
Selenomethionine Counteracts T-2 Toxin-Induced Liver Injury by Mitigating Oxidative Stress Damage Through the Enhancement of Antioxidant Enzymes

 ,
,
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Animals and Experimental Design
2.3. Sample Collection and Preparation
2.4. Histopathological Analysis
2.5. Serum Biochemical Analysis
2.6. Hepatic Biochemical Analysis
2.7. ROS Content Analysis
2.8. Real-Time qPCR Analysis
2.9. Immunohistochemistry Analysis
2.10. Statistical Analysis
3. Results
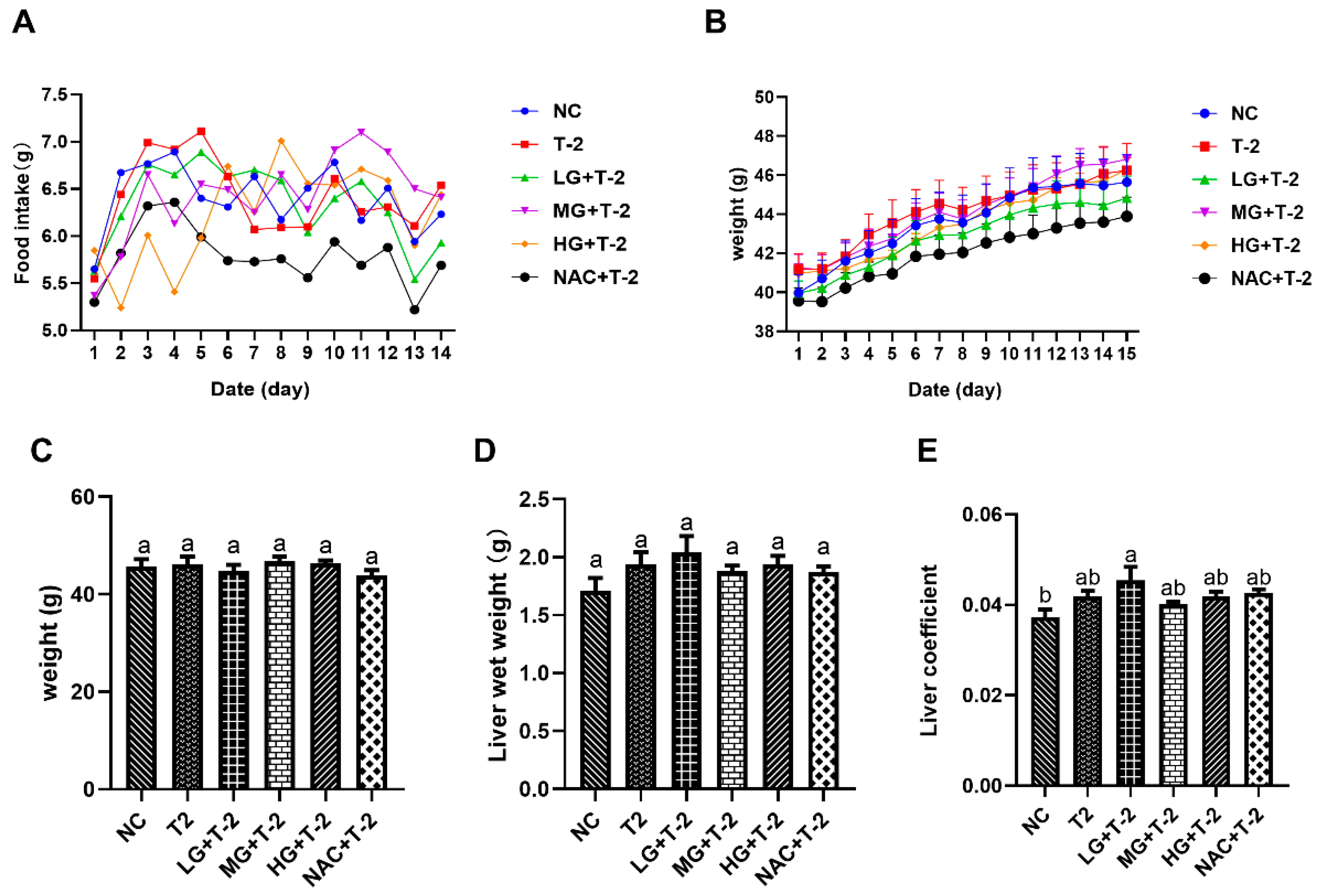
3.1. Effects of SeMet on Food Intake, Body Weight, and Liver Coefficient of T-2 Toxin-Exposed Mice
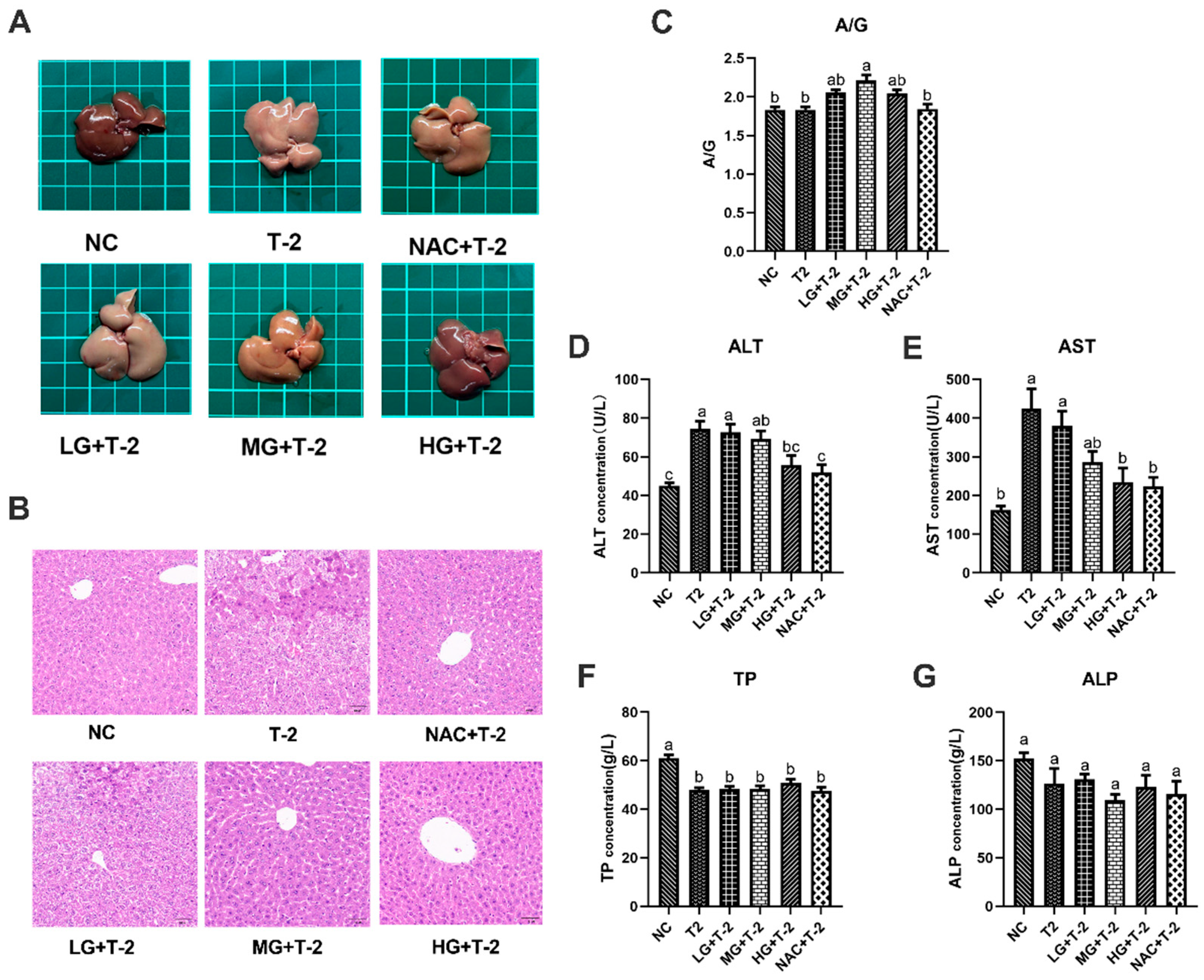
3.2. Protective Effects of SeMet on Liver Morphology and Function in T-2 Toxin-Exposed Mice
3.3. Effects of SeMet on T-2 Toxin-Induced Oxidative Stress in Mouse Livers
3.4. Modulation of Inflammation and Oxidative Stress-Related Gene Expression by SeMet in T-2 Toxin-Exposed Mice Livers
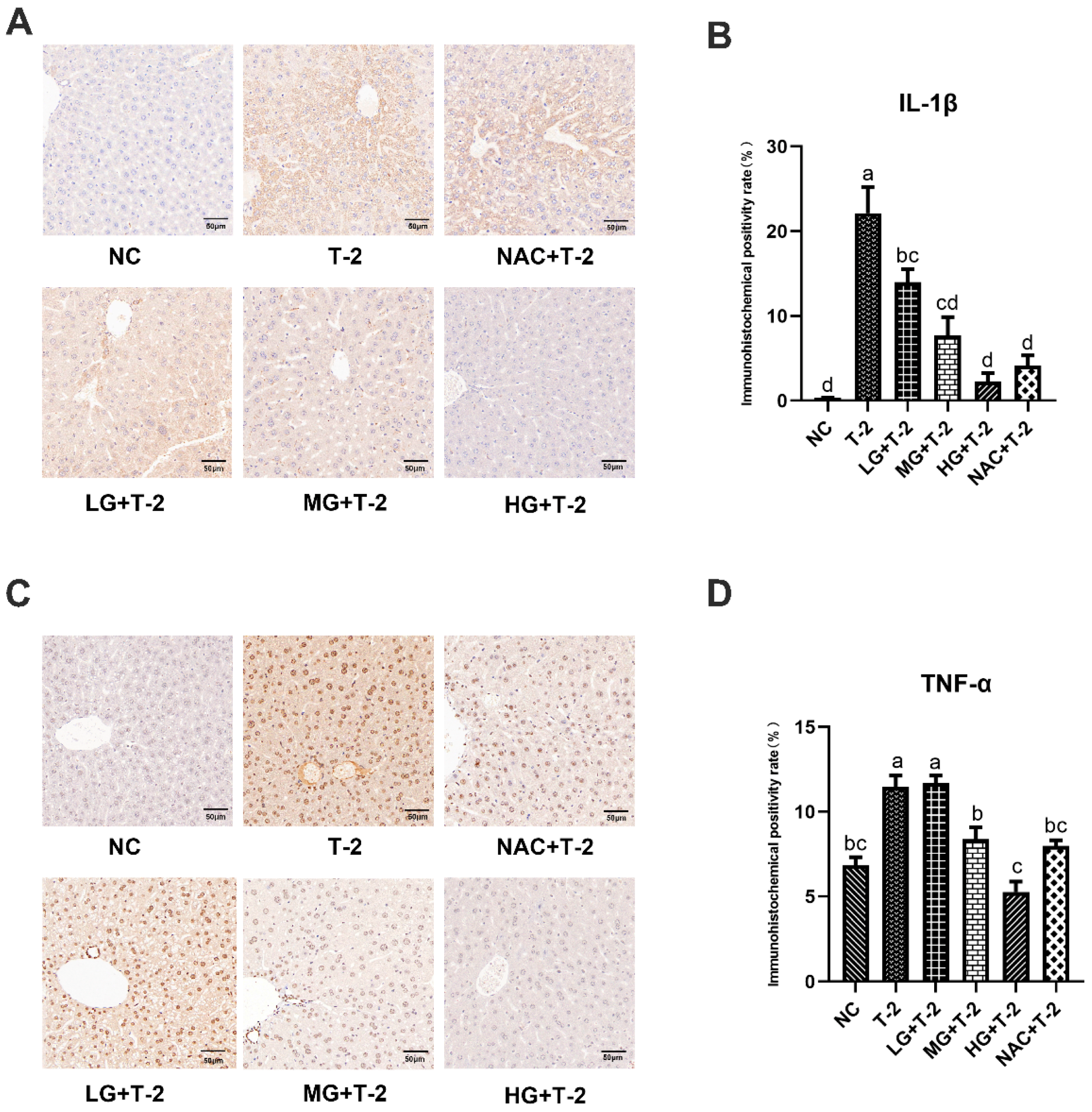
3.5. Impact of SeMet on IL-1β and TNF-α Expression in the Livers of T-2 Toxin-Exposed Mice
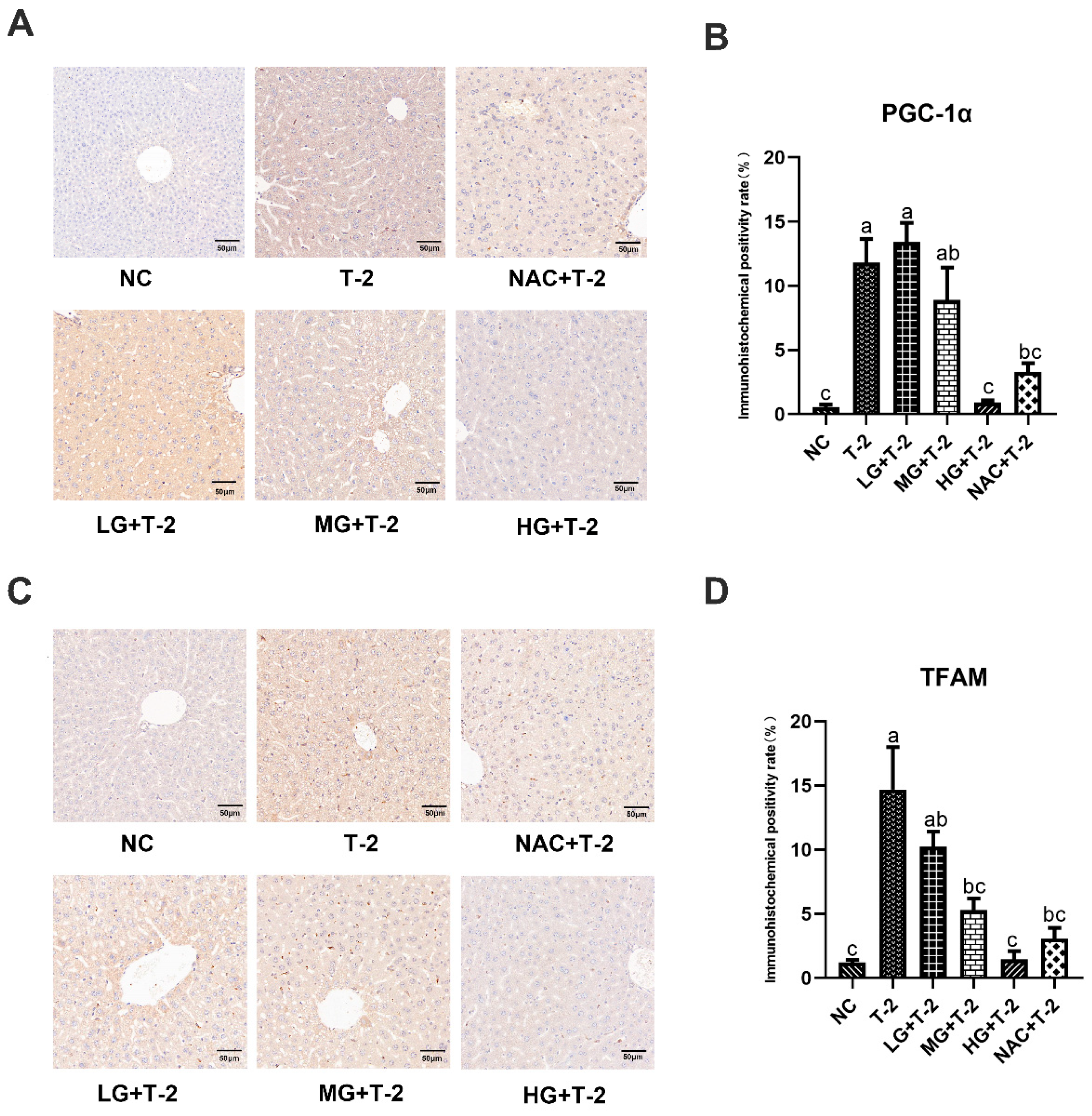
3.6. Effect of SeMet on the Immunohistochemical Expression of Mitochondrial Homeostasis-Related Proteins in the Livers of T-2 Toxin-Induced Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Song, C.; Wang, Z.; Cao, J.; Dong, Y.; Chen, Y. Neurotoxic mechanisms of mycotoxins: Focus on aflatoxin B1 and T-2 toxin. Environ. Pollut. 2024, 356, 124359. [Google Scholar] [CrossRef]
- Ueno, Y. Toxicological features of T-2 toxin and related trichothecenes. Fundam. Appl. Toxicol. 1984, 4, S124–S132. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Wang, Y.; Huang, T.; Liu, Y.; Chen, F.; Chen, Y.; Jiang, Y.; Zhang, C.; Yang, X. T-2 toxin metabolism and its hepatotoxicity: New insights on the molecular mechanism and detoxification. Environ. Pollut. 2023, 330, 121784. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Sun, L.H.; Wang, X.; Wu, Q.; Liu, A. Effective protective agents against the organ toxicity of T-2 toxin and corresponding detoxification mechanisms: A narrative review. Anim. Nutr. 2024, 16, 251–266. [Google Scholar] [CrossRef]
- Dai, C.; Xiao, X.; Sun, F.; Zhang, Y.; Hoyer, D.; Shen, J.; Tang, S.; Velkov, T. T-2 toxin neurotoxicity: Role of oxidative stress and mitochondrial dysfunction. Arch. Toxicol. 2019, 93, 3041–3056. [Google Scholar] [CrossRef]
- He, J.; Jin, H.; Guo, J.; Li, K.; Jia, L.; Li, Y.; Zhang, L. T-2 toxin-induced testicular impairment by triggering oxidative stress and ferroptosis. Ecotoxicol. Environ. Saf. 2024, 270, 115844. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Song, W.; Hua, Z.; Du, J.; Lucena, R.B.; Wang, X.; Zhang, C.; Yang, X. Overview of T-2 Toxin Enterotoxicity: From Toxic Mechanisms and Detoxification to Future Perspectives. J. Agric. Food Chem. 2024, 72, 3314–3324. [Google Scholar] [CrossRef]
- An, K.; Shi, B.; Lv, X.; Liu, Y.; Xia, Z. T-2 toxin triggers lipid metabolism disorder and oxidative stress in liver of ducks. Ecotoxicol. Environ. Saf. 2024, 286, 117169. [Google Scholar] [CrossRef]
- Yang, L.; Yu, Z.; Hou, J.; Deng, Y.; Zhou, Z.; Zhao, Z.; Cui, J. Toxicity and oxidative stress induced by T-2 toxin and HT-2 toxin in broilers and broiler hepatocytes. Food Chem. Toxicol. 2016, 87, 128–137. [Google Scholar] [CrossRef]
- Pecoraro, B.M.; Leal, D.F.; Frias-De-Diego, A.; Browning, M.; Odle, J.; Crisci, E. The health benefits of selenium in food animals: A review. J. Anim. Sci. Biotechnol. 2022, 13, 58. [Google Scholar] [CrossRef]
- Surai, P.F.; Kochish, I.I. Nutritional modulation of the antioxidant capacities in poultry: The case of selenium. Poult. Sci. 2019, 98, 4231–4239. [Google Scholar] [CrossRef]
- Edens, F.W.; Sefton, A.E. Organic selenium in animal nutrition—Utilisation, metabolism, storage and comparison with other selenium sources. J. Appl. Anim. Nutr. 2016, 4, e9. [Google Scholar] [CrossRef]
- Jing, J.; Zeng, H.; Shao, Q.; Tang, J.; Wang, L.; Jia, G.; Liu, G.; Chen, X.; Tian, G.; Cai, J.; et al. Selenomethionine alleviates environmental heat stress induced hepatic lipid accumulation and glycogen infiltration of broilers via maintaining mitochondrial and endoplasmic reticulum homeostasis. Redox Biol. 2023, 67, 102912. [Google Scholar] [CrossRef]
- Thuluvath, P.J.; Triger, D.R. Selenium in chronic liver disease. J. Hepatol. 1992, 14, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xu, J.; Zhang, X.; Wang, J.; Xie, H.; Sun, Y.; Zhang, Q.; Chang, Z.; Liu, Y. Protective Effect of SeMet on Liver Injury Induced by Ochratoxin A in Rabbits. Toxins 2022, 14, 628. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Li, H.; Li, Y.; Li, M.; Zhao, H.; Zhang, W.; Zhao, T.; Wang, Y.; Wang, J.; Wang, J. Selenomethionine supplementation mitigates fluoride-induced liver apoptosis and inflammatory reactions by blocking Parkin-mediated mitophagy in mice. Sci. Total Environ. 2024, 951, 175458. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, J.; Li, H.; Liu, W.; Xi, Y.; Liu, X. A Comprehensive Comparison of Different Selenium Supplements: Mitigation of Heat Stress and Exercise Fatigue-Induced Liver Injury. Front. Nutr. 2022, 9, 917349. [Google Scholar] [CrossRef]
- Zhang, J.; Zhou, H.; Li, H.; Ying, Z.; Liu, X. Research progress on separation of selenoproteins/Se-enriched peptides and their physiological activities. Food Funct. 2021, 12, 1390–1401. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, Q.; Li, M.; Xu, J.; Wang, J.; Li, M.; Wei, L.; Lv, Q.; Chen, X.; Wang, Y.; et al. SeMet attenuates AFB1-induced intestinal injury in rabbits by activating the Nrf2 pathway. Ecotoxicol. Environ. Saf. 2022, 239, 113640. [Google Scholar] [CrossRef]
- Gao, S.; Wang, K.; Xiong, K.; Xiao, S.; Wu, C.; Zhou, M.; Li, L.; Yuan, G.; Jiang, L.; Xiong, Q.; et al. Unraveling the Nrf2-ARE Signaling Pathway in the DF-1 Chicken Fibroblast Cell Line: Insights into T-2 Toxin-Induced Oxidative Stress Regulation. Toxins 2023, 15, 627. [Google Scholar] [CrossRef]
- Wu, Q.H.; Dohnal, V.; Huang, L.L.; Kuca, K.; Yuan, Z.H. Metabolic pathways of trichothecenes. Drug Metab. Rev. 2010, 42, 250–267. [Google Scholar] [CrossRef]
- Königs, M.; Mulac, D.; Schwerdt, G.; Gekle, M.; Humpf, H.U. Metabolism and cytotoxic effects of T-2 toxin and its metabolites on human cells in primary culture. Toxicology 2009, 258, 106–115. [Google Scholar] [CrossRef]
- Li, J.; Bai, Y.; Ma, K.; Ren, Z.; Li, J.; Zhang, J.; Shan, A. Dihydroartemisinin alleviates deoxynivalenol induced liver apoptosis and inflammation in piglets. Ecotoxicol. Environ. Saf. 2022, 241, 113811. [Google Scholar] [CrossRef]
- Yam, C.; Zhao, M.; Hayashi, K.; Ma, H.; Kishimoto, H.; McElroy, M.; Bouvet, M.; Hoffman, R.M. Monotherapy with a Tumor-Targeting Mutant of S. typhimurium Inhibits Liver Metastasis in a Mouse Model of Pancreatic Cancer. J. Surg. Res. 2010, 164, 248–255. [Google Scholar] [CrossRef]
- Wang, L.; Cheng, D.; Wang, H.; Di, L.; Zhou, X.; Xu, T.; Yang, X.; Liu, Y. The hepatoprotective and antifibrotic effects of Saururus chinensis against carbon tetrachloride induced hepatic fibrosis in rats. J. Ethnopharmacol. 2009, 126, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Ramaiah, S.K. A toxicologist guide to the diagnostic interpretation of hepatic biochemical parameters. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2007, 45, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Green, R.M.; Flamm, S. AGA technical review on the evaluation of liver chemistry tests. Gastroenterology 2002, 123, 1367–1384. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Oxidative stress: Oxidants and antioxidants. Exp. Physiol. 1997, 82, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Frijhoff, J.; Winyard, P.G.; Zarkovic, N.; Davies, S.S.; Stocker, R.; Cheng, D.; Knight, A.R.; Taylor, E.L.; Oettrich, J.; Ruskovska, T.; et al. Clinical Relevance of Biomarkers of Oxidative Stress. Antioxid. Redox Signal. 2015, 23, 1144–1170. [Google Scholar] [CrossRef]
- Fang, H.; Wu, Y.; Guo, J.; Rong, J.; Ma, L.; Zhao, Z.; Zuo, D.; Peng, S. T-2 toxin induces apoptosis in differentiated murine embryonic stem cells through reactive oxygen species-mediated mitochondrial pathway. Apoptosis 2012, 17, 895–907. [Google Scholar] [CrossRef]
- Chen, F.; Wang, Y.; Chen, Y.; Fan, J.; Zhang, C.; He, X.; Yang, X. JNK molecule is a toxic target for IPEC-J2 cell barrier damage induced by T-2 toxin. Ecotoxicol. Environ. Saf. 2023, 263, 115247. [Google Scholar] [CrossRef]
- Lauridsen, C. From oxidative stress to inflammation: Redox balance and immune system. Poult. Sci. 2019, 98, 4240–4246. [Google Scholar] [CrossRef]
- Dikalov, S.I.; Dikalova, A.E. Crosstalk Between Mitochondrial Hyperacetylation and Oxidative Stress in Vascular Dysfunction and Hypertension. Antioxid. Redox Signal. 2019, 31, 710–721. [Google Scholar] [CrossRef]
- Peoples, J.N.; Saraf, A.; Ghazal, N.; Pham, T.T.; Kwong, J.Q. Mitochondrial dysfunction and oxidative stress in heart disease. Exp. Mol. Med. 2019, 51, 1–13. [Google Scholar] [CrossRef]
- Chan, S.H.H.; Chan, J.Y.H. Mitochondria and Reactive Oxygen Species Contribute to Neurogenic Hypertension. Physiology 2017, 32, 308–321. [Google Scholar] [CrossRef]
- Scarpulla, R.C. Metabolic control of mitochondrial biogenesis through the PGC-1 family regulatory network. Biochim. Biophys. Acta 2011, 1813, 1269–1278. [Google Scholar] [CrossRef] [PubMed]
- Scarpulla, R.C.; Vega, R.B.; Kelly, D.P. Transcriptional integration of mitochondrial biogenesis. Trends Endocrinol. Metab. 2012, 23, 459–466. [Google Scholar] [CrossRef]
- Ma, S.; Zhao, Y.; Sun, J.; Mu, P.; Deng, Y. miR449a/SIRT1/PGC-1α Is Necessary for Mitochondrial Biogenesis Induced by T-2 Toxin. Front. Pharmacol. 2017, 8, 954. [Google Scholar] [CrossRef] [PubMed]
- Wredenberg, A.; Wibom, R.; Wilhelmsson, H.; Graff, C.; Wiener, H.H.; Burden, S.J.; Oldfors, A.; Westerblad, H.; Larsson, N.G. Increased mitochondrial mass in mitochondrial myopathy mice. Proc. Natl. Acad. Sci. USA 2002, 99, 15066–15071. [Google Scholar] [CrossRef]
- Vanasco, V.; Saez, T.; Magnani, N.D.; Pereyra, L.; Marchini, T.; Corach, A.; Vaccaro, M.I.; Corach, D.; Evelson, P.; Alvarez, S. Cardiac mitochondrial biogenesis in endotoxemia is not accompanied by mitochondrial function recovery. Free Radic. Biol. Med. 2014, 77, 1–9. [Google Scholar] [CrossRef]
- Sun, X.H.; Lv, M.W.; Zhao, Y.X.; Zhang, H.; Ullah Saleem, M.A.; Zhao, Y.; Li, J.L. Nano-Selenium Antagonized Cadmium-Induced Liver Fibrosis in Chicken. J. Agric. Food Chem. 2023, 71, 846–856. [Google Scholar] [CrossRef]
- Jiang, Y.; Qian, Y.; Hong, H.; Gao, X.; Liu, W.; Jin, Q.; Chen, M.; Jin, Z.; Liu, Q.; Wei, Z. Morin protects chicks with T-2 toxin poisoning by decreasing heterophil extracellular traps, oxidative stress and inflammatory response. Br. Poult. Sci. 2023, 64, 614–624. [Google Scholar] [CrossRef]
- Dong, B.; Jiang, Y.; Shi, B.; Zhang, Z.; Zhang, Z. Selenomethionine alleviates decabromodiphenyl ether-induced oxidative stress and ferroptosis via the NRF2/GPX4 pathway in the chicken brain. J. Hazard. Mater. 2024, 465, 133307. [Google Scholar] [CrossRef]
- Bellinger, F.P.; Raman, A.V.; Reeves, M.A.; Berry, M.J. Regulation and function of selenoproteins in human disease. Biochem. J. 2009, 422, 11–22. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zhou, M.; Gao, S.; Li, P.; Zheng, X.; Tu, D.; Yang, L. Selenomethionine Counteracts T-2 Toxin-Induced Liver Injury by Mitigating Oxidative Stress Damage Through the Enhancement of Antioxidant Enzymes. Antioxidants 2025, 14, 866. https://doi.org/10.3390/antiox14070866
Wang Y, Zhou M, Gao S, Li P, Zheng X, Tu D, Yang L. Selenomethionine Counteracts T-2 Toxin-Induced Liver Injury by Mitigating Oxidative Stress Damage Through the Enhancement of Antioxidant Enzymes. Antioxidants. 2025; 14(7):866. https://doi.org/10.3390/antiox14070866
Chicago/Turabian StyleWang, Yan, Mingxia Zhou, Suisui Gao, Pishun Li, Xiaofeng Zheng, Di Tu, and Lingchen Yang. 2025. "Selenomethionine Counteracts T-2 Toxin-Induced Liver Injury by Mitigating Oxidative Stress Damage Through the Enhancement of Antioxidant Enzymes" Antioxidants 14, no. 7: 866. https://doi.org/10.3390/antiox14070866
APA StyleWang, Y., Zhou, M., Gao, S., Li, P., Zheng, X., Tu, D., & Yang, L. (2025). Selenomethionine Counteracts T-2 Toxin-Induced Liver Injury by Mitigating Oxidative Stress Damage Through the Enhancement of Antioxidant Enzymes. Antioxidants, 14(7), 866. https://doi.org/10.3390/antiox14070866