
Impact of SGLT2i on Cardiac Remodeling and the Soleus Muscle of Infarcted Rats

 , , ,
, , ,  ,
,  , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Echocardiographic Evaluation
2.3. Tissue Collection
2.4. Histological Analysis
2.5. Antioxidant Enzyme Activity
2.6. Oxidative Stress Analysis
2.6.1. Malondialdehyde
2.6.2. Protein Carbonylation
2.7. Metabolic Enzyme Activity
2.8. Protein Expression
2.9. Statistical Analysis
3. Results
3.1. Experimental Groups, Anatomical Variables, and Echocardiogram Data
3.2. SGLT2 Inhibition Preserves Trophism Change Caused by Myocardial Infarction in Oxidative Skeletal Muscle
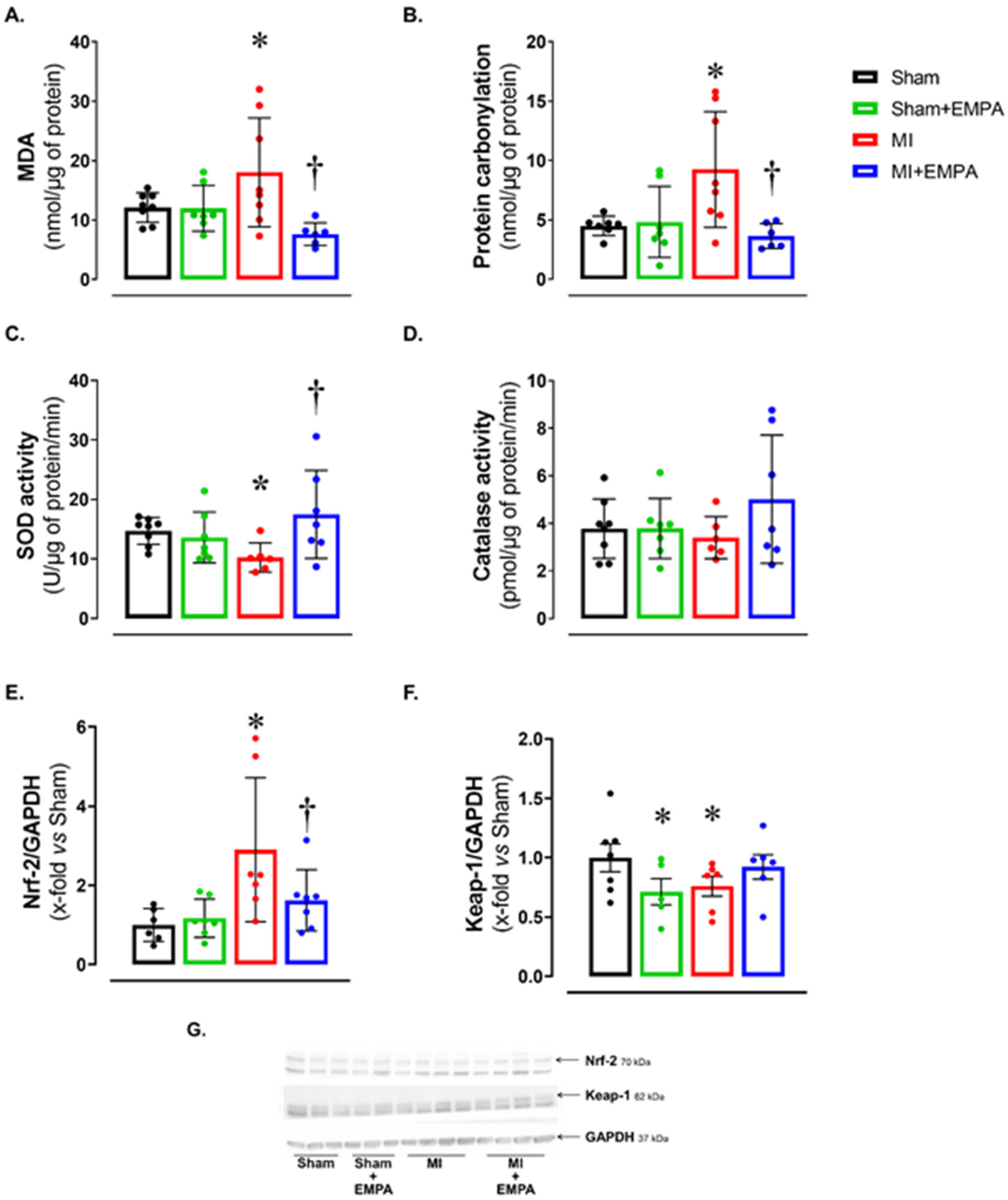
3.3. SGLT2 Inhibition Attenuates Oxidative Stress in the Skeletal Muscle of Infarcted Rats
3.4. Impact of Myocardial Infarction on Skeletal Muscle Energy Metabolism and the Restorative Effects of SGLT2 Inhibition
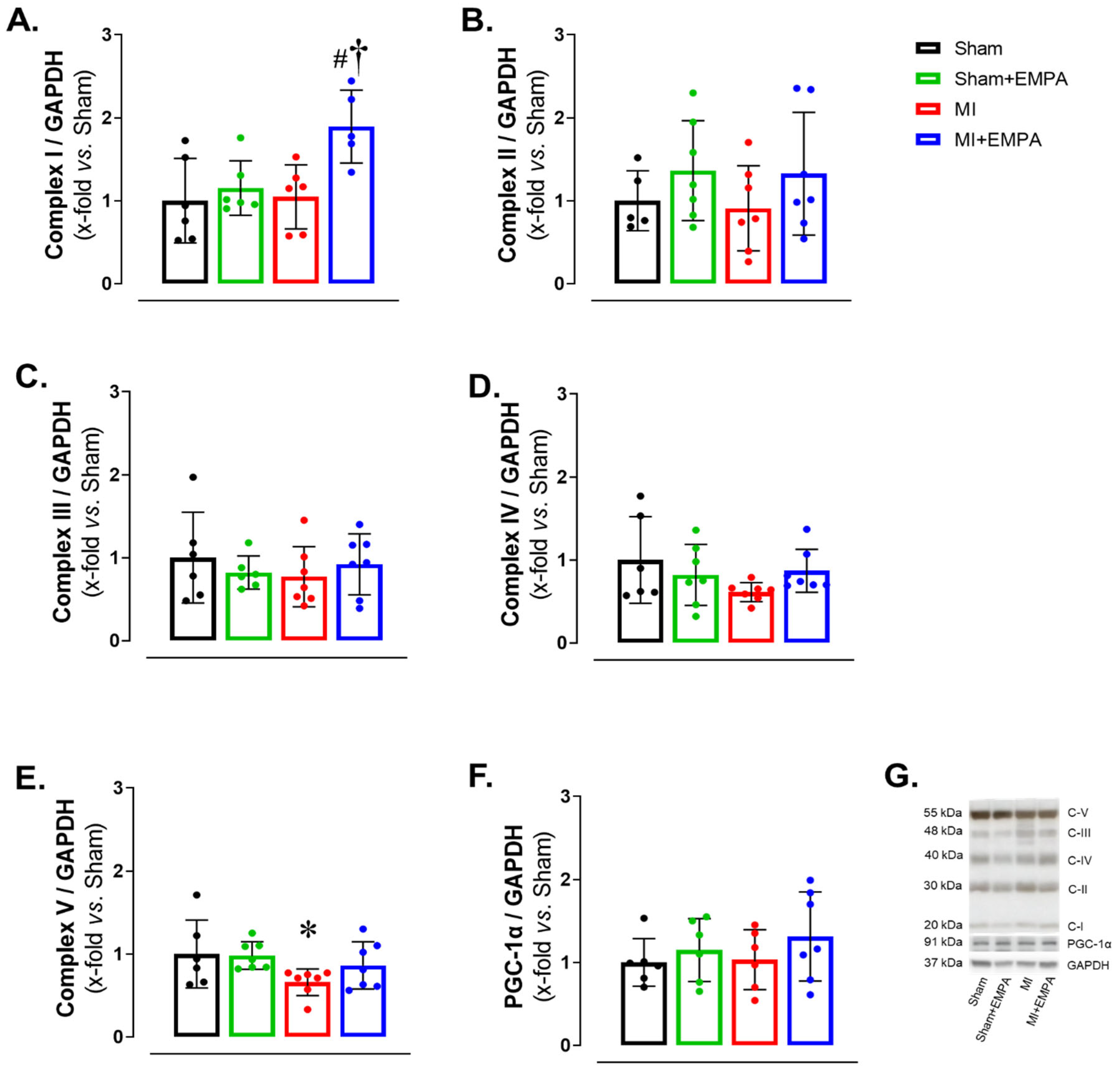
3.5. SGLT2 Inhibition of Mitochondrial Function in Oxidative Skeletal Muscle
3.6. SGLT2 Inhibition Improves Anabolic and Mitigates Catabolic Signaling in Oxidative Skeletal Muscle
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Akt | protein kinase B |
| ANOVA | analysis of variance |
| AO | aorta diameter |
| BHADH | beta-hydroxy-acyl dehydrogenase |
| BW | body weight |
| CS | citrate synthase |
| CSA | cross-sectional area |
| DNPH | 2,4-dinitrophenylhydrazine derivatizing agent |
| E | early diastolic mitral inflow velocity |
| E’ | tissue Doppler imaging of the systolic velocity of the mitral annulus |
| EF | ejection fraction |
| EMPA | empagliflozin |
| End-diast. area | LV end-diastolic area |
| End-sist. area | LV end-systolic area |
| FoxO3 | forkhead box O3 |
| G6PDH | glucose-6 phosphate dehydrogenase |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase |
| HF | heart failure |
| HK | hexokinase |
| i.p. | intraperitoneal |
| IGF-1 | insulin-like growth factor type 1 |
| IVRT | isovolumetric relaxation time |
| IVRT/R-R | IVRT normalized to heart rate |
| Keap-1 | Kelch-like ECH-associated protein 1 |
| LA | left atrial diameter |
| LV | left ventricle |
| LVDD | left ventricular (LV) diastolic diameters |
| LVSD | left ventricular (LV) systolic diameters |
| MAFbx | muscle atrophy F-box. |
| MDA | malondialdehyde |
| MI | myocardial infarction |
| MI+EMPA | MI treated with empagliflozin |
| mTOR | mammalian target of rapamycin |
| MuRF-1 | muscle RING-finger protein-1 |
| MyHC | myosin heavy chain |
| Nrf-2 | nuclear factor erythroid 2-related factor 2 |
| p70S6K | ribosomal protein S6 kinase beta-1 |
| PFK | phosphofructokinase |
| PGC-1α | peroxisomal proliferators-activated receptor γ-coactivator-1α |
| PK | pyruvate kinase |
| PWSV | posterior wall shortening velocity |
| PWT | LV posterior wall thickness |
| RV | right ventricular |
| RWT | relative wall thickness |
| SD | standard |
| SGLT2 | sodium–glucose cotransporter 2 |
| Sham+EMPA | Sham treated with empagliflozin |
| Tei index | myocardial performance index |
References
- Martin, S.S.; Aday, A.W.; Allen, N.B.; Almarzooq, Z.I.; Anderson, C.A.M.; Arora, P.; Avery, C.L.; Baker-Smith, C.M.; Bansal, N.; Beaton, A.Z.; et al. 2025 Heart Disease and Stroke Statistics: A report of US and global data from the American Heart Association. Circulation 2025, 151, e41–e660. [Google Scholar] [PubMed]
- Del Buono, M.G.; Arena, R.; Borlaug, B.A.; Carbone, S.; Canada, J.M.; Kirkman, D.L.; Garten, R.; Rodriguez-Miguelez, P.; Guazzi, M.; Lavie, C.J.; et al. Exercise intolerance in patients with heart failure: JACC State-of-the-Art Review. J. Am. Coll. Cardiol. 2019, 73, 2209–2225. [Google Scholar] [CrossRef] [PubMed]
- von Haehling, S. The wasting continuum in heart failure: From sarcopenia to cachexia. Proc. Nutr. Soc. 2015, 74, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Damluji, A.A.; Alfaraidhy, M.; AlHajri, N.; Rohant, N.N.; Kumar, M.; Al Malouf, C.; Bahrainy, S.; Kwak, M.J.; Batchelor, W.B.; Forman, D.E.; et al. Sarcopenia and cardiovascular diseases. Circulation 2023, 147, 1534–1553. [Google Scholar] [CrossRef]
- Lv, J.; Li, Y.; Shi, S.; Xu, X.; Wu, H.; Zhang, B.; Song, Q. Skeletal muscle mitochondrial remodeling in heart failure: An update on mechanisms and therapeutic opportunities. Biomed. Pharmacother. 2022, 155, 113833. [Google Scholar] [CrossRef]
- Wood, N.; Straw, S.; Scalabrin, M.; Roberts, L.D.; Witte, K.K.; Bowen, T.S. Skeletal muscle atrophy in heart failure with diabetes: From molecular mechanisms to clinical evidence. ESC Hear. Fail. 2021, 8, 3–15. [Google Scholar] [CrossRef]
- Heidenreich, P.A.; Bozkurt, B.; Aguilar, D.; Allen, L.A.; Byun, J.J.; Colvin, M.M.; Deswal, A.; Drazner, M.H.; Dunlay, S.M.; Evers, L.R.; et al. 2022 AHA/ACC/HFSA Guideline for the management of heart failure: Executive Summary: A report of the American College of Cardiology/American Heart Association Joint Committee on Clinical Practice Guidelines. J. Am. Coll. Cardiol. 2022, 79, 1757–1780. [Google Scholar] [CrossRef]
- Braunwald, E. SGLT2 inhibitors: The statins of the 21st century. Eur. Heart J. 2022, 43, 1029–1030. [Google Scholar] [CrossRef]
- Xie, Y.; Wei, Y.; Li, D.; Pu, J.; Ding, H.; Zhang, X. Mechanisms of SGLT2 inhibitors in heart failure and their clinical value. J. Cardiovasc. Pharmacol. 2023, 81, 4–14. [Google Scholar] [CrossRef]
- Takada, S.; Sabe, H.; Kinugawa, S. Treatments for skeletal muscle abnormalities in heart failure: Sodium-glucose transporter 2 and ketone bodies. Am. J. Physiol. Heart Circ. Physiol. 2022, 322, H117–H128. [Google Scholar] [CrossRef]
- Bodnar, P.; Mazurkiewicz, M.; Chwalba, T.; Romuk, E.; Ciszek-Chwalba, A.; Jacheć, W.; Wojciechowska, C. The impact of pharmacotherapy for heart failure on oxidative stress-role of new drugs, flozins. Biomedicines 2023, 11, 2236. [Google Scholar] [CrossRef] [PubMed]
- Wood, N.; Straw, S.; Cheng, C.W.; Hirata, Y.; Pereira, M.G.; Gallagher, H.; Egginton, S.; Ogawa, W.; Wheatcroft, S.B.; Witte, K.K.; et al. Sodium-glucose cotransporter 2 inhibitors influence skeletal muscle pathology in patients with heart failure and reduced ejection fraction. Eur. J. Heart Fail. 2024, 26, 925–935. [Google Scholar] [CrossRef] [PubMed]
- Nambu, H.; Takada, S.; Fukushima, A.; Matsumoto, J.; Kakutani, N.; Maekawa, S.; Shirakawa, R.; Nakano, I.; Furihata, T.; Katayama, T.; et al. Empagliflozin restores lowered exercise endurance capacity via the activation of skeletal muscle fatty acid oxidation in a murine model of heart failure. Eur. J. Pharmacol. 2020, 866, 172810. [Google Scholar] [CrossRef]
- Winzer, E.B.; Schauer, A.; Langner, E.; Augstein, A.; Goto, K.; Männel, A.; Barthel, P.; Jannasch, A.; Labeit, S.; Mangner, N.; et al. Empagliflozin preserves skeletal muscle function in a HFpEF rat model. Int. J. Mol. Sci. 2022, 23, 10989. [Google Scholar] [CrossRef]
- Okoshi, K.; Cezar, M.D.M.; Polin, M.A.M.; Paladino, J.R.; Martinez, P.F.; Oliveira, S.A., Jr.; Lima, A.R.R.; Damatto, R.L.; Paiva, S.A.R.; Zornoff, L.A.M.; et al. Influence of intermittent fasting on myocardial infarction-induced cardiac remodeling. BMC Cardiovasc. Disord. 2019, 19, 126. [Google Scholar] [CrossRef]
- Cezar, M.D.M.; Damatto, R.L.; Martinez, P.F.; Lima, A.R.R.; Campos, D.H.S.; Rosa, C.M.; Guizoni, D.M.; Bonomo, C.; Cicogna, A.C.; Gimenes, R.; et al. Aldosterone blockade reduces mortality without changing cardiac remodeling in spontaneously hypertensive rats. Cell Physiol. Biochem. 2013, 32, 1275–1287. [Google Scholar] [CrossRef]
- Zornoff, L.A.M.; Matsubara, L.S.; Matsubara, B.B.; Okoshi, M.P.; Okoshi, K.; Dal Pai-Silva, M.; Carvalho, R.F.; Cicogna, A.C.; Padovani, C.R.; Novelli, E.L.; et al. Beta-carotene supplementation attenuates cardiac remodeling induced by one-month tobacco-smoke exposure in rats. Toxicol. Sci. 2006, 90, 259–266. [Google Scholar] [CrossRef]
- Reyes, D.R.A.; Gomes, M.J.; Rosa, C.M.; Pagan, L.U.; Zanati, S.G.; Damatto, R.L.; Rodrigues, E.A.; Carvalho, R.F.; Fernandes, A.A.H.; Martinez, P.F.; et al. Exercise during transition from compensated left ventricular hypertrophy to heart failure in aortic stenosis rats. J. Cell Mol. Med. 2019, 23, 1235–1245. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.J.; Pagan, L.U.; Lima, A.R.R.; Reyes, D.R.A.; Martinez, P.F.; Damatto, F.C.; Pontes, T.H.D.; Rodrigues, E.A.; Souza, L.M.; Tosta, I.F.; et al. Effects of aerobic and resistance exercise on cardiac remodelling and skeletal muscle oxidative stress of infarcted rats. J. Cell Mol. Med. 2020, 24, 5352–5362. [Google Scholar] [CrossRef]
- Souza, L.M.; Gomes, M.J.; Brandao, B.B.; Pagan, L.U.; Gatto, M.; Damatto, F.C.; Rodrigues, E.A.; Pontes, T.H.D.; Borim, P.A.; Fernandes, A.A.H.; et al. Effects of resistance exercise on slow-twitch soleus muscle of infarcted rats. Antioxidants 2023, 12, 291. [Google Scholar] [CrossRef]
- Reyes, D.R.A.; Gomes, M.J.; Rosa, C.M.; Pagan, L.U.; Damatto, F.C.; Damatto, R.L.; Depra, I.; Campos, D.H.S.; Fernandez, A.A.H.; Martinez, P.F.; et al. N-Acetylcysteine influence on oxidative stress and cardiac remodeling in rats during transition from compensated left ventricular hypertrophy to heart failure. Cell Physiol. Biochem. 2017, 44, 2310–2321. [Google Scholar] [CrossRef] [PubMed]
- Mattei, L.; Francisqueti-Ferron, F.V.; Garcia, J.L.; Ferron, A.J.T.; De Almeida Silva, C.C.V.; Gregolin, C.S.; Nakandakare-Maia, E.T.; Silva, J.C.P.S.; Moreto, F.; Minatel, I.O.; et al. Antioxidant and anti-inflammatory properties of gamma- oryzanol attenuates insulin resistance by increasing GLUT- 4 expression in skeletal muscle of obese animals. Mol. Cell Endocrinol. 2021, 537, 111423. [Google Scholar] [CrossRef]
- Lee-Young, R.S.; Hoffman, N.J.; Murphy, K.T.; Henstridge, D.C.; Samocha-Bonet, D.; Siebel, A.L.; Iliades, P.; Zivanovic, B.; Hong, Y.H.; Colgan, T.D.; et al. Glucose-6-phosphate dehydrogenase contributes to the regulation of glucose uptake in skeletal muscle. Mol. Metab. 2016, 5, 1083–1091. [Google Scholar] [CrossRef] [PubMed]
- Guizoni, D.M.; Oliveira-Junior, S.A.; Noor, S.L.R.; Pagan, L.U.; Martinez, P.F.; Lima, A.R.R.; Gomes, M.J.; Damatto, R.L.; Cezar, M.D.M.; Bonomo, C.; et al. Effects of late exercise on cardiac remodeling and myocardial calcium handling proteins in rats with moderate and large size myocardial infarction. Int. J. Cardiol. 2016, 221, 406–412. [Google Scholar] [CrossRef]
- Damatto, R.L.; Lima, A.R.R.; Martinez, P.F.; Cezar, M.D.M.; Okoshi, K.; Okoshi, M.P. Myocardial myostatin in spontaneously hypertensive rats with heart failure. Int. J. Cardiol. 2016, 215, 384–387. [Google Scholar] [CrossRef] [PubMed]
- Anker, S.D.; Ponikowski, P.; Varney, S.; Chua, T.P.; Clark, A.L.; Webb-Peploe, K.M.; Harrington, D.; Kox, W.J.; Poole-Wilson, P.A.; Coats, A.J.; et al. Wasting as independent risk factor for mortality in chronic heart failure. Lancet 1997, 349, 1050–1053. [Google Scholar] [CrossRef]
- Takada, S.; Sabe, H.; Kinugawa, S. Abnormalities of skeletal muscle, adipocyte tissue, and lipid metabolism in heart failure: Practical therapeutic targets. Front. Cardiovasc. Med. 2020, 7, 79. [Google Scholar] [CrossRef] [PubMed]
- Rosca, M.G.; Okere, I.A.; Sharma, N.; Stanley, W.C.; Recchia, F.A.; Hoppel, C.L. Altered expression of the adenine nucleotide translocase isoforms and decreased ATP synthase activity in skeletal muscle mitochondria in heart failure. J. Mol. Cell Cardiol. 2009, 46, 927–935. [Google Scholar] [CrossRef]
- Wüst, R.C.I.; de Vries, H.J.; Wintjes, L.T.; Rodenburg, R.J.; Niessen, H.W.M.; Stienen, G.J.M. Mitochondrial complex I dysfunction and altered NAD(P)H kinetics in rat myocardium in cardiac right ventricular hypertrophy and failure. Cardiovasc. Res. 2016, 111, 362–372. [Google Scholar] [CrossRef]
- Sartori, R.; Romanello, V.; Sandri, M. Mechanisms of muscle atrophy and hypertrophy: Implications in health and disease. Nat. Commun. 2021, 12, 330. [Google Scholar] [CrossRef]
- Xie, K.; Sugimoto, K.; Tanaka, M.; Akasaka, H.; Fujimoto, T.; Takahashi, T.; Onishi, Y.; Minami, T.; Yoshida, S.; Takami, Y.; et al. Effects of luseogliflozin treatment on hyperglycemia-induced muscle atrophy in rats. J. Clin. Biochem. Nutr. 2023, 72, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Barreiro, E. Role of protein carbonylation in skeletal muscle mass loss associated with chronic conditions. Proteomes 2016, 4, 18. [Google Scholar] [CrossRef] [PubMed]
- Bellezza, I.; Giambanco, I.; Minelli, A.; Donato, R. Nrf2-Keap1 signaling in oxidative and reductive stress. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Baird, L.; Yamamoto, M. The molecular mechanisms regulating the KEAP1-NRF2 pathway. Mol. Cell Biol. 2020, 40, e00099-20. [Google Scholar] [CrossRef]
- Zamani, P.; Proto, E.A.; Wilson, N.; Fazelinia, H.; Ding, H.; Spruce, L.A.; Davila, A., Jr.; Hanff, T.C.; Mazurek, J.A.; Prenner, S.B.; et al. Multimodality assessment of heart failure with preserved ejection fraction skeletal muscle reveals differences in the machinery of energy fuel metabolism. ESC Heart Fail. 2021, 8, 2698–2712. [Google Scholar] [CrossRef]
- Schauer, A.; Adams, V.; Kämmerer, S.; Langner, E.; Augstein, A.; Barthel, P.; Männel, A.; Fabig, G.; Alves, P.K.N.; Günscht, M.; et al. Empagliflozin improves diastolic function in HFpEF by restabilizing the mitochondrial respiratory chain. Circ. Heart Fail. 2024, 17, e011107. [Google Scholar] [CrossRef]
- Radlinger, B.; Ress, C.; Folie, S.; Salzmann, K.; Lechuga, A.; Weiss, B.; Salvenmoser, W.; Graber, M.; Hirsch, J.; Holfeld, J.; et al. Empagliflozin protects mice against diet-induced obesity, insulin resistance and hepatic steatosis. Diabetologia 2023, 66, 754–767. [Google Scholar] [CrossRef]
- Sanchez, A.M.J.; Candau, R.B.; Bernardi, H. FoxO transcription factors: Their roles in the maintenance of skeletal muscle homeostasis. Cell Mol. Life Sci. 2014, 71, 1657–1671. [Google Scholar] [CrossRef]
- Braunwald, E. Gliflozins in the management of cardiovascular disease. N. Engl. J. Med. 2022, 386, 2024–2034. [Google Scholar] [CrossRef]
- Castro, P.C.; Santos-Rios, T.M.; Martins, F.L.; Crajoinas, R.O.; Caetano, M.V.; Lessa, L.M.A.; Luchi, W.M.; McCormick, J.A.; Girardi, A.C.C. Renal upregulation of NCC counteracts empagliflozin-mediated NHE3 inhibition in normotensive but not in hypertensive male rat. Am. J. Physiol. Cell Physiol. 2024, 326, C1573–C1589. [Google Scholar] [CrossRef]
- Masuda, T.; Yoshida, M.; Onaka, T.; Nagata, D. Water and sodium conservation response induced by SGLT2 inhibitor ipragliflozin in Dahl salt-sensitive hypertensive rats. Hypertens. Res. 2024, 47, 3173–3181. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sham (n = 10) | Sham + EMPA (n = 12) | MI (n = 09) | MI + EMPA (n = 10) | |
|---|---|---|---|---|
| BW (g) | 485 ± 47 | 464 ± 43 | 491 ± 35 | 465 ± 44 |
| LV (g) | 0.81 ± 0.07 | 0.78 ± 0.07 | 0.82 ± 0.22 | 0.87 ± 0.06 |
| LV/BW (mg/g) | 1.68 ± 0.14 | 1.68 ± 0.16 | 1.71 ± 0.52 | 1.82 ± 0.17 |
| RV (g) | 0.29 ± 0.05 | 0.26 ± 0.04 | 0.34 ± 0.14 | 0.35 ± 0.14 |
| RV/BW (mg/g) | 0.61 ± 0.08 | 0.57 ± 0.06 | 0.77 ± 0.23 * | 0.75 ± 0.27 |
| Atria (g) | 0.09 ± 0.04 | 0.10 ± 0.02 | 0.18 ± 0.04 * | 0.16 ± 0.03 # |
| Atria/BW (mg/g) | 0.19 ± 0.08 | 0.22 ± 0.05 | 0.37 ± 0.09 * | 0.34 ± 0.10 # |
| Sham (n = 10) | Sham + EMPA (n = 12) | MI (n = 09) | MI + EMPA (n = 10) | |
|---|---|---|---|---|
| LVDD (mm) | 8.31 ± 0.39 | 7.99 ± 0.32 | 10.13 ± 0.93 * | 10.02 ± 0.81 # |
| LVDD/BW (mg/kg) | 17.3 ± 1.90 | 17.3 ± 1.20 | 20.8 ± 2.97 * | 22.3 ± 2.56 # |
| LVSD (mm) | 4.21 ± 0.74 | 3.78 ± 0.57 | 8.09 ± 1.16 * | 7.61 ± 1.02 # |
| PWT (mm) | 1.41 (1.37–1.45) | 1.43 (1.38–1.45) | 1.77 (1.64–2.14) * | 1.83 (1.62–2.03) # |
| RWT | 0.34 ± 0.02 | 0.36 ± 0.02 | 0.37 ± 0.07 | 0.37 ± 0.06 |
| AO (mm) | 4.07 ± 0.08 | 4.04 ± 0.10 | 3.85 ± 0.15 * | 3.86 ± 0.10 # |
| LA (mm) | 5.62 ± 0.22 | 5.65 ± 0.32 | 7.69 ± 1.26 * | 6.85 ± 0.90 †# |
| LA/AO | 1.40 ± 0.05 | 1.40 ± 0.07 | 2.01 ± 0.36 * | 1.78 ± 0.24 †# |
| LA/BW (mm/kg) | 11.7 ± 1.4 | 12.2 ± 0.9 | 15.8 ± 3.2 * | 14.8 ± 1.9 # |
| End-diast. area (mm2) | 47 ± 7.1 | 48 ± 4.5 | 100 ± 16.1 * | 85 ± 10.1 †# |
| End-syst. area (mm2) | 17 ± 4.4 | 15 ± 3.7 | 65 ± 19.5 * | 63 ± 9.2 # |
| EF | 0.86 ± 0.06 | 0.89 ± 0.03 | 0.49 ± 0.08 * | 0.55 ± 0.11 # |
| PWSV (mm/s) | 41 ± 5.7 | 43 ± 3.6 | 24 ± 6.2 * | 28 ± 5.6 # |
| Tei index | 0.49 ± 0.12 | 0.46 ± 0.07 | 0.76 ± 0.15 * | 0.73 ± 0.15 # |
| IVRT (ms) | 26 ± 3.8 | 25 ± 2.1 | 34 ± 3.4 * | 32 ± 4.5 # |
| IVRT/R-R | 54 ± 6.5 | 51 ± 4.1 | 70 ± 6.4 * | 64 ± 10.4 # |
| E/E’ (average) | 19.3 ± 2.1 | 19.0 ± 2.4 | 22.4 ± 5.5 | 25.3 ± 5.1 # |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Souza, L.M.; Damatto, F.C.; Brandão, B.B.; Rodrigues, E.A.; Santos, A.C.C.; Silva, R.C.F.; Gatto, M.; Pagan, L.U.; Martinez, P.F.; Murata, G.M.; et al. Impact of SGLT2i on Cardiac Remodeling and the Soleus Muscle of Infarcted Rats. Antioxidants 2025, 14, 647. https://doi.org/10.3390/antiox14060647
Souza LM, Damatto FC, Brandão BB, Rodrigues EA, Santos ACC, Silva RCF, Gatto M, Pagan LU, Martinez PF, Murata GM, et al. Impact of SGLT2i on Cardiac Remodeling and the Soleus Muscle of Infarcted Rats. Antioxidants. 2025; 14(6):647. https://doi.org/10.3390/antiox14060647
Chicago/Turabian StyleSouza, Lidiane Moreira, Felipe Cesar Damatto, Bruna Brasil Brandão, Eder Anderson Rodrigues, Anna Clara Consorti Santos, Rafael Campos França Silva, Mariana Gatto, Luana Urbano Pagan, Paula Felippe Martinez, Gilson Masahiro Murata, and et al. 2025. "Impact of SGLT2i on Cardiac Remodeling and the Soleus Muscle of Infarcted Rats" Antioxidants 14, no. 6: 647. https://doi.org/10.3390/antiox14060647
APA StyleSouza, L. M., Damatto, F. C., Brandão, B. B., Rodrigues, E. A., Santos, A. C. C., Silva, R. C. F., Gatto, M., Pagan, L. U., Martinez, P. F., Murata, G. M., Zornoff, L. A. M., Gaiolla, P. S. A., Falcão-Pires, I., Okoshi, K., & Okoshi, M. P. (2025). Impact of SGLT2i on Cardiac Remodeling and the Soleus Muscle of Infarcted Rats. Antioxidants, 14(6), 647. https://doi.org/10.3390/antiox14060647