Hazard. Mater. 2024, 461, 132538. [Google Scholar] [CrossRef]
- Knight, K. Mitochondria Are Responsible for Reactive Oxygen Species Damage in Fish Muscle. J. Exp. Biol. 2021, 224, 99–107. [Google Scholar] [CrossRef]
- Wenske, S.; Lackmann, J.W.; Busch, L.M.; Bekeschus, S.; von Woedtke, T.; Wende, K. Reactive Species Driven Oxidative Modifications of Peptides—Tracing Physical Plasma Liquid Chemistry. J. Appl. Phys. 2021, 129, 193305. [Google Scholar] [CrossRef]
- Grassi, L.; Cabrele, C. Susceptibility of Protein Therapeutics to Spontaneous Chemical Modifications by Oxidation, Cyclization, and Elimination Reactions. Amino Acids 2019, 51, 1409–1431. [Google Scholar] [CrossRef]
- Kehm, R.; Baldensperger, T.; Raupbach, J.; Höhn, A. Protein Oxidation—Formation Mechanisms, Detection and Relevance as Biomarkers in Human Diseases. Redox Biol. 2021, 42, 101901. [Google Scholar] [CrossRef]
- Wang, J.; Chen, L.; Liu, Y.; Olajide, T.M.; Jiang, Y.; Cao, W. Identification of Key Aroma-Active Compounds in Beef Tallow Varieties Using Flash GC Electronic Nose and GC × GC-TOF/MS. Eur. Food Res. Technol. 2022, 248, 1733–1747. [Google Scholar] [CrossRef]
- Wang, L.; Deng, W.; Wang, P.; Huang, W.; Zheng, T.; Cai, J.; Wu, J.; Chen, J. Comparative Analyses of Three Sterilization Processes on Volatile Compounds in Sugarcane Juice. Trans. ASABE 2019, 62, 1689–1696. [Google Scholar] [CrossRef]
- Jo, Y.; Carter, B.G.; Barbano, D.M.; Drake, M.A. Identification of the Source of Volatile Sulfur Compounds Produced in Milk during Thermal Processing. J. Dairy. Sci. 2019, 102, 8658–8669. [Google Scholar] [CrossRef] [PubMed]
- Cadwallader, D.C.; Pranata, J.; Liu, Y.; Barbano, D.M.; Drake, M. Effects of Storage Time and Temperature on the Chemical and Sensory Properties of Aseptic Milk. J. Dairy. Sci. 2025, 108, 4818–4838. [Google Scholar] [CrossRef]
- Jayme, S.B.; Prado, F.M.; Massafera, M.P.; Ronsein, G.E.; Di Mascio, P. Characterization and Quantification of Tryptophan- and Tyrosine-Derived Hydroperoxides†. Photochem. Photobiol. 2022, 98, 678–686. [Google Scholar] [CrossRef]
- Domínguez, R.; Pateiro, M.; Munekata, P.E.S.; Zhang, W.; Garcia-Oliveira, P.; Carpena, M.; Prieto, M.A.; Bohrer, B.; Lorenzo, J.M. Protein Oxidation in Muscle Foods: A Comprehensive Review. Antioxidants 2021, 11, 60. [Google Scholar] [CrossRef]
- Kim, S.; Kim, E.; Park, M.; Kim, S.H.; Kim, B.-G.; Na, S.; Sadongo, V.W.; Wijesinghe, W.C.B.; Eom, Y.-G.; Yoon, G.; et al. Hidden Route of Protein Damage through Oxygen-Confined Photooxidation. Nat. Commun. 2024, 15, 10873. [Google Scholar] [CrossRef]
- Maitra, D.; Bragazzi Cunha, J.; Elenbaas, J.S.; Bonkovsky, H.L.; Shavit, J.A.; Omary, M.B. Porphyrin-Induced Protein Oxidation and Aggregation as a Mechanism of Porphyria-Associated Cell Injury. Cmgh 2019, 8, 535–548. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Liu, Y.; Zhou, M.; Shi, X.; Pu, X.; He, Z.; Zhang, S.; Qin, F.; Luo, C. Small Molecule–Engineered Nanoassembly for Lipid Peroxidation–Amplified Photodynamic Therapy. Drug Deliv. Transl. Res. 2024, 14, 1860–1871. [Google Scholar] [CrossRef]
- Shahidi, F.; Hossain, A. Role of Lipids in Food Flavor Generation. Molecules 2022, 27, 5014. [Google Scholar] [CrossRef]
- Sharma, S.; Sharma, P.; Bailey, T.; Bhattarai, S.; Subedi, U.; Miller, C.; Ara, H.; Kidambi, S.; Sun, H.; Panchatcharam, M.; et al. Electrophilic Aldehyde 4-Hydroxy-2-Nonenal Mediated Signaling and Mitochondrial Dysfunction. Biomolecules 2022, 12, 1555. [Google Scholar] [CrossRef]
- Wang, L.; Lin, Z.; Peng, C.; Zhang, H.; Zhang, L.; Zheng, S.; Chen, J. Roles of ROS in Physiological, Microbial and Metabolomic Alterations of Fresh–Cut Sugarcane under Red and Blue Light Irradiation. Food Chem. X 2025, 26, 102344. [Google Scholar] [CrossRef] [PubMed]
- Larsson, K.; Harrysson, H.; Havenaar, R.; Alminger, M.; Undeland, I. Formation of Malondialdehyde (MDA), 4-Hydroxy-2-Hexenal (HHE) and 4-Hydroxy-2-Nonenal (HNE) in Fish and Fish Oil during Dynamic Gastrointestinal in Vitro Digestion. Food Funct. 2016, 7, 1176–1187. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, Z.; Liu, X.; Wu, D.; Ding, Y.; Li, G.; Wu, Y. Typical Reactive Carbonyl Compounds in Food Products: Formation, Influence on Food Quality, and Detection Methods. Compr. Rev. Food Sci. Food Saf. 2020, 19, 503–529. [Google Scholar] [CrossRef] [PubMed]
- Bernerd, F.; Passeron, T.; Castiel, I.; Marionnet, C. The Damaging Effects of Long UVA (UVA1) Rays: A Major Challenge to Preserve Skin Health and Integrity. Int. J. Mol. Sci. 2022, 23, 8243. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, L.; Zhen, W.; Li, S.; Jiang, X. Generation of Singlet Oxygen via Iron-Dependent Lipid Peroxidation and Its Role in Ferroptosis. Fundam. Res. 2022, 2, 66–73. [Google Scholar] [CrossRef]
- Tsouri, A.M.; Robinson, D. Reactions of Singlet Oxygen with Cholesterol: A Computational Study. Mol. Phys. 2023, 121, e2139305. [Google Scholar] [CrossRef]
- Yagura, T.; Schuch, A.P.; Carrião, C.; Garcia, M.; Ribeiro, C.; Rocha, R.; Moreno, N.C.; Pedro, J.; Angeli, F.; Mendes, D.; et al. Direct Participation of DNA in the Formation of Singlet Oxygen and Base Damage under UVA Irradiation. Free. Radic. Biol. Med. 2017, 108, 86–93. [Google Scholar] [CrossRef]
- De Paepe, L.; Madder, A.; Cadoni, E. Exploiting G-Quadruplex-DNA Damage as a Tool to Quantify Singlet Oxygen Production. Small Methods 2024, 8, e2301570. [Google Scholar] [CrossRef]
- Wang, X.; Chen, J.; Hu, H.; Gong, M.; Wu, M.; Ye, B.; Hu, H.; Du, Z.; Liu, A.; Huang, S.; et al. The Resveratrol Attenuates Reactive Oxygen Species Mediated DNA Damage in Cardiac Malformations Caused by 4-Tert-Octylphenol. Toxicol. Appl. Pharmacol. 2025, 498, 117284. [Google Scholar] [CrossRef]
- Skovgaard, N. Safety of Genetically Engineered Foods. Approaches to Assessing Unintended Health Effects. Int. J. Food Microbiol. 2005, 101, 349–350. [Google Scholar] [CrossRef]
- Khedia, J.; Agarwal, P.; Agarwal, P.K. Deciphering Hydrogen Peroxide-Induced Signalling towards Stress Tolerance in Plants. 3 Biotech. 2019, 9, 395. [Google Scholar] [CrossRef]
- Dogra, V.; Kim, C. Singlet Oxygen Metabolism: From Genesis to Signaling. Front. Plant Sci. 2020, 10, 500205. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Zandalinas, S.I.; Fichman, Y.; Van Breusegem, F. Reactive Oxygen Species Signalling in Plant Stress Responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 663–679. [Google Scholar] [CrossRef]
- Dmitrieva, V.A.; Tyutereva, E.V.; Voitsekhovskaja, O.V. Singlet Oxygen in Plants: Generation, Detection, and Signaling Roles. Int. J. Mol. Sci. 2020, 21, 3237. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Li, L.; Zhang, Y.; Gao, H.; Zhao, Y.; Yu, X. Chemical Inducers Regulate ROS Signalling to Stimulate Astaxanthin Production in Haematococcus Pluvialis under Environmental Stresses: A Review. Trends Food Sci. 2023, 136, 181–193. [Google Scholar] [CrossRef]
- Hong, Y.; Yu, Z.; Zhou, Q.; Chen, C.; Hao, Y.; Wang, Z.; Zhu, J.K.; Guo, H.; Huang, A.C. NAD+ Deficiency Primes Defense Metabolism via 1O2-Escalated Jasmonate Biosynthesis in Plants. Nat. Commun. 2024, 15, 6652. [Google Scholar] [CrossRef]
- Fredimoses, M.; Li, P.; Zhang, Y.; Jia, H.; Liu, S.; Tian, J.; Nie, W.; Liu, K.; Song, M.; Dong, Z. Design, Synthesis, and Antiproliferative Activity Evaluation of Novel α-Mangostin Derivatives by ROS/MAPK Signaling Pathway. Bioorganic Chem. 2024, 153, 107968. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Huang, J.; Pei, S.; Ouyang, Y.; Ding, Y.; Jiang, L.; Lu, J.; Kang, L.; Huang, L.; Xiang, H.; et al. Ganoderma Lucidum Polysaccharide Inhibits UVB-Induced Melanogenesis by Antagonizing CAMP/PKA and ROS/MAPK Signaling Pathways. J. Cell Physiol. 2018, 234, 7330–7340. [Google Scholar] [CrossRef]
- Mohanta, T.K.; Bashir, T.; Hashem, A.; Abd Allah, E.F.; Khan, A.L.; Al-Harrasi, A.S. Early Events in Plant Abiotic Stress Signaling: Interplay Between Calcium, Reactive Oxygen Species and Phytohormones. J. Plant Growth Regul. 2018, 37, 1033–1049. [Google Scholar] [CrossRef]
- Li, M.; Park, B.M.; Dai, X.; Xu, Y.; Huang, J.; Sun, F. Controlling Synthetic Membraneless Organelles by a Red-Light-Dependent Singlet Oxygen-Generating Protein. Nat. Commun. 2022, 13, 3197. [Google Scholar] [CrossRef]
- Przygoda, M.; Bartusik-Aebisher, D.; Dynarowicz, K.; Cieślar, G.; Kawczyk-Krupka, A.; Aebisher, D. Cellular Mechanisms of Singlet Oxygen in Photodynamic Therapy. Int. J. Mol. Sci. 2023, 24, 16890. [Google Scholar] [CrossRef] [PubMed]
- Kruk, J.; Szymańska, R. Singlet Oxygen Oxidation Products of Carotenoids, Fatty Acids and Phenolic Prenyllipids. J. Photochem. Photobiol. B 2021, 216, 112148. [Google Scholar] [CrossRef]
- Elgendey, F.; Al Wakeel, R.A.; Hemeda, S.A.; Elshwash, A.M.; Fadl, S.E.; Abdelazim, A.M.; Alhujaily, M.; Khalifa, O.A. Selenium and/or Vitamin E Upregulate the Antioxidant Gene Expression and Parameters in Broilers. BMC Vet. Res. 2022, 18, 310. [Google Scholar] [CrossRef] [PubMed]
- Mititelu, M.; Lupuliasa, D.; Neacșu, S.M.; Olteanu, G.; Busnatu, Ș.S.; Mihai, A.; Popovici, V.; Măru, N.; Boroghină, S.C.; Mihai, S.; et al. Polyunsaturated Fatty Acids and Human Health: A Key to Modern Nutritional Balance in Association with Polyphenolic Compounds from Food Sources. Foods 2024, 14, 46. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Tang, M.; Wang, L.; Chu, W.; Shi, W.; Zhou, Q.; Pan, Y. How to Achieve Adequate Quenching for DBP Analysis in Drinking Water? Water Res. 2024, 253, 121264. [Google Scholar] [CrossRef]
- Salim Akhter, M.; Noreen, S.; Mahmood, S.; Athar, H.-U.-R.; Ashraf, M.; Abdullah Alsahli, A.; Ahmad, P. Influence of Salinity Stress on PSII in Barley (Hordeum vulgare L.) Genotypes, Probed by Chlorophyll-a Fluorescence. J. King Saud Univ.-Sci. 2021, 33, 101239. [Google Scholar] [CrossRef]
- Zbyradowski, M.; Duda, M.; Wisniewska-Becker, A.; Heriyanto; Rajwa, W.; Fiedor, J.; Cvetkovic, D.; Pilch, M.; Fiedor, L. Triplet-Driven Chemical Reactivity of β-Carotene and Its Biological Implications. Nat. Commun. 2022, 13, 2474. [Google Scholar] [CrossRef] [PubMed]
- Mukai, K. Antioxidant Activity of Foods: Development of Singlet Oxygen Absorption Capacity (SOAC) Assay Method. J. Nutr. Sci. Vitaminol. 2019, 65, 285–302. [Google Scholar] [CrossRef]
- Lee, J.H.; Jung, M.Y. Direct Spectroscopic Observation of Singlet Oxygen Quenching and Kinetic Studies of Physical and Chemical Singlet Oxygen Quenching Rate Constants of Synthetic Antioxidants (BHA, BHT, and TBHQ) in Methanol. J. Food Sci. 2010, 75, C506–C513. [Google Scholar] [CrossRef]
- Zheng, X.; Huang, Q. Assessment of the Antioxidant Activities of Representative Optical and Geometric Isomers of Astaxanthin against Singlet Oxygen in Solution by a Spectroscopic Approach. Food Chem. 2022, 395, 133584. [Google Scholar] [CrossRef]
- Li, X.; Cai, X.; Zeng, Q.; Ren, X.; Kong, Q. Exploring the Mechanism of Stilbenes to Quench Singlet Oxygen Based on the Key Structures of Resveratrol and Its Analogues. Food Chem. 2023, 403, 134350. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.Y.; Choi, D.S. Electron Spin Resonance and Luminescence Spectroscopic Observation and Kinetic Study of Chemical and Physical Singlet Oxygen Quenching by Resveratrol in Methanol. J. Agric. Food Chem. 2010, 58, 11888–11895. [Google Scholar] [CrossRef]
- Lund, M.N. Reactions of Plant Polyphenols in Foods: Impact of Molecular Structure. Trends Food Sci. Technol. 2021, 112, 241–251. [Google Scholar] [CrossRef]
- Tournaire, C.; Croux, S.; Maurette, M.T.; Beck, I.; Hocquaux, M.; Braun, A.M.; Oliveros, E. Antioxidant Activity of Flavonoids: Efficiency of Singlet Oxygen (1Δg) Quenching. J. Photochem. Photobiol. B 1993, 19, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Lian, P.; Wang, K.; Liu, H.; Li, R.; Li, M.; Bao, X.; Wan, X. Reacting Molecular Oxygen with Butanone under Visible Light Irradiation: A General Aerobic Oxidation of Alkenes, Sulfides, Phosphines, and Silanes. Org. Lett. 2023, 25, 7984–7989. [Google Scholar] [CrossRef]
- Skolia, E.; Gkizis, P.L.; Kokotos, C.G. Aerobic Photocatalysis: Oxidation of Sulfides to Sulfoxides. Chempluschem 2022, 87, e202200008. [Google Scholar] [CrossRef]
- Petrou, A.L.; Petrou, P.L.; Ntanos, T.; Liapis, A. A Possible Role for Singlet Oxygen in the Degradation of Various Antioxidants. A Meta-Analysis and Review of Literature Data. Antioxidants 2018, 7, 35. [Google Scholar] [CrossRef]
- Gulcin, İ. Antioxidants: A Comprehensive Review. Arch. Toxicol. 2025, 99, 1893–1997. [Google Scholar] [CrossRef]
- Oyim, J.; Matshitse, R.; Malomane, N.; Openda, Y.I.; Nyokong, T.; Managa, M. In Vitro Photoinactivation of S. Aureus and E. Coli Using 5,10,15,20-Tetrakis [4-(Benzyloxy) Phenyl] Porphyrin and Its Metal Derivatives Conjugated to Pristine Graphene Quantum Dots. J. Porphyr. Phthalocyanines 2023, 27, 634–644. [Google Scholar] [CrossRef]
- Vanhaelewyn, L.; Van Der Straeten, D.; De Coninck, B.; Vandenbussche, F. Ultraviolet Radiation From a Plant Perspective: The Plant-Microorganism Context. Front. Plant Sci. 2020, 11, 597642. [Google Scholar] [CrossRef]
- Mao, W.; Qin, J.; Li, M.; Chen, M.; Fu, W.; Ma, Z.; Chen, J.; Fan, T.; Long, Y.; Ma, J. La0.4Sr0.6CoO3-Catalyzed Selective Oxidation of Ethylbenzene to Acetophenone without Solvent: A New Reactive Oxygen Species Transformation Mechanism Mediated by •O2– Derived from 1O2. ACS Sustain. Chem. 2024, 12, 17556–17565. [Google Scholar] [CrossRef]
- Shi, H.; Wu, T.; Duan, M.; Yu, J.; Liu, M.; Wen, X.; Wang, L.; Xu, Y. Electrocatalytic Generation of Singlet Oxygen via ROS-Mediated Redox Chain Reaction for Efficient Disinfection. Nano Lett. 2024, 24, 6939–6947. [Google Scholar] [CrossRef] [PubMed]
- Urbaniak, S.K.; Boguszewska, K.; Szewczuk, M.; Kázmierczak-Barańska, J.; Karwowski, B.T. 8-Oxo-7,8-Dihydro-2′-Deoxyguanosine (8-OxodG) and 8-Hydroxy-2′-Deoxyguanosine (8-OHdG) as a Potential Biomarker for Gestational Diabetes Mellitus (GDM) Development. Molecules 2020, 25, 202. [Google Scholar] [CrossRef]
- Lin, Y.; Hu, J.; Li, S.; Hamzah, S.S.; Jiang, H.; Zhou, A.; Zeng, S.; Lin, S. Curcumin-Based Photodynamic Sterilization for Preservation of Fresh-Cut Hami Melon. Molecules 2019, 24, 2374. [Google Scholar] [CrossRef]
- Chen, B.; Huang, J.; Liu, Y.; Liu, H.; Zhao, Y.; Wang, J.J. Effects of the Curcumin-Mediated Photodynamic Inactivation on the Quality of Cooked Oysters with Vibrio Parahaemolyticus during Storage at Different Temperature. Int. J. Food Microbiol. 2021, 345, 109152. [Google Scholar] [CrossRef]
- Szewczyk, G.; Mokrzyński, K.; Sarna, T. Generation of Singlet Oxygen inside Living Cells: Correlation between Phosphorescence Decay Lifetime, Localization and Outcome of Photodynamic Action. Photochemical 2024, 23, 1673–1685. [Google Scholar] [CrossRef]
- Davis, S.J.; Zhao, Y.; Yu, T.C.; Maytin, E.V.; Anand, S.; Hasan, T.; Pogue, B.W. Singlet Molecular Oxygen: From COIL Lasers to Photodynamic Cancer Therapy. J. Phys. Chem. B 2023, 127, 2289–2301. [Google Scholar] [CrossRef]
- Lu, X.; Zhou, X.; Qiu, W.; Wang, Z.; Cheng, H.; Zhang, H.; Yu, J.; Wang, D.; Ma, J. Singlet Oxygen Involved Electrochemical Disinfection by Anodic Oxidation of H2O2 in the Presence of Cl−. Chem. Eng. J. 2022, 446, 136871. [Google Scholar] [CrossRef]
- Leanse, L.G.; Marasini, S.; dos Anjos, C.; Dai, T. Antimicrobial Resistance: Is There a ‘Light’ at the End of the Tunnel? Antibiotics 2023, 12, 1437. [Google Scholar] [CrossRef]
- Kou, M.; Qin, F.; Lü, W.; Hu, Z.; Zhang, Z. A New Insight into the Singlet Oxygen Mechanism for Photodynamic Therapy. J. Phys. Chem. Lett. 2023, 14, 5613–5617. [Google Scholar] [CrossRef]
- Tang, Y.; Li, Y.; Li, B.; Song, W.; Qi, G.; Tian, J.; Huang, W.; Fan, Q.; Liu, B. Oxygen-Independent Organic Photosensitizer with Ultralow-Power NIR Photoexcitation for Tumor-Specific Photodynamic Therapy. Nat. Commun. 2024, 15, 2530. [Google Scholar] [CrossRef] [PubMed]
- Sheng, L.; Li, X.; Wang, L. Photodynamic Inactivation in Food Systems: A Review of Its Application, Mechanisms, and Future Perspective. Trends Food Sci. Technol. 2022, 124, 167–181. [Google Scholar] [CrossRef]
- Szewczyk, G.; Mokrzyński, K. Concentration-Dependent Photoproduction of Singlet Oxygen by Common Photosensitizers. Molecules 2025, 30, 1130. [Google Scholar] [CrossRef] [PubMed]
- Chignell, C.F.; Bilskj, P.; Reszka, K.J.; Motten, A.G.; Sik, R.H.; Dahl, T.A. Spectral and photochemical properties of curcumin. Photochem. Photobiol. 1994, 59, 295–302. [Google Scholar] [CrossRef]
- Galstyan, A.; Dobrindt, U. Determining and Unravelling Origins of Reduced Photoinactivation Efficacy of Bacteria in Milk. J. Photochem. Photobiol. B 2019, 197, 111554. [Google Scholar] [CrossRef]
- Lutkus, L.V.; Rickenbach, S.S.; McCormick, T.M. Singlet Oxygen Quantum Yields Determined by Oxygen Consumption. J. Photochem. Photobiol. A Chem. 2019, 378, 131–135. [Google Scholar] [CrossRef]
- Galstyan, A. Turning Photons into Drugs: Phthalocyanine-Based Photosensitizers as Efficient Photoantimicrobials. Chem.—A Eur. J. 2020, 27, 1903–1920. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Sun, Y.; Zhou, B.; Zhu, G.; Chen, X.; Qi, Y.; Wang, K. Photosensitization of Riboflavin Reduces the Susceptibility to Gray Mold in Postharvest Kiwifruit. Postharvest Biol. Technol. 2024, 212, 112836. [Google Scholar] [CrossRef]
- Izquierdo, N.; Gamez, E.; Alejo, T.; Mendoza, G.; Arruebo, M. Antimicrobial Photodynamic Therapy Using Encapsulated Protoporphyrin IX for the Treatment of Bacterial Pathogens. Materials 2024, 17, 1717. [Google Scholar] [CrossRef]
- Nguyen, K.U.; Zhang, Y.; Liu, Q.; Zhang, R.; Jin, X.; Taniguchi, M.; Miller, E.S.; Lindsey, J.S. Tolyporphins–Exotic Tetrapyrrole Pigments in a Cyanobacterium—A Review. Molecules 2023, 28, 6132. [Google Scholar] [CrossRef]
- Zhou, X.; Ying, X.; Wu, L.; Liu, L.; Wang, Y.; He, Y.; Han, M. Research Progress of Natural Product Photosensitizers in Photodynamic Therapy. Planta Med. 2024, 90, 368–379. [Google Scholar] [CrossRef] [PubMed]
- Uliana, M.P.; da Cruz Rodrigues, A.; Ono, B.A.; Pratavieira, S.; de Oliveira, K.T.; Kurachi, C. Photodynamic Inactivation of Microorganisms Using Semisynthetic Chlorophyll a Derivatives as Photosensitizers. Molecules 2022, 27, 5769. [Google Scholar] [CrossRef] [PubMed]
- Zarepour, A.; Khosravi, A.; Yücel Ayten, N.; Çakır Hatır, P.; Iravani, S.; Zarrabi, A. Innovative Approaches for Cancer Treatment: Graphene Quantum Dots for Photodynamic and Photothermal Therapies. J. Mater. Chem. B 2024, 12, 4307–4334. [Google Scholar] [CrossRef]
- Horiuchi, H.; Isogai, M.; Hirakawa, K.; Okutsu, T. Improvement of the ON/OFF Switching Performance of a PH-Activatable Porphyrin Derivative by the Introduction of Phosphorus(V). ChemPhotoChem 2019, 3, 138–144. [Google Scholar] [CrossRef]
- Yonemura, H.; Motomura, K. Efficient Generation of Singlet Oxygen Due to Localized Surface Plasmon Resonance of Silver Nanoparticles in Rose Bengal–Silver Nanoparticle Composite Films. Jpn. J. Appl. Phys. 2023, 62, 065001. [Google Scholar] [CrossRef]
- Tamtaji, M.; Guo, X.; Tyagi, A.; Galligan, P.R.; Liu, Z.; Roxas, A.; Liu, H.; Cai, Y.; Wong, H.; Zeng, L.; et al. Machine Learning-Aided Design of Gold Core–Shell Nanocatalysts toward Enhanced and Selective Photooxygenation. ACS Appl. Mater. 2022, 14, 46471–46480. [Google Scholar] [CrossRef]
- de França, B.M.; Oliveira, S.S.C.; Souza, L.O.P.; Mello, T.P.; Santos, A.L.S.; Bello Forero, J.S. Synthesis and Photophysical Properties of Metal Complexes of Curcumin Dyes: Solvatochromism, Acidochromism, and Photoactivity. Dye. Pigment. 2022, 198, 110011. [Google Scholar] [CrossRef]
- Li, H.; Tan, L.; Chen, B.; Huang, J.; Zeng, Q.; Liu, H.; Zhao, Y.; Wang, J.J. Antibacterial Potency of Riboflavin-Mediated Photodynamic Inactivation against Salmonella and Its Influences on Tuna Quality. LWT 2021, 146, 111462. [Google Scholar] [CrossRef]
- Yu, J.; Zhang, F.; Zhang, J.; Han, Q.; Song, L.; Meng, X. Effect of Photodynamic Treatments on Quality and Antioxidant Properties of Fresh-Cut Potatoes. Food Chem. 2021, 362, 130224. [Google Scholar] [CrossRef]
- Kingsley, D.H.; Perez-Perez, R.E.; Boyd, G.; Sites, J.; Niemira, B.A. Evaluation of 405-Nm Monochromatic Light for Inactivation of Tulane Virus on Blueberry Surfaces. J. Appl. Microbiol. 2018, 124, 1017–1022. [Google Scholar] [CrossRef]
- Corrêa, T.Q.; Blanco, K.C.; Garcia, É.B.; Perez, S.M.L.; Chianfrone, D.J.; Morais, V.S.; Bagnato, V.S. Effects of Ultraviolet Light and Curcumin-Mediated Photodynamic Inactivation on Microbiological Food Safety: A Study in Meat and Fruit. Photodiagnosis Photodyn. Ther. 2020, 30, 101678. [Google Scholar] [CrossRef]
- Ma, T.; Chen, Y.; Zhi, X.; Du, B. Cellulose Laurate Films Containing Curcumin as Photoinduced Antibacterial Agent for Meat Preservation. Int. J. Biol. Macromol. 2021, 193, 1986–1995. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Hu, X.; Zou, L.; Ren, T. Preparation of Photosensitive Antibacterial Nanocomposite Film and Its Preservation Effect on Pork Loin. LWT 2024, 191, 115586. [Google Scholar] [CrossRef]
- Su, L.; Huang, J.; Li, H.; Pan, Y.; Zhu, B.; Zhao, Y.; Liu, H. Chitosan-Riboflavin Composite Film Based on Photodynamic Inactivation Technology for Antibacterial Food Packaging. Int. J. Biol. Macromol. 2021, 172, 231–240. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, L.; Zheng, S.; Lin, Z.; Zhang, C.; Zhang, H.; Chen, J.; Wang, L. Singlet Oxygen in Food: A Review on Its Formation, Oxidative Damages, Quenchers, and Applications in Preservation. Antioxidants 2025, 14, 865. https://doi.org/10.3390/antiox14070865
Xiao L, Zheng S, Lin Z, Zhang C, Zhang H, Chen J, Wang L. Singlet Oxygen in Food: A Review on Its Formation, Oxidative Damages, Quenchers, and Applications in Preservation. Antioxidants. 2025; 14(7):865. https://doi.org/10.3390/antiox14070865
Chicago/Turabian StyleXiao, Limei, Shoujing Zheng, Zhengrong Lin, Chunyan Zhang, Hua Zhang, Jiebo Chen, and Lu Wang. 2025. "Singlet Oxygen in Food: A Review on Its Formation, Oxidative Damages, Quenchers, and Applications in Preservation" Antioxidants 14, no. 7: 865. https://doi.org/10.3390/antiox14070865
APA StyleXiao, L., Zheng, S., Lin, Z., Zhang, C., Zhang, H., Chen, J., & Wang, L. (2025). Singlet Oxygen in Food: A Review on Its Formation, Oxidative Damages, Quenchers, and Applications in Preservation. Antioxidants, 14(7), 865. https://doi.org/10.3390/antiox14070865