
Skin Photoaging and the Biological Mechanism of the Protective Effects of Hesperidin and Derived Molecules



Abstract
1. Introduction
- The principal objective of cosmetics is to enhance the appearance of the skin; they are incapable of modifying the underlying structure or functionality of the skin. Rather, they provide a transient superficial enhancement, as exemplified by moisturizers;
- The term “cosmeceutical” has been the subject of considerable debate in recent years. Nevertheless, it can be defined as a cross-disciplinary entity, situated at the nexus of the cosmetic and pharmaceutical industries, with the dual objective of combining cosmetic and, to a certain extent, therapeutic benefits for the skin. This results in a range of benefits for the skin, including a reduction in the appearance of wrinkles;
- Nutricosmetics and nutraceuticals, which are products designed for oral ingestion, are employed for the purpose of promoting dermal wellbeing. These products contain molecules that are believed to have a beneficial effect on dermal health when ingested.
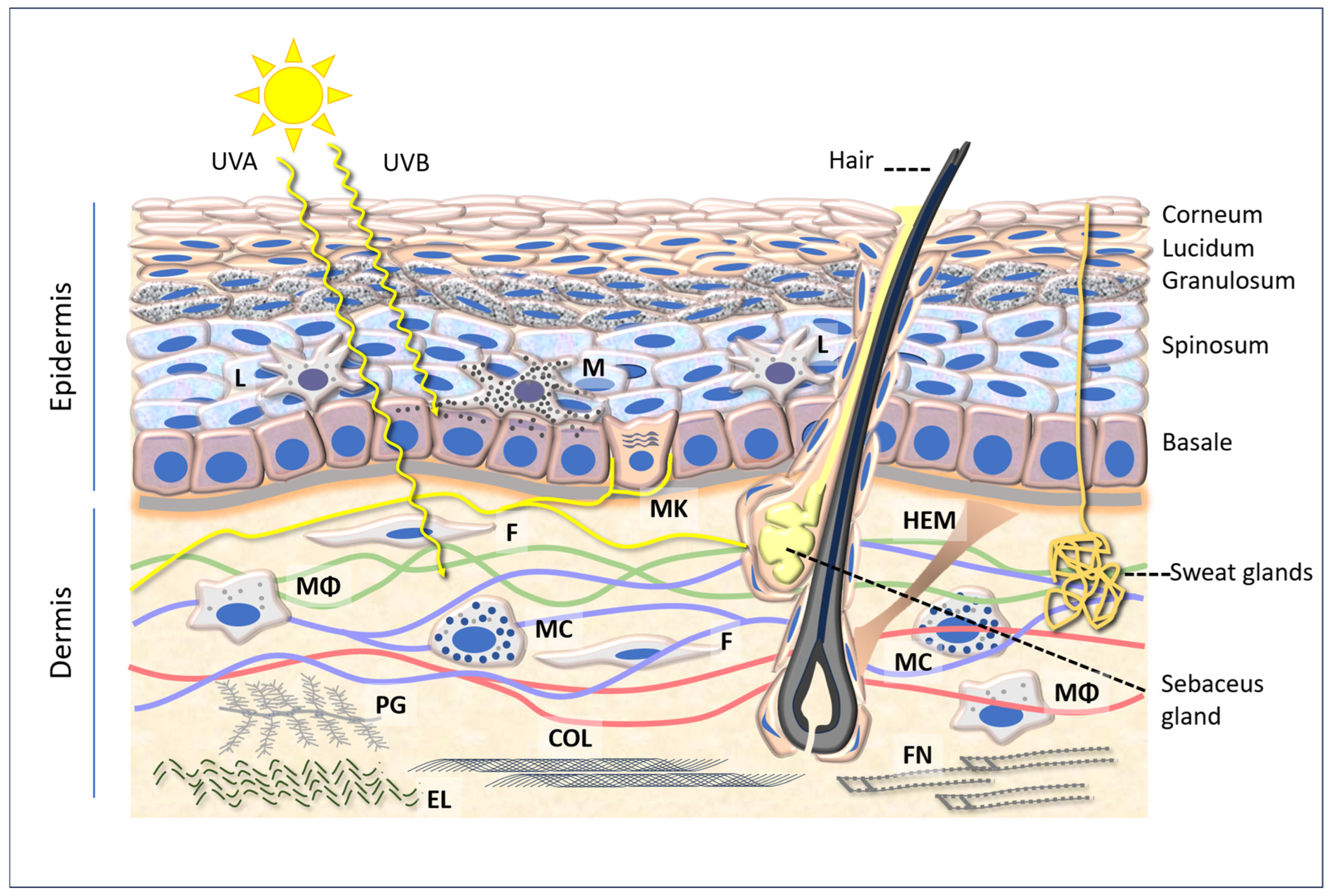
2. Skin Structure Basics
2.1. Epidermis
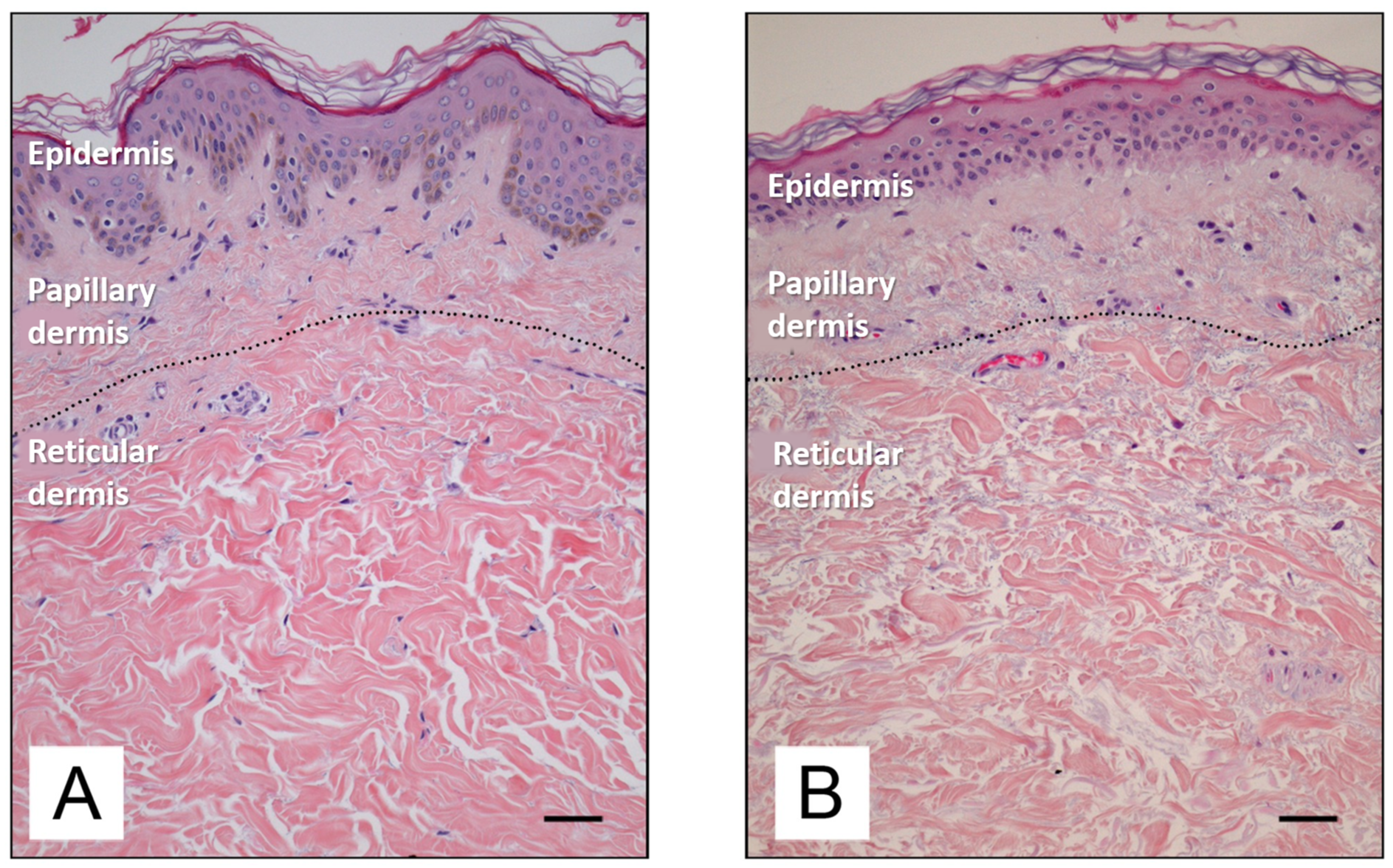
2.2. Dermis
3. Aging of the Skin
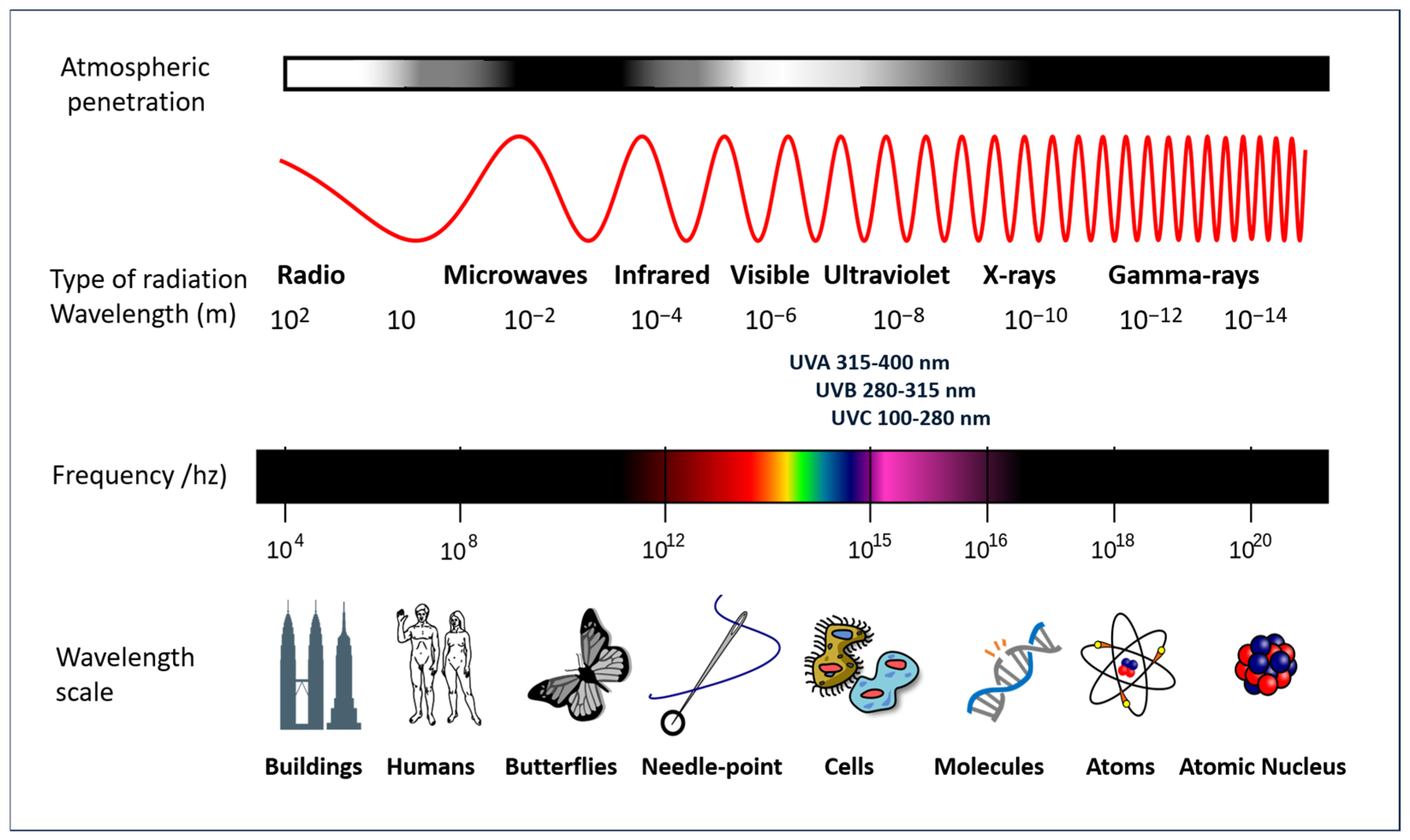
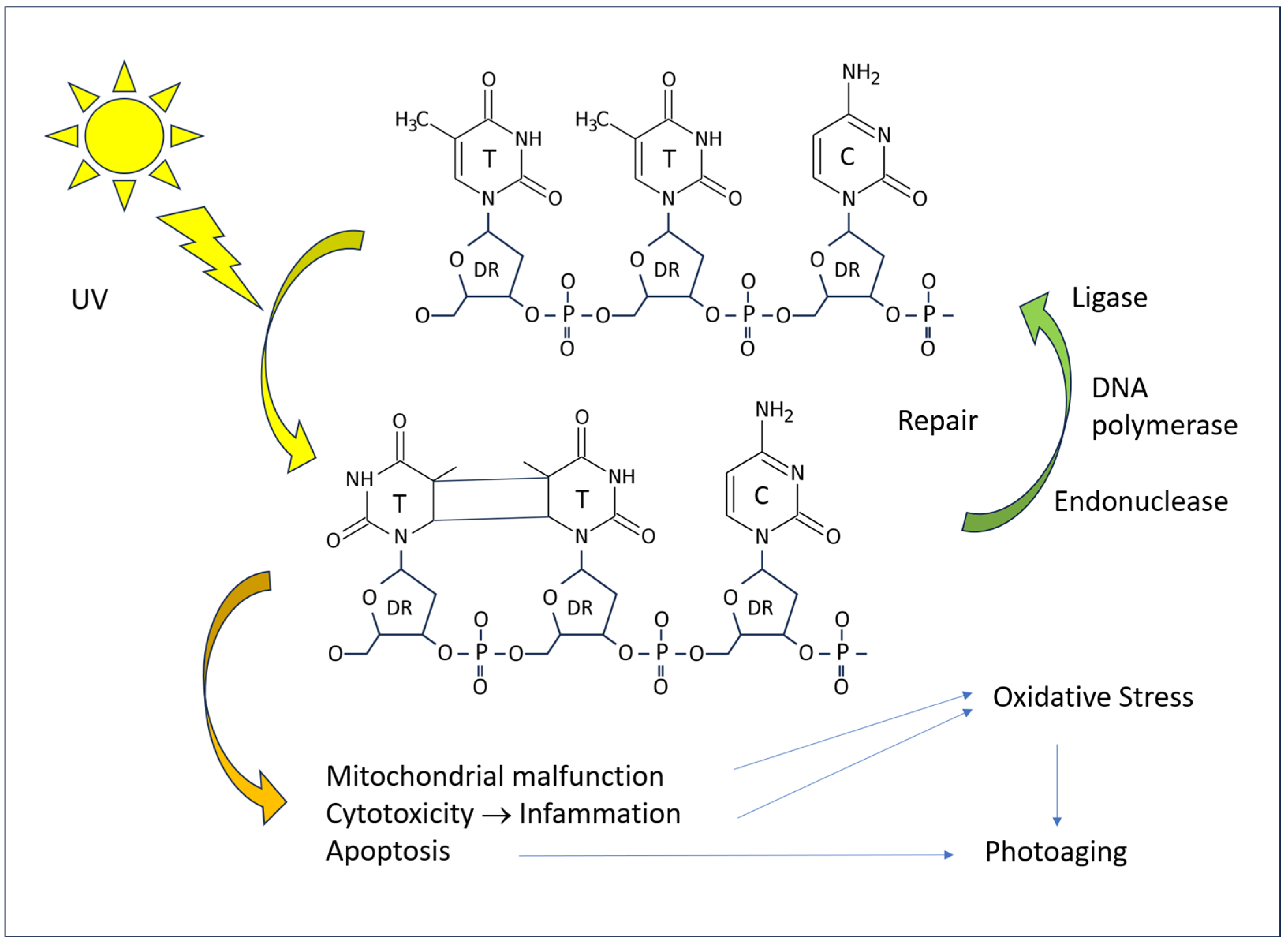
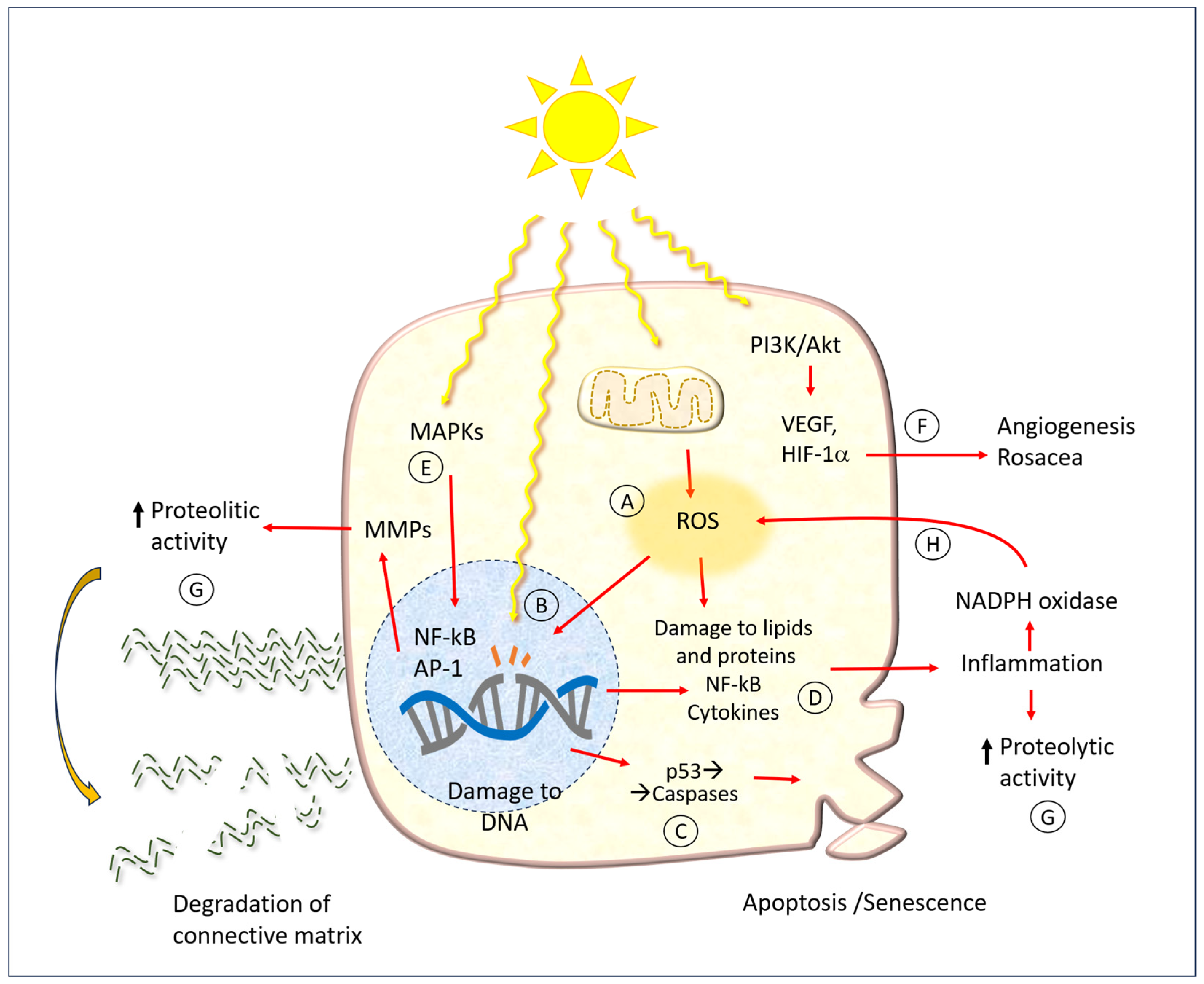
4. Solar Radiation Pathology
4.1. Molecular Mechanisms
4.2. Cellular Mechanisms
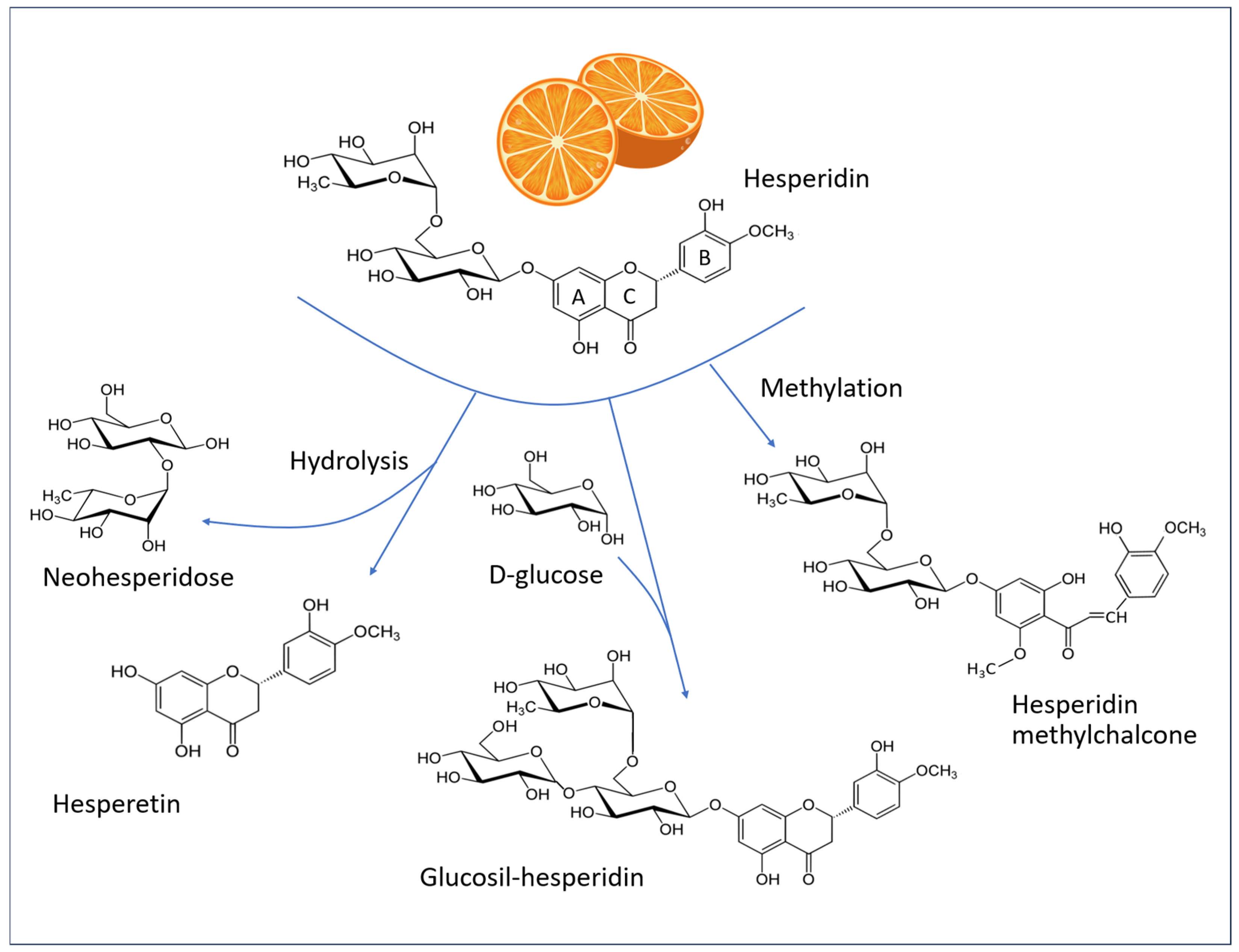
5. Hesperidin and Related Molecules
5.1. Chemistry of Hesperidin
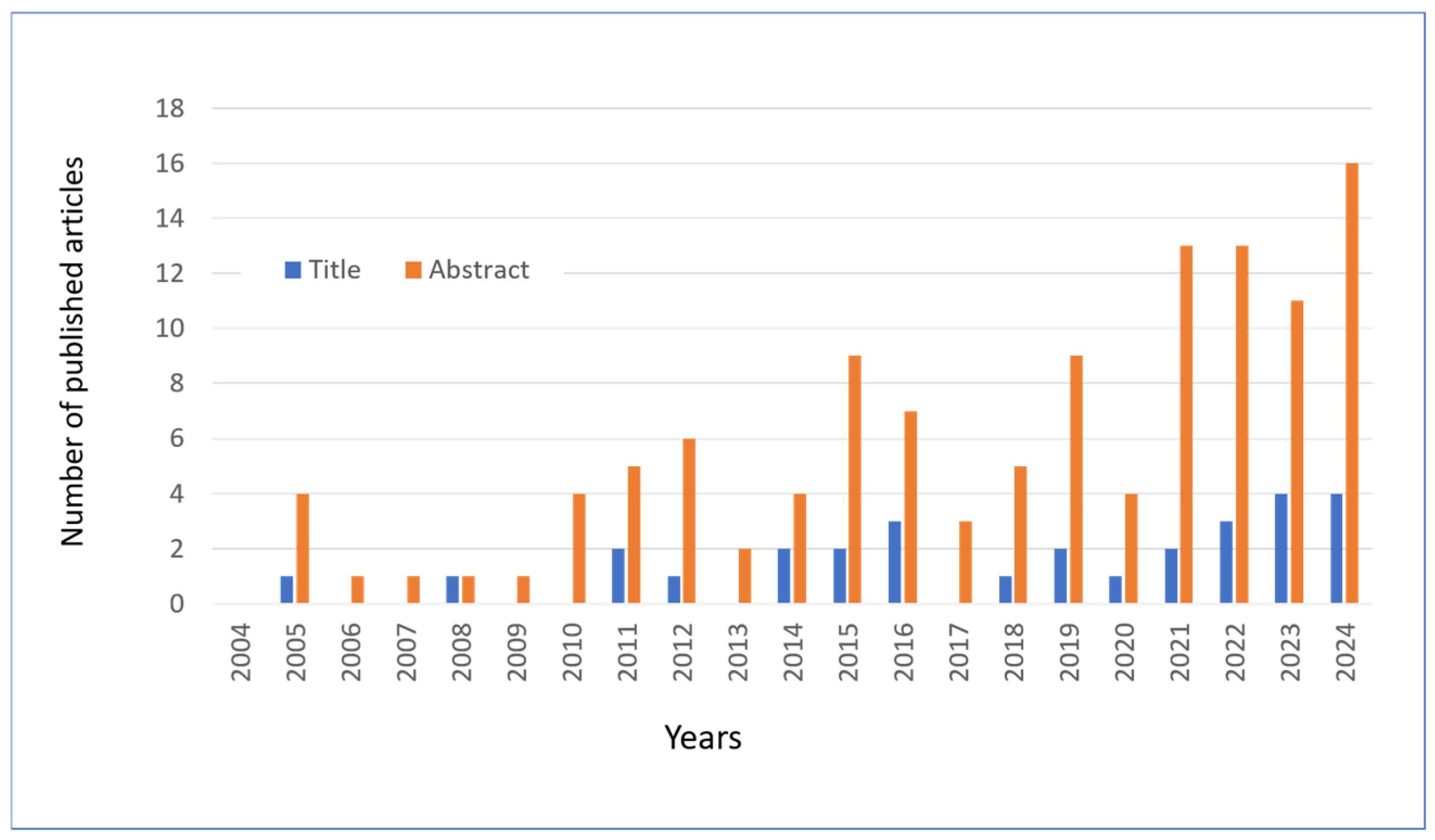
5.2. Bibliographic Research
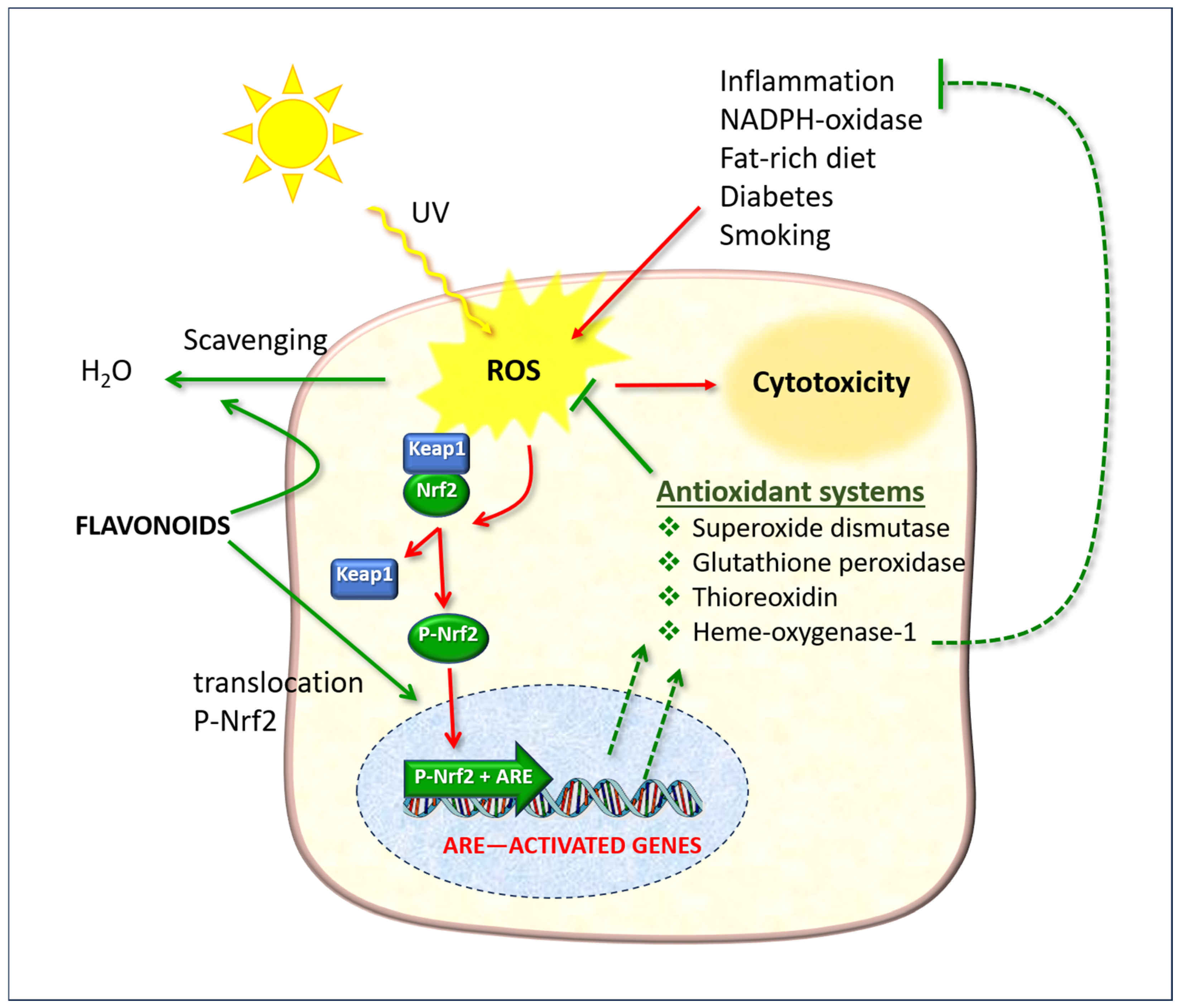
6. Experimental Models of Skin Irradiation
6.1. The Role of the Nrf2/ARE System
6.2. Murine Models
6.3. NADPH Oxidases
7. Other Anti-Aging and Skin Health Actions
7.1. Enzyme Studies
7.2. In Vitro Models
7.3. In Vivo Studies
7.4. Hesperidin and Melanogenesis
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ABCA12 | ATP Binding Cassette Subfamily A Member 12 |
| AP-1 | Activator Protein 1 |
| ARE | Antioxidant Response Element |
| CISD2 | CDGSH Iron Sulfur Domain-containing Protein 2 |
| CPDs | Cyclobutane Pyrimidine Dimers |
| CTLA4 | Cytotoxic T-Lymphocyte Antigen 4 |
| ECM | Extracellular Matrix |
| H2O2 | Hydrogen Peroxide |
| HEK001 | Elderly keratinocytes |
| HIF-1α | Hypoxia-Inducible Factor 1-alpha |
| HMC | Hesperidin Methylchalcone |
| HO-1 | Heme Oxygenase 1 |
| Keap1 | Kelch-like ECH-associated Protein 1 |
| LO2H | Lipid Hydroperoxide |
| LPS | lipopolysaccharide |
| Maf | Musculoaponeurotic Fibrosarcoma protein (small) |
| MAPK | Mitogen-Activated Protein Kinase |
| MCs | Merkel Cells |
| MCC | Merkel Cell Carcinoma |
| MDA | Malondialdehyde |
| MMPs | Matrix Metalloproteinases |
| NF-κB | Nuclear Factor kappa-light-chain-enhancer of activated B cells |
| NO | Nitric Oxide |
| NO3− | Peroxynitrite |
| NOX | NADPH Oxidase |
| Nrf2 | Nuclear factor erythroid 2-related factor 2 |
| OH | Hydroxyl Radical |
| O2− | Superoxide anion |
| PI3K | Phosphoinositide 3-Kinase |
| PIP2 | Phosphatidylinositol 4,5-bisphosphate |
| PIP3 | Phosphatidylinositol 3,4,5-trisphosphate |
| ROS | Reactive Oxygen Species |
| SOD | Superoxide Dismutase |
| T-AOC | Total Antioxidant Capacity |
| Treg | Regulatory T Cell (T-cell subtype) |
| UV | Ultraviolet |
| VEGF | Vascular Endothelial Growth Factor |
References
- Rodrigues, C.V.; Pintado, M. Hesperidin from Orange Peel as a Promising Skincare Bioactive: An Overview. Int. J. Mol. Sci. 2024, 25, 1890. [Google Scholar] [CrossRef]
- Afaq, F.; Katiyar, S.K. Polyphenols: Skin photoprotection and inhibition of photocarcinogenesis. Mini Rev. Med. Chem. 2011, 11, 1200–1215. [Google Scholar]
- Petruk, G.; Del Giudice, R.; Rigano, M.M.; Monti, D.M. Antioxidants from Plants Protect against Skin Photoaging. Oxidative Med. Cell. Longev. 2018, 2018, 1454936. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.K. Natural products in cosmetics. Nat. Prod. Bioprospect. 2022, 12, 40. [Google Scholar] [CrossRef]
- Jesus, A.; Ratanji, S.; Cidade, H.; Sousa, E.; Cruz, M.T.; Oliveira, R.; Almeida, I.F. Phenolics as Active Ingredients in Skincare Products: A Myth or Reality? Molecules 2025, 30, 1423. [Google Scholar] [CrossRef] [PubMed]
- Pla-Paga, L.; Pedret, A.; Valls, R.M.; Calderon-Perez, L.; Llaurado, E.; Companys, J.; Martin-Lujan, F.; Moragas, A.; Canela, N.; Puiggros, F.; et al. Effects of Hesperidin Consumption on the Cardiovascular System in Pre- and Stage 1 Hypertensive Subjects: Targeted and Non-Targeted Metabolomic Approaches (CITRUS Study). Mol. Nutr. Food Res. 2021, 65, e2001175. [Google Scholar] [CrossRef]
- Ji, Z.; Deng, W.; Chen, D.; Liu, Z.; Shen, Y.; Dai, J.; Zhou, H.; Zhang, M.; Xu, H.; Dai, B. Recent understanding of the mechanisms of the biological activities of hesperidin and hesperetin and their therapeutic effects on diseases. Heliyon 2024, 10, e26862. [Google Scholar] [CrossRef] [PubMed]
- Sivakumar, P.M.; Prabhakar, P.K.; Cetinel, S.; Neelakandan, R.; Prabhawathi, V. Molecular Insights on the Therapeutic Effect of Selected Flavonoids on Diabetic Neuropathy. Mini Rev. Med. Chem. 2022, 22, 1828–1846. [Google Scholar] [CrossRef]
- Akdad, M.; Ameziane, R.; Khallouki, F.; Bakri, Y.; Eddouks, M. Antidiabetic Phytocompounds Acting as Glucose Transport Stimulators. Endocr. Metab. Immune Disord. Drug Targets 2023, 23, 147–168. [Google Scholar] [CrossRef]
- Joshi, S.; Dhingra, A.K.; Chopra, B.; Guarve, K.; Bhateja, D. Therapeutic Potential and Clinical Evidence of Hesperidin as Neuroprotective Agent. Cent. Nerv. Syst. Agents Med. Chem. 2022, 22, 5–14. [Google Scholar] [CrossRef]
- Bellavite, P. Neuroprotective Potentials of Flavonoids: Experimental Studies and Mechanisms of Action. Antioxidants 2023, 12, 280. [Google Scholar] [CrossRef] [PubMed]
- Montenegro-Landivar, M.F.; Tapia-Quiros, P.; Vecino, X.; Reig, M.; Valderrama, C.; Granados, M.; Cortina, J.L.; Saurina, J. Polyphenols and their potential role to fight viral diseases: An overview. Sci. Total Environ. 2021, 801, 149719. [Google Scholar] [CrossRef] [PubMed]
- Bellavite, P.; Donzelli, A. Hesperidin and SARS-CoV-2: New Light on the Healthy Function of Citrus Fruits. Antioxidants 2020, 9, 742. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Shin, K.O.; Muraoka, S.; Choi, Y.; Park, J.H.; Park, S.H.; Hwang, J.T.; Park, K.; Uchida, Y. The Epidermal Environment’s Influence on the Dermal Environment in Response to External Stress. Skin Pharmacol. Physiol. 2023, 36, 149–159. [Google Scholar] [CrossRef]
- Furukawa, T.; Yokono, C.; Nomura, Y. Immature mandarin orange extract increases the amount of hyaluronic acid in human skin fibroblast and keratinocytes. Cytotechnology 2024, 76, 271–277. [Google Scholar] [CrossRef]
- Stanisic, D.; Liu, L.H.B.; dos Santos, R.V.; Costa, A.F.; Durán, N.; Tasic, L. New Sustainable Process for Hesperidin Isolation and Anti-Ageing Effects of Hesperidin Nanocrystals. Molecules 2020, 25, 4534. [Google Scholar] [CrossRef]
- Amer, R.I.; Ezzat, S.M.; Aborehab, N.M.; Ragab, M.F.; Mohamed, D.; Hashad, A.; Attia, D.; Salama, M.M.; El Bishbishy, M.H. Downregulation of MMP1 expression mediates the anti-aging activity of Citrus sinensis peel extract nanoformulation in UV induced photoaging in mice. Biomed. Pharmacother. 2021, 138, 111537. [Google Scholar] [CrossRef]
- Pozos-Nonato, S.; Dominguez-Delgado, C.L.; Campos-Santander, K.A.; Benavides, A.A.; Pacheco-Ortin, S.M.; Higuera-Piedrahita, R.I.; Resendiz-Gonzalez, G.; Molina-Trinidad, E.M. Novel Nanotechnological Strategies for Skin Anti-aging. Curr. Pharm. Biotechnol. 2023, 24, 1397–1419. [Google Scholar] [CrossRef]
- Sun, X.; Yang, H.; Zhang, H.; Zhang, W.; Liu, C.; Wang, X.; Song, W.; Wang, L.; Zhao, Q. Magnetic gelatin-hesperidin microrobots promote proliferation and migration of dermal fibroblasts. Front. Chem. 2024, 12, 1478338. [Google Scholar] [CrossRef]
- Talens-Visconti, R.; Belarbi, Y.; Diez-Sales, O.; Julian-Ortiz, J.V.; Vila-Buso, O.; Nacher, A. A New Hyaluronic Emulgel of Hesperetin for Topical Application-An In Vitro Evaluation. J. Funct. Biomater. 2024, 15, 89. [Google Scholar] [CrossRef]
- Maranduca, M.A.; Branisteanu, D.; Serban, D.N.; Branisteanu, D.C.; Stoleriu, G.; Manolache, N.; Serban, I.L. Synthesis and physiological implications of melanic pigments. Oncol. Lett. 2019, 17, 4183–4187. [Google Scholar] [CrossRef] [PubMed]
- Hughes, B.K.; Bishop, C.L. Current Understanding of the Role of Senescent Melanocytes in Skin Ageing. Biomedicines 2022, 10, 3111. [Google Scholar] [CrossRef] [PubMed]
- D’Alba, L.; Shawkey, M.D. Melanosomes: Biogenesis, Properties, and Evolution of an Ancient Organelle. Physiol. Rev. 2019, 99, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, N.C.; Grossman, D. Role of melanin in melanocyte dysregulation of reactive oxygen species. BioMed Res. Int. 2013, 2013, 908797. [Google Scholar] [CrossRef]
- Abraham, J.; Mathew, S. Merkel Cells: A Collective Review of Current Concepts. Int. J. Appl. Basic Med. Res. 2019, 9, 9–13. [Google Scholar] [CrossRef]
- Lacour, J.P.; Dubois, D.; Pisani, A.; Ortonne, J.P. Anatomical mapping of Merkel cells in normal human adult epidermis. Br. J. Dermatol. 1991, 125, 535–542. [Google Scholar] [CrossRef]
- Akaike, T.; Nghiem, P. Scientific and clinical developments in Merkel cell carcinoma: A polyomavirus-driven, often-lethal skin cancer. J. Dermatol. Sci. 2022, 105, 2–10. [Google Scholar] [CrossRef]
- Naylor, E.C.; Watson, R.E.; Sherratt, M.J. Molecular aspects of skin ageing. Maturitas 2011, 69, 249–256. [Google Scholar] [CrossRef]
- Gratchev, A.; Guillot, P.; Hakiy, N.; Politz, O.; Orfanos, C.E.; Schledzewski, K.; Goerdt, S. Alternatively activated macrophages differentially express fibronectin and its splice variants and the extracellular matrix protein betaIG-H3. Scand. J. Immunol. 2001, 53, 386–392. [Google Scholar] [CrossRef]
- Zollinger, A.J.; Smith, M.L. Fibronectin, the extracellular glue. Matrix Biol. 2016, 60, 27–37. [Google Scholar] [CrossRef]
- Ittycheri, A.; Lipsky, Z.W.; Hookway, T.A.; German, G.K. Ultraviolet light induces mechanical and structural changes in full thickness human skin. J. Mech. Behav. Biomed. Mater. 2023, 143, 105880. [Google Scholar] [CrossRef] [PubMed]
- Sarbacher, C.A.; Halper, J.T. Connective Tissue and Age-Related Diseases. Subcell. Biochem. 2019, 91, 281–310. [Google Scholar] [CrossRef] [PubMed]
- Gordon, J.R.; Brieva, J.C. Images in clinical medicine. Unilateral dermatoheliosis. N. Engl. J. Med. 2012, 366, e25. [Google Scholar] [CrossRef]
- Man, G.; Mauro, T.M.; Zhai, Y.; Kim, P.L.; Cheung, C.; Hupe, M.; Crumrine, D.; Elias, P.M.; Man, M.Q. Topical hesperidin enhances epidermal function in an aged murine model. J. Investig. Dermatol. 2015, 135, 1184–1187. [Google Scholar] [CrossRef]
- Mine, S.; Fortunel, N.O.; Pageon, H.; Asselineau, D. Aging alters functionally human dermal papillary fibroblasts but not reticular fibroblasts: A new view of skin morphogenesis and aging. PLoS ONE 2008, 3, e4066. [Google Scholar] [CrossRef]
- Wei, M.; He, X.; Liu, N.; Deng, H. Role of reactive oxygen species in ultraviolet-induced photodamage of the skin. Cell Div. 2024, 19, 1. [Google Scholar] [CrossRef] [PubMed]
- Schuch, A.P.; Moreno, N.C.; Schuch, N.J.; Menck, C.F.M.; Garcia, C.C.M. Sunlight damage to cellular DNA: Focus on oxidatively generated lesions. Free Radic. Biol. Med. 2017, 107, 110–124. [Google Scholar] [CrossRef]
- Papaccio, F.; D’Arino, A.; Caputo, S.; Bellei, B. Focus on the Contribution of Oxidative Stress in Skin Aging. Antioxidants 2022, 11, 1121. [Google Scholar] [CrossRef]
- Yeh, C.H.; Shen, Z.Q.; Lin, C.C.; Lu, C.K.; Tsai, T.F. Rejuvenation: Turning Back Time by Enhancing CISD2. Int. J. Mol. Sci. 2022, 23, 14014. [Google Scholar] [CrossRef]
- Shen, Z.Q.; Chang, C.Y.; Yeh, C.H.; Lu, C.K.; Hung, H.C.; Wang, T.W.; Wu, K.S.; Tung, C.Y.; Tsai, T.F. Hesperetin activates CISD2 to attenuate senescence in human keratinocytes from an older person and rejuvenates naturally aged skin in mice. J. Biomed. Sci. 2024, 31, 15. [Google Scholar] [CrossRef]
- Khavkin, J.; Ellis, D.A. Aging skin: Histology, physiology, and pathology. Facial Plast. Surg. Clin. N. Am. 2011, 19, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Webb, A.R.; Alghamdi, R.; Kift, R.; Rhodes, L.E. 100 YEARS OF VITAMIN D: Dose-response for change in 25-hydroxyvitamin D after UV exposure: Outcome of a systematic review. Endocr. Connect. 2021, 10, R248–R266. [Google Scholar] [CrossRef]
- Hayes, C.E.; Nashold, F.E.; Spach, K.M.; Pedersen, L.B. The immunological functions of the vitamin D endocrine system. Cell. Mol. Biol. 2003, 49, 277–300. [Google Scholar]
- Ribeiro, F.M.; Volpato, H.; Lazarin-Bidoia, D.; Desoti, V.C.; de Souza, R.O.; Fonseca, M.J.V.; Ueda-Nakamura, T.; Nakamura, C.V.; Silva, S.O. The extended production of UV-induced reactive oxygen species in L929 fibroblasts is attenuated by posttreatment with Arrabidaea chica through scavenging mechanisms. J. Photochem. Photobiol. B 2018, 178, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Danno, K.; Toda, K.; Horio, T. Ultraviolet-B radiation suppresses mast cell degranulation induced by compound 48/80. J. Investig. Dermatol. 1986, 87, 775–778. [Google Scholar] [CrossRef]
- Siiskonen, H.; Smorodchenko, A.; Krause, K.; Maurer, M. Ultraviolet radiation and skin mast cells: Effects, mechanisms and relevance for skin diseases. Exp. Dermatol. 2018, 27, 3–8. [Google Scholar] [CrossRef]
- Elder, D.E.; Bastian, B.C.; Cree, I.A.; Massi, D.; Scolyer, R.A. The 2018 World Health Organization Classification of Cutaneous, Mucosal, and Uveal Melanoma: Detailed Analysis of 9 Distinct Subtypes Defined by Their Evolutionary Pathway. Arch. Pathol. Lab. Med. 2020, 144, 500–522. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Yang, T.; Yu, D.; Xiong, H.; Zhang, S. Current insights and future perspectives of ultraviolet radiation (UV) exposure: Friends and foes to the skin and beyond the skin. Environ. Int. 2024, 185, 108535. [Google Scholar] [CrossRef]
- Leung, A.K.; Barankin, B.; Lam, J.M.; Leong, K.F.; Hon, K.L. Xeroderma pigmentosum: An updated review. Drugs Context 2022, 11, 2022-2-5. [Google Scholar] [CrossRef]
- Meinke, M.C.; Busch, L.; Lohan, S.B. Wavelength, dose, skin type and skin model related radical formation in skin. Biophys. Rev. 2021, 13, 1091–1100. [Google Scholar] [CrossRef]
- de Jager, T.L.; Cockrell, A.E.; Du Plessis, S.S. Ultraviolet Light Induced Generation of Reactive Oxygen Species. Adv. Exp. Med. Biol. 2017, 996, 15–23. [Google Scholar] [CrossRef]
- Morgado-Carrasco, D.; Granger, C.; Trullas, C.; Piquero-Casals, J. Impact of ultraviolet radiation and exposome on rosacea: Key role of photoprotection in optimizing treatment. J. Cosmet. Dermatol. 2021, 20, 3415–3421. [Google Scholar] [CrossRef] [PubMed]
- Nagula, R.L.; Wairkar, S. Recent advances in topical delivery of flavonoids: A review. J. Control. Release 2019, 296, 190–201. [Google Scholar] [CrossRef]
- Fernandes, A.; Rodrigues, P.M.; Pintado, M.; Tavaria, F.K. A systematic review of natural products for skin applications: Targeting inflammation, wound healing, and photo-aging. Phytomedicine 2023, 115, 154824. [Google Scholar] [CrossRef] [PubMed]
- Khurana, S.; Venkataraman, K.; Hollingsworth, A.; Piche, M.; Tai, T.C. Polyphenols: Benefits to the cardiovascular system in health and in aging. Nutrients 2013, 5, 3779–3827. [Google Scholar] [CrossRef]
- Zanini, S.; Renzi, S.; Limongi, A.R.; Bellavite, P.; Giovinazzo, F.; Bermano, G. A review of lifestyle and environment risk factors for pancreatic cancer. Eur. J. Cancer 2021, 145, 53–70. [Google Scholar] [CrossRef] [PubMed]
- Zanini, S.; Marzotto, M.; Giovinazzo, F.; Bassi, C.; Bellavite, P. Effects of dietary components on cancer of the digestive system. Crit. Rev. Food Sci. Nutr. 2015, 55, 1870–1885. [Google Scholar] [CrossRef]
- Martiniakova, M.; Babikova, M.; Mondockova, V.; Blahova, J.; Kovacova, V.; Omelka, R. The Role of Macronutrients, Micronutrients and Flavonoid Polyphenols in the Prevention and Treatment of Osteoporosis. Nutrients 2022, 14, 523. [Google Scholar] [CrossRef]
- Porquet, D.; Casadesus, G.; Bayod, S.; Vicente, A.; Canudas, A.M.; Vilaplana, J.; Pelegri, C.; Sanfeliu, C.; Camins, A.; Pallas, M.; et al. Dietary resveratrol prevents Alzheimer’s markers and increases life span in SAMP8. Age 2013, 35, 1851–1865. [Google Scholar] [CrossRef]
- Ozsoy Gokbilen, S.; Becer, E.; Vatansever, H.S. Senescence-mediated anticancer effects of quercetin. Nutr. Res. 2022, 104, 82–90. [Google Scholar] [CrossRef]
- Man, M.Q.; Yang, B.; Elias, P.M. Benefits of Hesperidin for Cutaneous Functions. Evid. Based Complement. Altern. Med. 2019, 2019, 2676307. [Google Scholar] [CrossRef] [PubMed]
- Novotna, R.; Skarupova, D.; Hanyk, J.; Ulrichova, J.; Kren, V.; Bojarova, P.; Brodsky, K.; Vostalova, J.; Frankova, J. Hesperidin, Hesperetin, Rutinose, and Rhamnose Act as Skin Anti-Aging Agents. Molecules 2023, 28, 1728. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.A.; Kasum, C.M. Dietary flavonoids: Bioavailability, metabolic effects, and safety. Annu. Rev. Nutr. 2002, 22, 19–34. [Google Scholar] [CrossRef]
- Sheen, Y.S.; Huang, H.Y.; Liao, Y.H. The efficacy and safety of an antiaging topical serum containing hesperetin and sodium cyclic lysophosphatidic acid: A single-center clinical trial. J. Cosmet. Dermatol. 2021, 20, 3960–3967. [Google Scholar] [CrossRef]
- Gattuso, G.; Barreca, D.; Gargiulli, C.; Leuzzi, U.; Caristi, C. Flavonoid composition of Citrus juices. Molecules 2007, 12, 1641–1673. [Google Scholar] [CrossRef]
- Meneguzzo, F.; Ciriminna, R.; Zabini, F.; Pagliaro, M. Review of Evidence Available on Hesperidin-Rich Products as Potential Tools against COVID-19 and Hydrodynamic Cavitation-Based Extraction as a Method of Increasing Their Production. Processes 2020, 8, 549. [Google Scholar] [CrossRef]
- Stevens, Y.; Rymenant, E.V.; Grootaert, C.; Camp, J.V.; Possemiers, S.; Masclee, A.; Jonkers, D. The Intestinal Fate of Citrus Flavanones and Their Effects on Gastrointestinal Health. Nutrients 2019, 11, 1464. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Shi, Z.; Lei, H.; Chen, G.; Yuan, P.; Cao, Z.; Chen, C.; Zhu, X.; Liu, C.; Dong, M.; et al. Short-Term Intake of Hesperetin-7-O-Glucoside Affects Fecal Microbiota and Host Metabolic Homeostasis in Mice. J. Agric. Food Chem. 2021, 69, 1478–1486. [Google Scholar] [CrossRef]
- Garg, A.; Garg, S.; Zaneveld, L.J.; Singla, A.K. Chemistry and pharmacology of the Citrus bioflavonoid hesperidin. Phytother. Res. 2001, 15, 655–669. [Google Scholar] [CrossRef]
- Hering, A.; Ochocka, J.R.; Baranska, H.; Cal, K.; Stefanowicz-Hajduk, J. Mangiferin and Hesperidin Transdermal Distribution and Permeability through the Skin from Solutions and Honeybush Extracts (Cyclopia sp.)-A Comparison Ex Vivo Study. Molecules 2021, 26, 6547. [Google Scholar] [CrossRef] [PubMed]
- Wdowiak, K.; Walkowiak, J.; Pietrzak, R.; Bazan-Wozniak, A.; Cielecka-Piontek, J. Bioavailability of Hesperidin and Its Aglycone Hesperetin-Compounds Found in Citrus Fruits as a Parameter Conditioning the Pro-Health Potential (Neuroprotective and Antidiabetic Activity)—Mini-Review. Nutrients 2022, 14, 2647. [Google Scholar] [CrossRef]
- Nielsen, I.L.; Chee, W.S.; Poulsen, L.; Offord-Cavin, E.; Rasmussen, S.E.; Frederiksen, H.; Enslen, M.; Barron, D.; Horcajada, M.N.; Williamson, G. Bioavailability is improved by enzymatic modification of the citrus flavonoid hesperidin in humans: A randomized, double-blind, crossover trial. J. Nutr. 2006, 136, 404–408. [Google Scholar] [CrossRef]
- Gil-Izquierdo, A.; Gil, M.I.; Ferreres, F.; Tomas-Barberan, F.A. In vitro availability of flavonoids and other phenolics in orange juice. J. Agric. Food Chem. 2001, 49, 1035–1041. [Google Scholar] [CrossRef]
- Nakazawa, Y.; Aoki, M.; Ishiwa, S.; Morishita, N.; Endo, S.; Nagai, N.; Yamamoto, N.; Funakoshi-Tago, M.; Tamura, H. Oral intake of alpha-glucosyl-hesperidin ameliorates selenite-induced cataract formation. Mol. Med. Rep. 2020, 21, 1258–1266. [Google Scholar] [CrossRef]
- Li, M.; Lin, X.F.; Lu, J.; Zhou, B.R.; Luo, D. Hesperidin ameliorates UV radiation-induced skin damage by abrogation of oxidative stress and inflammatory in HaCaT cells. J. Photochem. Photobiol. B 2016, 165, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.M.; Im, A.R.; Lee, J.Y.; Kim, T.; Ji, K.Y.; Park, D.H.; Chae, S. Hesperidin Inhibits UVB-Induced VEGF Production and Angiogenesis via the Inhibition of PI3K/Akt Pathway in HR-1 Hairless Mice. Biol. Pharm. Bull. 2021, 44, 1492–1498. [Google Scholar] [CrossRef] [PubMed]
- Proteggente, A.R.; Basu-Modak, S.; Kuhnle, G.; Gordon, M.J.; Youdim, K.; Tyrrell, R.; Rice-Evans, C.A. Hesperetin glucuronide, a photoprotective agent arising from flavonoid metabolism in human skin fibroblasts. Photochem. Photobiol. 2003, 78, 256–261. [Google Scholar] [CrossRef]
- Bae, J.T.; Ko, H.J.; Kim, G.B.; Pyo, H.B.; Lee, G.S. Protective effects of fermented Citrus unshiu peel extract against ultraviolet-A-induced photoageing in human dermal fibrobolasts. Phytother. Res. 2012, 26, 1851–1856. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.H.; Lee, K.F.; Huang, Y.B.; Huang, C.T.; Wu, P.C. In vitro permeation and in vivo whitening effect of topical hesperetin microemulsion delivery system. Int. J. Pharm. 2010, 388, 257–262. [Google Scholar] [CrossRef]
- Jin, S.; Zhou, B.; Luo, D. Hesperidin promotes cyclobutane pyrimidine dimer repair in UVB-exposed mice epidermis. Ir. J. Med. Sci. 2011, 180, 709–714. [Google Scholar] [CrossRef]
- Petrova, A.; Davids, L.M.; Rautenbach, F.; Marnewick, J.L. Photoprotection by honeybush extracts, hesperidin and mangiferin against UVB-induced skin damage in SKH-1 mice. J. Photochem. Photobiol. B 2011, 103, 126–139. [Google Scholar] [CrossRef]
- Martinez, R.M.; Pinho-Ribeiro, F.A.; Steffen, V.S.; Caviglione, C.V.; Vignoli, J.A.; Baracat, M.M.; Georgetti, S.R.; Verri, W.A., Jr.; Casagrande, R. Hesperidin methyl chalcone inhibits oxidative stress and inflammation in a mouse model of ultraviolet B irradiation-induced skin damage. J. Photochem. Photobiol. B 2015, 148, 145–153. [Google Scholar] [CrossRef]
- Martinez, R.M.; Pinho-Ribeiro, F.A.; Steffen, V.S.; Caviglione, C.V.; Pala, D.; Baracat, M.M.; Georgetti, S.R.; Verri, W.A.; Casagrande, R. Topical formulation containing hesperidin methyl chalcone inhibits skin oxidative stress and inflammation induced by ultraviolet B irradiation. Photochem. Photobiol. Sci. 2016, 15, 554–563. [Google Scholar] [CrossRef]
- Lee, H.J.; Im, A.R.; Kim, S.M.; Kang, H.S.; Lee, J.D.; Chae, S. The flavonoid hesperidin exerts anti-photoaging effect by downregulating matrix metalloproteinase (MMP)-9 expression via mitogen activated protein kinase (MAPK)-dependent signaling pathways. BMC Complement. Altern. Med. 2018, 18, 39. [Google Scholar] [CrossRef] [PubMed]
- de Araujo Andrade, T.; Heimfarth, L.; Dos Santos, D.M.; Dos Santos, M.R.V.; de Albuquerque-Junior, R.L.C.; Dos Santos-Neto, A.G.; de Araujo, G.R.S.; Lira, A.A.M.; Matos, S.S.; Frank, L.A.; et al. Hesperetin-Based Hydrogels Protect the Skin against UV Radiation-Induced Damage. AAPS PharmSciTech 2022, 23, 170. [Google Scholar] [CrossRef] [PubMed]
- Madduma Hewage, S.R.; Piao, M.J.; Kang, K.A.; Ryu, Y.S.; Han, X.; Oh, M.C.; Jung, U.; Kim, I.G.; Hyun, J.W. Hesperidin Attenuates Ultraviolet B-Induced Apoptosis by Mitigating Oxidative Stress in Human Keratinocytes. Biomol. Ther. 2016, 24, 312–319. [Google Scholar] [CrossRef]
- Choi, I.Y.; Kim, S.J.; Jeong, H.J.; Park, S.H.; Song, Y.S.; Lee, J.H.; Kang, T.H.; Park, J.H.; Hwang, G.S.; Lee, E.J.; et al. Hesperidin inhibits expression of hypoxia inducible factor-1 alpha and inflammatory cytokine production from mast cells. Mol. Cell. Biochem. 2007, 305, 153–161. [Google Scholar] [CrossRef]
- Takahara, Y.; Miyachi, N.; Nawa, M.; Matsuoka, M. Calmodulin-like skin protein suppresses the increase in senescence-associated beta-galactosidase induced by hydrogen peroxide or ultraviolet irradiation in keratinocytes. Cell Biol. Int. 2019, 43, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Valieva, Y.; Ivanova, E.; Fayzullin, A.; Kurkov, A.; Igrunkova, A. Senescence-Associated beta-Galactosidase Detection in Pathology. Diagnostics 2022, 12, 2309. [Google Scholar] [CrossRef]
- Park, H.K.; Kang, S.W.; Park, M.S. Hesperidin Ameliorates Hepatic Ischemia-Reperfusion Injury in Sprague-Dawley Rats. Transplant. Proc. 2019, 51, 2828–2832. [Google Scholar] [CrossRef]
- Kuwano, T.; Watanabe, M.; Kagawa, D.; Murase, T. Hydrolyzed Methylhesperidin Induces Antioxidant Enzyme Expression via the Nrf2-ARE Pathway in Normal Human Epidermal Keratinocytes. J. Agric. Food Chem. 2015, 63, 7937–7944. [Google Scholar] [CrossRef] [PubMed]
- Chaiprasongsuk, A.; Panich, U. Role of Phytochemicals in Skin Photoprotection via Regulation of Nrf2. Front. Pharmacol. 2022, 13, 823881. [Google Scholar] [CrossRef]
- Jo, S.H.; Kim, M.E.; Cho, J.H.; Lee, Y.; Lee, J.; Park, Y.D.; Lee, J.S. Hesperetin inhibits neuroinflammation on microglia by suppressing inflammatory cytokines and MAPK pathways. Arch. Pharm. Res. 2019, 42, 695–703. [Google Scholar] [CrossRef]
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. An Overview of Nrf2 Signaling Pathway and Its Role in Inflammation. Molecules 2020, 25, 5474. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Mohammed, H.M.; Khadrawy, S.M.; Galaly, S.R. Hesperidin protects against chemically induced hepatocarcinogenesis via modulation of Nrf2/ARE/HO-1, PPARÎ3 and TGF-Î21/Smad3 signaling, and amelioration of oxidative stress and inflammation. Chem. Biol. Interact. 2017, 277, 146–158. [Google Scholar] [CrossRef]
- Tabeshpour, J.; Hosseinzadeh, H.; Hashemzaei, M.; Karimi, G. A review of the hepatoprotective effects of hesperidin, a flavanon glycoside in citrus fruits, against natural and chemical toxicities. Daru 2020, 28, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Schafer, M.; Dutsch, S.; auf dem Keller, U.; Navid, F.; Schwarz, A.; Johnson, D.A.; Johnson, J.A.; Werner, S. Nrf2 establishes a glutathione-mediated gradient of UVB cytoprotection in the epidermis. Genes Dev. 2010, 24, 1045–1058. [Google Scholar] [CrossRef] [PubMed]
- Soares, M.P.; Seldon, M.P.; Gregoire, I.P.; Vassilevskaia, T.; Berberat, P.O.; Yu, J.; Tsui, T.Y.; Bach, F.H. Heme oxygenase-1 modulates the expression of adhesion molecules associated with endothelial cell activation. J. Immunol. 2004, 172, 3553–3563. [Google Scholar] [CrossRef]
- Ganesh Yerra, V.; Negi, G.; Sharma, S.S.; Kumar, A. Potential therapeutic effects of the simultaneous targeting of the Nrf2 and NF-kappaB pathways in diabetic neuropathy. Redox Biol. 2013, 1, 394–397. [Google Scholar] [CrossRef]
- Hernandez-Pigeon, H.; Garidou, L.; Galliano, M.F.; Delga, H.; Aries, M.F.; Duplan, H.; Bessou-Touya, S.; Castex-Rizzi, N. Effects of dextran sulfate, 4-t-butylcyclohexanol, pongamia oil and hesperidin methyl chalcone on inflammatory and vascular responses implicated in rosacea. Clin. Cosmet. Investig. Dermatol. 2018, 11, 421–429. [Google Scholar] [CrossRef]
- Bellavite, P. The superoxide-forming enzymatic system of phagocytes. Free Radic. Biol. Med. 1988, 4, 225–261. [Google Scholar] [CrossRef]
- Vermot, A.; Petit-Hartlein, I.; Smith, S.M.E.; Fieschi, F. NADPH Oxidases (NOX): An Overview from Discovery, Molecular Mechanisms to Physiology and Pathology. Antioxidants 2021, 10, 890. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.P.; Tse, H.M. The role of NADPH oxidases in infectious and inflammatory diseases. Redox Biol. 2021, 48, 102159. [Google Scholar] [CrossRef]
- Bedard, K.; Krause, K.H. The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Ha, S.J.; So, B.R.; Kim, C.K.; Kim, K.M.; Jung, S.K. NADPH Oxidase and Epidermal Growth Factor Receptor Are Promising Targets of Phytochemicals for Ultraviolet-Induced Skin Carcinogenesis. Antioxidants 2021, 10, 1909. [Google Scholar] [CrossRef] [PubMed]
- Raad, H.; Serrano-Sanchez, M.; Harfouche, G.; Mahfouf, W.; Bortolotto, D.; Bergeron, V.; Kasraian, Z.; Dousset, L.; Hosseini, M.; Taieb, A.; et al. NADPH Oxidase-1 Plays a Key Role in Keratinocyte Responses to UV Radiation and UVB-Induced Skin Carcinogenesis. J. Investig. Dermatol. 2017, 137, 1311–1321. [Google Scholar] [CrossRef]
- Kocak, A.; Ural, C.; Harmanci, D.; Oktan, M.A.; Afagh, A.; Sarioglu, S.; Yilmaz, O.; Birlik, M.; Akdogan, G.G.; Cavdar, Z. Protective effects of alpha-lipoic acid on bleomycin-induced skin fibrosis through the repression of NADPH Oxidase 4 and TGF-beta1/Smad3 signaling pathways. Hum. Exp. Toxicol. 2022, 41, 9603271211065975. [Google Scholar] [CrossRef]
- Yousefian, M.; Shakour, N.; Hosseinzadeh, H.; Hayes, A.W.; Hadizadeh, F.; Karimi, G. The natural phenolic compounds as modulators of NADPH oxidases in hypertension. Phytomedicine 2019, 55, 200–213. [Google Scholar] [CrossRef]
- Yamamoto, M.; Jokura, H.; Suzuki, A.; Hase, T.; Shimotoyodome, A. Effects of continuous ingestion of hesperidin and glucosyl hesperidin on vascular gene expression in spontaneously hypertensive rats. J Nutr. Sci Vitaminol. 2013, 59, 470–473. [Google Scholar] [CrossRef]
- Wunpathe, C.; Potue, P.; Maneesai, P.; Bunbupha, S.; Prachaney, P.; Kukongviriyapan, U.; Kukongviriyapan, V.; Pakdeechote, P. Hesperidin Suppresses Renin-Angiotensin System Mediated NOX2 Over-Expression and Sympathoexcitation in 2K-1C Hypertensive Rats. Am. J. Chin. Med. 2018, 46, 751–767. [Google Scholar] [CrossRef]
- Syed, A.A.; Reza, M.I.; Yadav, H.; Gayen, J.R. Hesperidin inhibits NOX4 mediated oxidative stress and inflammation by upregulating SIRT1 in experimental diabetic neuropathy. Exp. Gerontol. 2023, 172, 112064. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Suzuki, A.; Jokura, H.; Yamamoto, N.; Hase, T. Glucosyl hesperidin prevents endothelial dysfunction and oxidative stress in spontaneously hypertensive rats. Nutrition 2008, 24, 470–476. [Google Scholar] [CrossRef]
- Hou, M.; Man, M.; Man, W.; Zhu, W.; Hupe, M.; Park, K.; Crumrine, D.; Elias, P.M.; Man, M.Q. Topical hesperidin improves epidermal permeability barrier function and epidermal differentiation in normal murine skin. Exp. Dermatol. 2012, 21, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Man, G.; Mauro, T.M.; Kim, P.L.; Hupe, M.; Zhai, Y.; Sun, R.; Crumrine, D.; Cheung, C.; Nuno-Gonzalez, A.; Elias, P.M.; et al. Topical hesperidin prevents glucocorticoid-induced abnormalities in epidermal barrier function in murine skin. Exp. Dermatol. 2014, 23, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Kandhare, A.D.; Mukherjee, A.A.; Bodhankar, S.L. Hesperidin, a plant flavonoid accelerated the cutaneous wound healing in streptozotocin-induced diabetic rats: Role of TGF-ss/Smads and Ang-1/Tie-2 signaling pathways. EXCLI J. 2018, 17, 399–419. [Google Scholar] [CrossRef]
- Wang, L.; He, T.; Fu, A.; Mao, Z.; Yi, L.; Tang, S.; Yang, J. Hesperidin enhances angiogenesis via modulating expression of growth and inflammatory factor in diabetic foot ulcer in rats. Eur. J. Inflamm. 2018, 16, 1–13. [Google Scholar] [CrossRef]
- Vabeiryureilai, M.; Lalrinzuali, K.; Jagetia, G.C. NF-kappaB and COX-2 repression with topical application of hesperidin and naringin hydrogels augments repair and regeneration of deep dermal wounds. Burns 2022, 48, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Jagetia, G.C.; Rao, K.V.N.M. Hesperidin treatment abates radiation-induced delay in healing of deep cutaneous excision wound of mice hemi-body exposed to different doses of γ-radiation. Clin. Dermatol. Dermat. 2018, 1, 104. [Google Scholar]
- Yang, P.; Zhong, J.; Zhao, X.; Ali, K.; Wu, L.; Bu, Z. Exploring the Potential of Hesperidin in Preventing Hypertrophic Scars: Insights from a Rabbit Ear Model. Clin. Cosmet. Investig. Dermatol. 2023, 16, 2957–2963. [Google Scholar] [CrossRef]
- Moon, P.D.; Kim, H.M. Antiinflammatory effects of traditional Korean medicine, JinPi-tang and its active ingredient, hesperidin in HaCaT cells. Phytother. Res. 2012, 26, 657–662. [Google Scholar] [CrossRef]
- Kazlowska, K.; Hsu, T.; Hou, C.C.; Yang, W.C.; Tsai, G.J. Anti-inflammatory properties of phenolic compounds and crude extract from Porphyra dentata. J. Ethnopharmacol. 2010, 128, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.L.; Chen, S.C.; Senthil Kumar, K.J.; Yu, K.N.; Lee Chao, P.D.; Tsai, S.Y.; Hou, Y.C.; Hseu, Y.C. Antioxidant and anti-inflammatory potential of hesperetin metabolites obtained from hesperetin-administered rat serum: An ex vivo approach. J. Agric. Food Chem. 2012, 60, 522–532. [Google Scholar] [CrossRef] [PubMed]
- Nagashio, Y.; Matsuura, Y.; Miyamoto, J.; Kometani, T.; Suzuki, T.; Tanabe, S. Hesperidin inhibits development of atopic dermatitis-like skin lesions in NC/Nga mice by suppressing Th17 activity. J. Funct. Foods 2013, 5, 1633–1641. [Google Scholar] [CrossRef]
- Vidhya, R.; Anbumani, V.I.; Dinakara, R.A.; Anuradha, C.V. Identification of novel human neutrophil elastase inhibitors from dietary phytochemicals using in silico and in vitro studies. J. Biomol. Struct. Dyn. 2020, 40, 3451–3461. [Google Scholar] [CrossRef]
- Emim, J.A.; Oliveira, A.B.; Lapa, A.J. Pharmacological evaluation of the anti-inflammatory activity of a citrus bioflavonoid, hesperidin, and the isoflavonoids, duartin and claussequinone, in rats and mice. J. Pharm. Pharmacol. 1994, 46, 118–122. [Google Scholar] [CrossRef]
- Pelzer, L.E.; Guardia, T.; Osvaldo Juarez, A.; Guerreiro, E. Acute and chronic antiinflammatory effects of plant flavonoids. Farmaco 1998, 53, 421–424. [Google Scholar] [CrossRef]
- Lee, N.K.; Choi, S.H.; Park, S.H.; Park, E.K.; Kim, D.H. Antiallergic activity of hesperidin is activated by intestinal microflora. Pharmacology 2004, 71, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Deshetty, U.M.; Tamatam, A.; Bhattacharjee, M.; Perumal, E.; Natarajan, G.; Khanum, F. Ameliorative Effect of Hesperidin Against Motion Sickness by Modulating Histamine and Histamine H1 Receptor Expression. Neurochem. Res. 2020, 45, 371–384. [Google Scholar] [CrossRef]
- Artero, N.A.; Manchope, M.F.; Carvalho, T.T.; Saraiva-Santos, T.; Bertozzi, M.M.; Carneiro, J.A.; Franciosi, A.; Dionisio, A.M.; Zaninelli, T.H.; Fattori, V.; et al. Hesperidin Methyl Chalcone Reduces the Arthritis Caused by TiO2 in Mice: Targeting Inflammation, Oxidative Stress, Cytokine Production, and Nociceptor Sensory Neuron Activation. Molecules 2023, 28, 872. [Google Scholar] [CrossRef]
- Buzdagli, Y.; Eyipinar, C.D.; Kaci, F.N.; Tekin, A. Effects of hesperidin on anti-inflammatory and antioxidant response in healthy people: A meta-analysis and meta-regression. Int. J. Environ. Health Res. 2022, 33, 1390–1405. [Google Scholar] [CrossRef]
- Suto, H.; Matsuda, H.; Mitsuishi, K.; Hira, K.; Uchida, T.; Unno, T.; Ogawa, H.; Ra, C. NC/Nga mice: A mouse model for atopic dermatitis. Int. Arch. Allergy Immunol. 1999, 120 (Suppl. S1), 70–75. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, Y.; Doki, Y.; Sugiyama, Y.; Kobayashi, R.; Nagai, N.; Morisita, N.; Endo, S.; Funakoshi-Tago, M.; Tamura, H. Effect of Alpha-Glucosyl-Hesperidin Consumption on Lens Sclerosis and Presbyopia. Cells 2021, 10, 382. [Google Scholar] [CrossRef]
- Yu, L.; Zhang, Q.; Zhou, L.; Wei, Y.; Li, M.; Wu, X.; Xin, M. Ocular topical application of alpha-glucosyl hesperidin as an active pharmaceutical excipient: In vitro and in vivo experimental evaluation. Drug Deliv. Transl. Res. 2024, 14, 373–385. [Google Scholar] [CrossRef]
- Nakazawa, Y.; Aoki, M.; Doki, Y.; Morishita, N.; Endo, S.; Nagai, N.; Funakoshi-Tago, M.; Tamura, H. Oral consumption of alpha-glucosyl-hesperidin could prevent lens hardening, which causes presbyopia. Biochem. Biophys. Rep. 2021, 25, 100885. [Google Scholar] [CrossRef]
- Rehman, U.; Sheikh, A.; Alsayari, A.; Wahab, S.; Kesharwani, P. Hesperidin-loaded cubogel as a novel therapeutic armamentarium for full-thickness wound healing. Colloids Surf. B Biointerfaces 2024, 234, 113728. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Miyazawa, K.W.; Pinho-Ribeiro, F.A.; Borghi, S.M.; Staurengo-Ferrari, L.; Fattori, V.; Amaral, F.A.; Teixeira, M.M.; Alves-Filho, J.C.; Cunha, T.M.; Cunha, F.Q.; et al. Hesperidin Methylchalcone Suppresses Experimental Gout Arthritis in Mice by Inhibiting NF-kappaB Activation. J. Agric. Food Chem. 2018, 66, 6269–6280. [Google Scholar] [CrossRef]
- Morishita, N.; Ogihara, S.; Endo, S.; Mitsuzumi, H.; Ushio, S. Effects of glucosyl hesperidin on skin blood flow and temperature: A randomized, double-blind, placebo-controlled, crossover study. Shinryo Shinyaku 2020, 57, 129–134. [Google Scholar]
- Kapoor, M.P.; Moriwaki, M.; Abe, A.; Morishima, S.; Ozeki, M.; Sato, N. Hesperetin-7-O-glucoside/beta-cyclodextrin Inclusion Complex Induces Acute Vasodilator Effect to Inhibit the Cold Sensation Response during Localized Cold-Stimulate Stress in Healthy Human Subjects: A Randomized, Double-Blind, Crossover, and Placebo-Controlled Study. Nutrients 2023, 15, 3702. [Google Scholar] [CrossRef]
- Verma, A.; Dedar, R.K.; Kumar, R.; Chander, Y.; Kamboj, H.; Kumar, G.; Verma, R.; Kumari, S.; Sharma, S.; Tripathi, B.N.; et al. Hesperetin blocks poxvirus replication with a low tendency to select for drug-resistant viral variants. J. Med. Virol. 2024, 96, e29555. [Google Scholar] [CrossRef]
- Kim, B.; Lee, J.Y.; Lee, H.Y.; Nam, K.Y.; Park, J.; Lee, S.M.; Kim, J.E.; Lee, J.D.; Hwang, J.S. Hesperidin Suppresses Melanosome Transport by Blocking the Interaction of Rab27A-Melanophilin. Biomol. Ther. 2013, 21, 343–348. [Google Scholar] [CrossRef]
- Huang, Y.C.; Liu, K.C.; Chiou, Y.L. Melanogenesis of murine melanoma cells induced by hesperetin, a Citrus hydrolysate-derived flavonoid. Food Chem. Toxicol. 2012, 50, 653–659. [Google Scholar] [CrossRef] [PubMed]
- Usach, I.; Talens-Visconti, R.; Magraner-Pardo, L.; Peris, J.E. Hesperetin induces melanin production in adult human epidermal melanocytes. Food Chem. Toxicol. 2015, 80, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Lee, W.J.; Chang, S.E.; Lee, G.Y. Hesperidin, A Popular Antioxidant Inhibits Melanogenesis via Erk1/2 Mediated MITF Degradation. Int. J. Mol. Sci. 2015, 16, 18384–18395. [Google Scholar] [CrossRef]
- Si, Y.X.; Wang, Z.J.; Park, D.; Chung, H.Y.; Wang, S.F.; Yan, L.; Yang, J.M.; Qian, G.Y.; Yin, S.J.; Park, Y.D. Effect of hesperetin on tyrosinase: Inhibition kinetics integrated computational simulation study. Int. J. Biol. Macromol. 2012, 50, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Tian, H.; Guo, F.; Wu, Y. Inhibition characteristics and mechanism of tyrosinase using five citrus flavonoids: A spectroscopic and molecular dynamics simulation study. J. Food Biochem. 2022, 46, e14484. [Google Scholar] [CrossRef]
- Shivasaraun, U.V.; Sureshkumar, R.; Karthika, C.; Puttappa, N. Flavonoids as adjuvant in psoralen based photochemotherapy in the management of vitiligo/leucoderma. Med. Hypotheses 2018, 121, 26–30. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Route | Treatment | Effects | Mechanisms | Ref. |
|---|---|---|---|---|---|
| Human keratinocytes (HaCaT) exposed to UVA (10 J/cm2) | In vitro | 220 µg/mL of pure hesperidin, purity ≥ 98% (commercial product, Procter & Gamble, Cincinnati, OH, USA) applied to the cell culture for 24 h | ↑ Cellular vitality ↓ malondialdehyde production | ↓ mRNA levels of TNF-α, IL-1β, and IL-6 ↑ SOD activity | [75] |
| Keratinocytes from an elderly person (HEK001 cells) exposed to UVB (20 J/m2) | In vitro | Pre-incubation for 24 h with 1–10 µM pure hesperidin, purity ≥ 98% (Sigma-Aldrich, St. Louis, MO, USA) | Photoprotection | ↓ VEGF expression ↓ Expression of MMP-9, MMP-13 ↓ PI3K/Akt signaling | [76] |
| Keratinocytes from an elderly person (HEK001 cells), exposed to UVB (20 J/m2) | In vitro | 10 µM pure hesperetin, purity ≥ 98% (Sigma-Aldrich) for 24/48 h | ↑ Mitochondrial function ↓ Oxidative stress | ↑ CISD2 ↑ FOXM1 ↑ FOXO3a ↓ MMP-1 | [40] |
| Human skin fibroblasts (FEK4), exposed to UVA (500 kJ/m2) | In vitro | 3 or 30 µM hesperetin glucuronide (synthesized and verified by HPLC-DAD, LC-MS/MS) for 18 h | ↓ Cellular necrosis | Not determined | [77] |
| Human dermal fibroblasts irradiated with UVA (10 J/cm2) | In vitro | Fermented Citrus unshiu peel extract (commercial product; rich in hesperetin and naringenin) for 72 h | Photoprotection ↑ Biosynthesis of collagen | ↓ Expression of MMPs ↓ Expression of β-galactosidase | [78] |
| Guinea pig, exposed to UVB for 2 weeks, dose not specified | Topical application | Pure hesperetin 1% in microemulsion (commercial product, supplier not specified) | ↓ Skin irritation ↓ Pigmentation ↓ Trans epidermal water loss | Not determined | [79] |
| Balb/C mouse, exposed once to UVB (180 mJ/cm2) | Topical application | 1 mg hesperidin in 100 µL acetone/cm2 applied once daily for 3 days, purity ≥ 98% (from National Institute for the Control of Pharmaceutical and Biological Products, Beijing, China) | Photoprotection | ↑ p53 expression ↓ Cyclobutane-pyrimidine epidermal dimers | [80] |
| SKH-1 mouse, exposed daily to UVB (180 mJ/cm2) | Topical application | 30 min pretreatment with hesperidin 3 mg/mL, purity ≥ 98% (commercial product; supplier not specified) for 10 days | ↓ Erythema and skin edema ↓ Epidermal hyperplasia ↓ Lipid peroxidation ↓ Inflammation | ↑ Catalase and superoxide dismutase activity ↓ DNA damage | [81] |
| Hairless mouse exposed to UVB (4.14 mJ/cm2) | Intraperitoneal injection | 300 mg/kg HMC, purity ≥ 98% (Sigma-Aldrich), 1 h before and 7 h after each exposure | ↓ Skin edema and neutrophil recruitment ↓ Expression of inflammatory cytokines ↓ Lipid peroxidation | ↑ Glutathione levels and catalase activity ↓ Myeloperoxidase ↓ MMP9 ↓ NOX | [82] |
| SKH-1 hairless mice exposed to UVB (4.14 mJ/cm2) | Topical application | 1% HMC, purity ≥ 98% (Sigma-Aldrich) in lanette cream base, daily for 5 days | ↓ Skin edema ↓ Lipid peroxidation ↑ Antioxidant capacity ↓ O2− production | ↓ IL-6 and TNF-a ↓ NOX ↑ Nrf2 expression ↑ Glutathione peroxidase, glutathione reductase, and heme oxygenase-1 | [83] |
| Hairless male mouse (6-week-old) exposed to UVB (60–90 mJ/cm2) 3x/week for 12 weeks | Oral administration | 100 mg/kg/day hesperidin, purity ≥ 98% (commercial product, supplier not specified) | ↓ Wrinkle formation ↓ Trans epidermal water loss ↓ Cytokine expression ↓ Epidermal hyperplasia | ↓ MMP-9 ↓ Phosphorylation of MAPK and extracellular signal-regulated kinases | [84] |
| HR-1 hairless mouse exposed to UVB (60–90 mJ/cm2), 3×/week for 12 weeks | Oral administration | 100 mg/kg/day hesperidin, purity ≥ 98% (commercial product, supplier not specified) | ↓ Cutaneous neovascularization | ↓ VEGF ↓ MMP-9 and MMP-13 ↓ Hypoxia-inducible factor 1 (HIF-1) alpha | [76] |
| Swiss albino mouse exposed to UVB (290–320 nm) for 5 weeks | Topical application | Citrus sinensis peel extract (ethanolic commercial extract; 15.53 mg% hesperidin), formulated in lipid nanoparticle cream | Slows down skin photoaging and wrinkles ↑ Antioxidant activity | ↑ Collagen and SOD ↓ PGE2, COX2, MMP1, collagenase, and elastase activities. | [17] |
| C57BL/6 and CISD2-KO mice, exposed to UVB (349 mJ/cm2) daily for 5 days during hesperetin treatment | Intraperitoneal injection | 2-day pretreatment, then treatment for 5 days during UV exposure with 10 mg/kg/day pure hesperetin, purity ≥ 95% (Sigma-Aldrich) for 7 days | Skin protection from photoaging (thickness) | ↑ CISD2 expression | [40] |
| Rat exposed to UVA-UVB for 5 consecutive days | Topical application | Pretreatment with 10% hesperetin-based hydrogel, once daily (commercial product, supplier not specified) | Protection of dermal–epidermal tissue ↓ Erythema ↓ Lipid peroxidation | ↓ Myeloperoxidase ↑ catalase and superoxide dismutase | [85] |
| Model | Route | Treatment | Effects | Mechanisms | Ref. |
|---|---|---|---|---|---|
| ECM enzyme activity | In vitro | Hesperidin and hesperetin purity ≥ 98% (Sigma-Aldrich) dissolved in ethanol (<1% final concentration) and tested in solution at 0.5–100 µM | Inhibition of hyaluronidase and elastase by 60–70% | Not determined | [62] |
| ECM enzyme activity | In vitro | Commercial cream formulation containing hesperidin (0.08–0.90 mmol/L); tested on human skin samples for enzyme assay. Supplier of hesperidin not specified | Interaction with collagenase and its inhibition | Zinc chelation | [16] |
| Physiologically aged fibroblasts | In vitro | Hesperidin and hesperetin ≥ 98% purity, commercial grade (Sigma-Aldrich), applied to cells at 0.5–100 µM (final ethanol concentration <1%) | Anti-aging | ↓ MMP-1 and MMP-2 ↓ Elastase and hyaluronidase | [62] |
| Skin barrier test in hairless mice | Topical application | 2% hesperidin cream, ≥98% purity (cosmetic grade, Procter & Gamble), applied topically twice daily for 6 days | Stimulation of proliferation, epidermal differentiation, and increased secretion of lamellar bodies | ↑ Barrier function measured as transepidermal water loss | [113] |
| Skin barrier test in corticosteroid-treated mice | Topical application | 2% hesperidin cream, ≥98% purity (cosmetic grade, Procter & Gamble), applied topically 1 h after 0.05% clobetasol propionate, twice daily for 9 days | ↑ Proliferation ↑ Barrier function ↓ pH skin surface | ↑ Expression of flaggrin ↑ Formation of lamellar bodies ↑ β-glucocerebrosidase ↑ Glutathione reductase | [114] |
| Skin barrier test in aged mice | Topical application | 2% hesperidin cream, ≥98% purity (Procter & Gamble, cosmetic grade), applied topically twice daily for 9 days | Anti-aging ↑ barrier function ↓ Skin surface pH | ↑ Expression of ABCA12, NHE1, and PLA2 ↑ Formation of lamellar bodies | [34] |
| Healing of skin wounds in diabetic rats | Oral administration | Hesperidin (25–100 mg/kg/day); administered once daily for 21 days after skin injury in diabetic rats (source and purity not reported) | ↓ Wound closure time Improvement of skin architecture | ↑ mRNA expression of VEGF, Ang-1/Tie-2, TGF-β, and Smad-2/3 ↑ SOD and GSH ↓ Levels of MDA and NO | [115] |
| Healing of skin wounds in diabetic rats | Oral administration | Hesperidin (10–80 mg/kg/day) for 20 consecutive days after wound induction in diabetic rats (source and purity not reported) | Healing with wound reduction | ↑ VEGFR1 and VEGFR2 levels ↓ TNF-α, IL-6 ↑ SOD and GSH ↓ MDA levels | [116] |
| Healing of skin wounds in mice | Topical application | 5% hesperidin or 5% naringin hydrogel (source not reported); applied once daily after wound induction | Reduced the average wound healing time ↓ Lipid peroxidation | ↑ Collagen synthesis ↑ GSH and SOD ↓ Expression of NF-kappaB and COX-2 | [117] |
| Healing of skin wounds in gamma-irradiated mice | Oral administration | Hesperidin (100 mg/kg), commercial grade (Sigma-Aldrich); administered orally 1 h prior to irradiation | ↑ Contraction of the wound ↓ Wound healing time | ↑ NO ↑ DNA synthesis ↑ Collagen ↑ Density of vessels and fibroblasts | [118] |
| Healing of skin wounds in rabbits | Topical application | Hesperidin 1% (formulation, source, and purity not reported); applied topically on ear wounds in rabbits | Scar was softer and lighter in color | ↓ Number of fibroblasts, capillaries, and inflammatory cells | [119] |
| Human keratinocytes (HaCaT line) exposed to H2O2 (48 h) | In vitro | Hesperidin 20 µg/mL, ≥98% purity (Sigma-Aldrich), 2 h pre-incubation | ↓ IL-8 (protein and mRNA) ↓ TNF-α (protein and mRNA) ↓ COX-2 expression | ↓ NF-κB, phosphorylated IκBα, and phosphorylated MAPK p38 | [120] |
| Macrophages RAW 264.7 cell line (mice) | In vitro | Hesperidin (5–250 µg/mL), incubated with LPS-stimulated RAW 264.7 macrophages (commercial source not specified) | ↓ NO production induced by lipopolysaccharide | Not determined | [121] |
| Macrophages RAW 264.7 cell line (mice) | In vitro | Hesperidin or hesperetin (40–100 µM), commercial grade (Sigma-Aldrich); 30 min pre-treatment before LPS stimulation | ↓ Oxidative stress ↓ PGE2 ↓ COX-2 expression ↓ NO production | ↓ Activation of NF-κB ↓ Phosphorylation of JNK1/2 and p38 ↓ IκBα ↓ mRNA of iNOS | [122] |
| NC/Nga mouse (spontaneous atopic dermatitis) | Oral administration | Hesperidin or α-glucopyranosyl hesperidin (0.1% in diet for 8 weeks); source and purity not reported | ↓ IgE ↓ Dermatitis symptoms | ↓ IL-17, IFN-γ ↓ CTLA4 | [123] |
| Human skin explants (rosacea pattern) | In vitro | HMC 0.2 mg/mL, pre-treatment of human skin explants followed by Substance P stimulation (24 h). Source and purity not reported | ↓ Proportion of dilated vessels ↓ Total surface area of blood vessels | ↓ Production of IL-8 | [100] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bellavite, P.; Imbriano, A. Skin Photoaging and the Biological Mechanism of the Protective Effects of Hesperidin and Derived Molecules. Antioxidants 2025, 14, 788. https://doi.org/10.3390/antiox14070788
Bellavite P, Imbriano A. Skin Photoaging and the Biological Mechanism of the Protective Effects of Hesperidin and Derived Molecules. Antioxidants. 2025; 14(7):788. https://doi.org/10.3390/antiox14070788
Chicago/Turabian StyleBellavite, Paolo, and Alice Imbriano. 2025. "Skin Photoaging and the Biological Mechanism of the Protective Effects of Hesperidin and Derived Molecules" Antioxidants 14, no. 7: 788. https://doi.org/10.3390/antiox14070788
APA StyleBellavite, P., & Imbriano, A. (2025). Skin Photoaging and the Biological Mechanism of the Protective Effects of Hesperidin and Derived Molecules. Antioxidants, 14(7), 788. https://doi.org/10.3390/antiox14070788