


Melatonin Promotes Muscle Growth and Redirects Fat Deposition in Cashmere Goats via Gut Microbiota Modulation and Enhanced Antioxidant Capacity

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Treatment
2.2. Carcass Trait Analysis
2.3. Meat Quality Evaluation
2.4. Determination of Chemical Composition of Muscles
2.5. Histological Analysis of Tissues
2.6. Blood Sampling and Biochemical Analyses
2.7. Gastrointestinal Microbiota Analysis
2.8. Statistical Analyses
3. Results
3.1. Slaughter Performance
3.2. Meat Quality and Muscle Composition
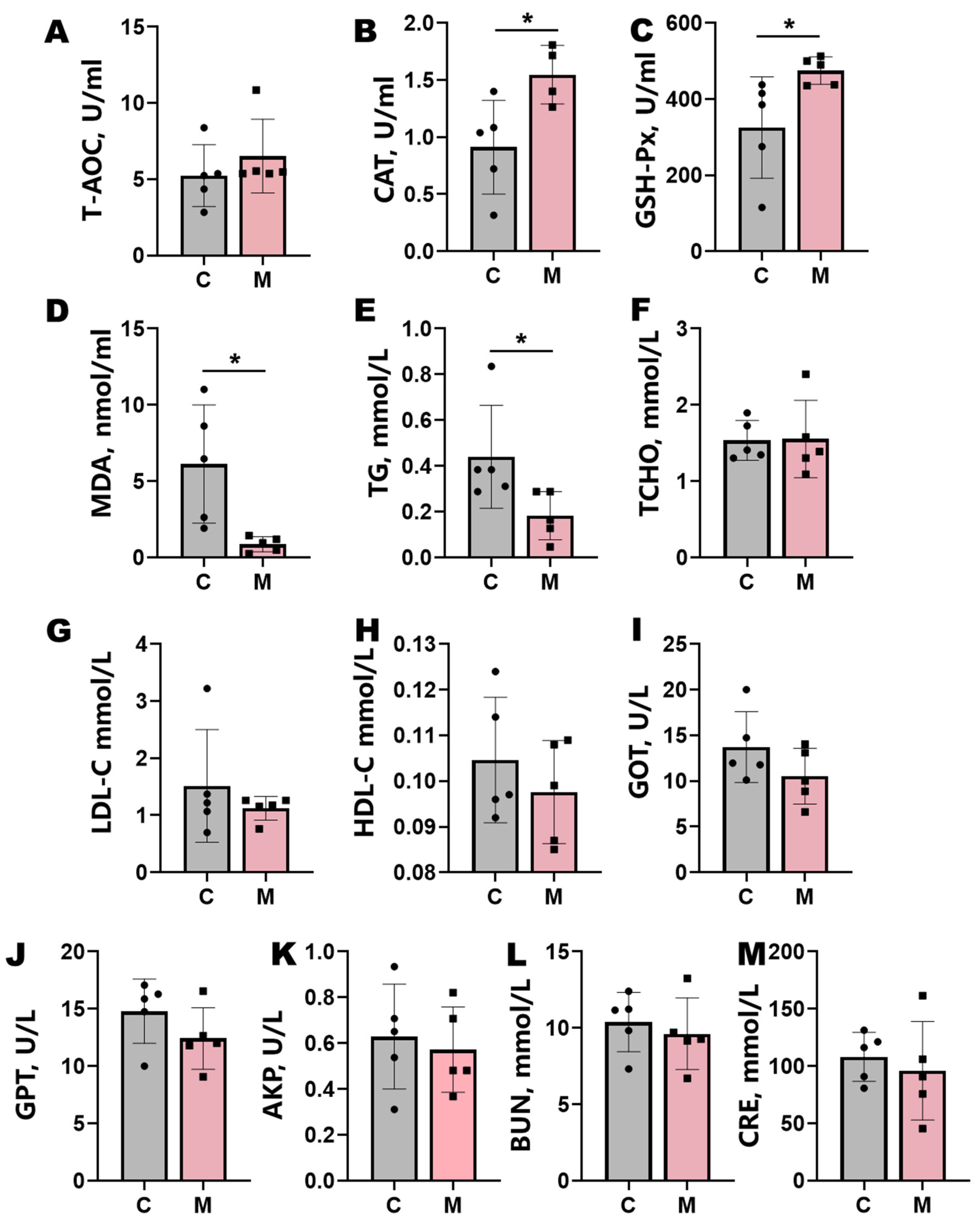
3.3. Blood Biochemical Parameters
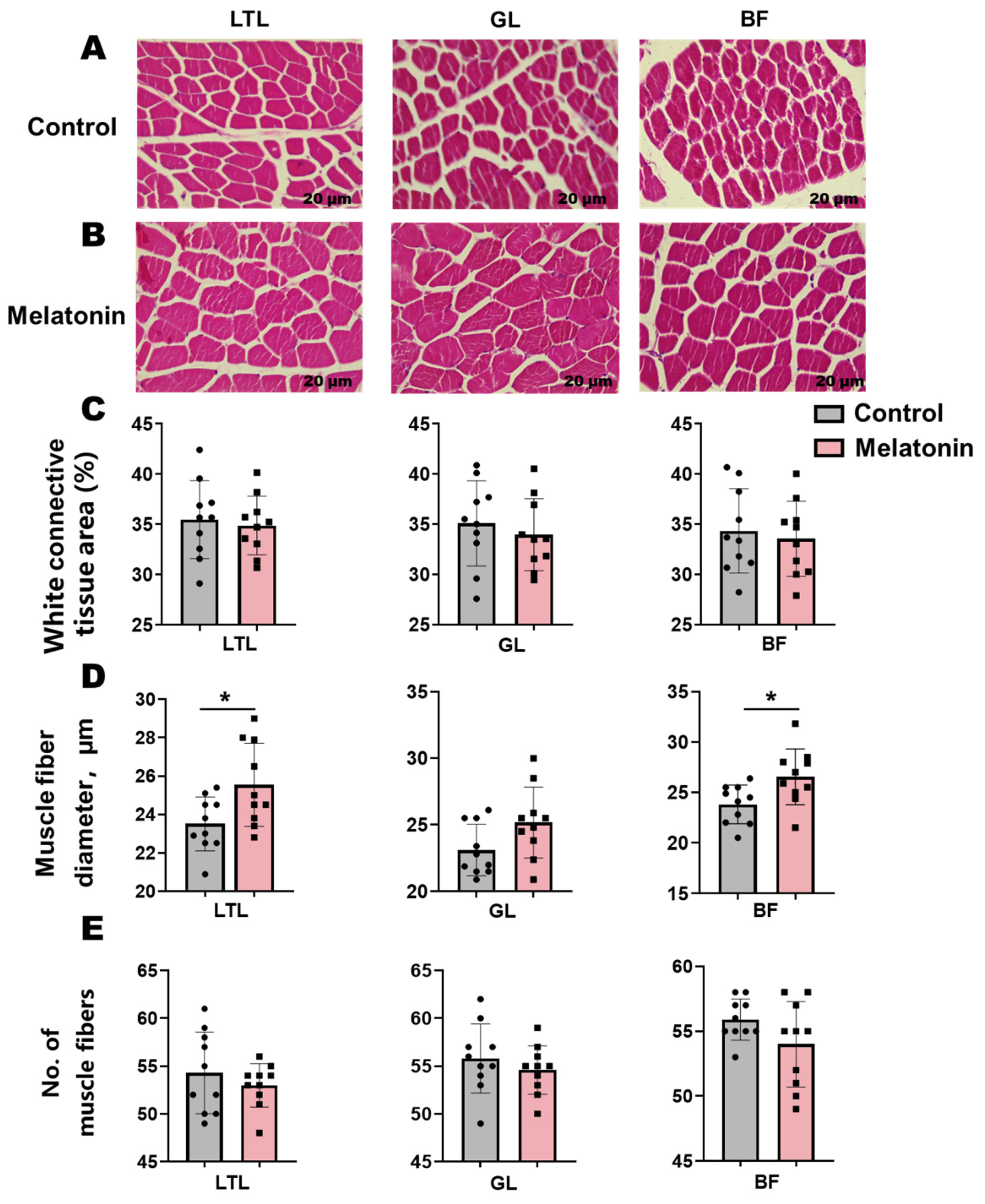
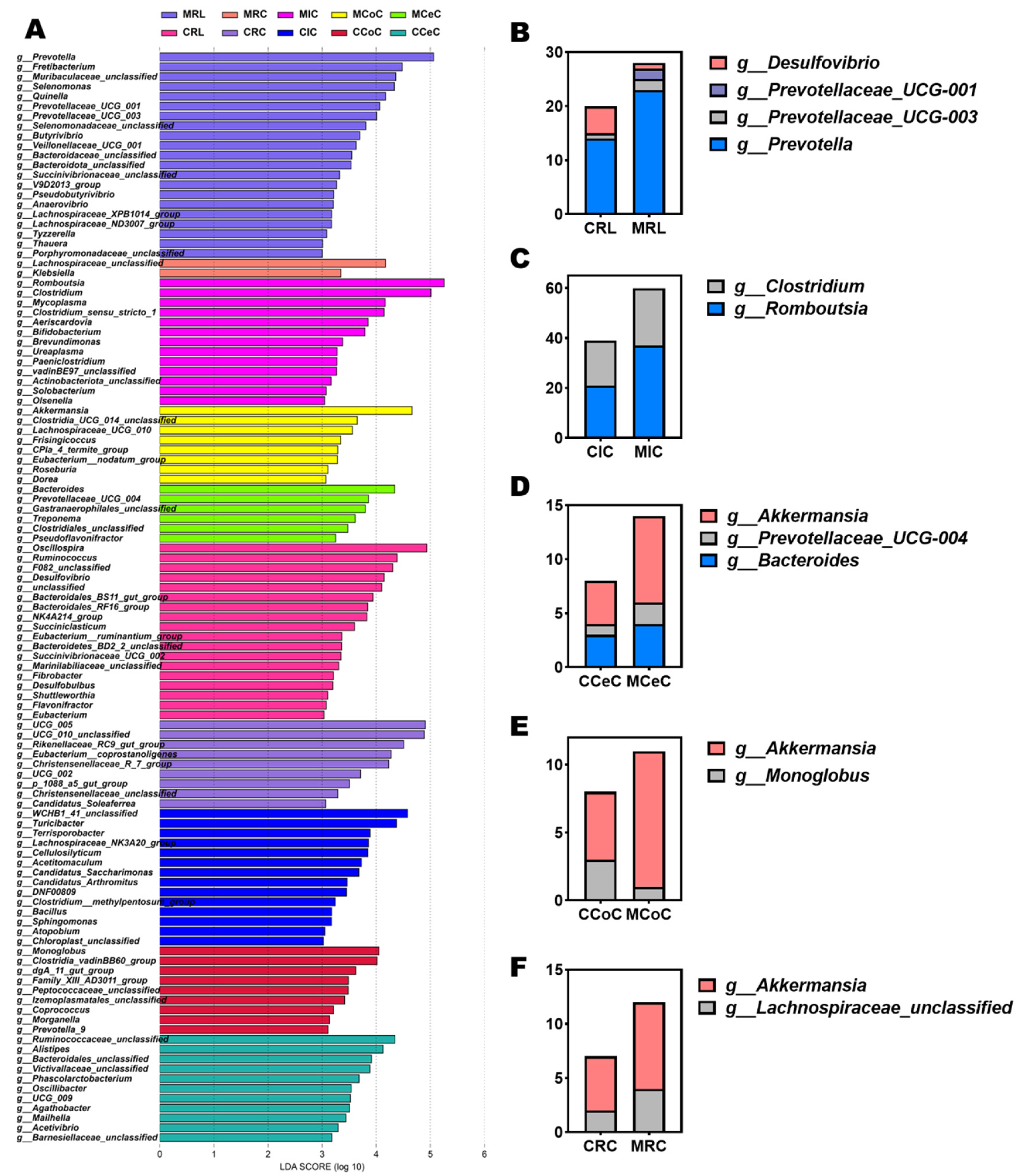
3.4. Gastrointestinal Histomorphology and Microbiome Composition
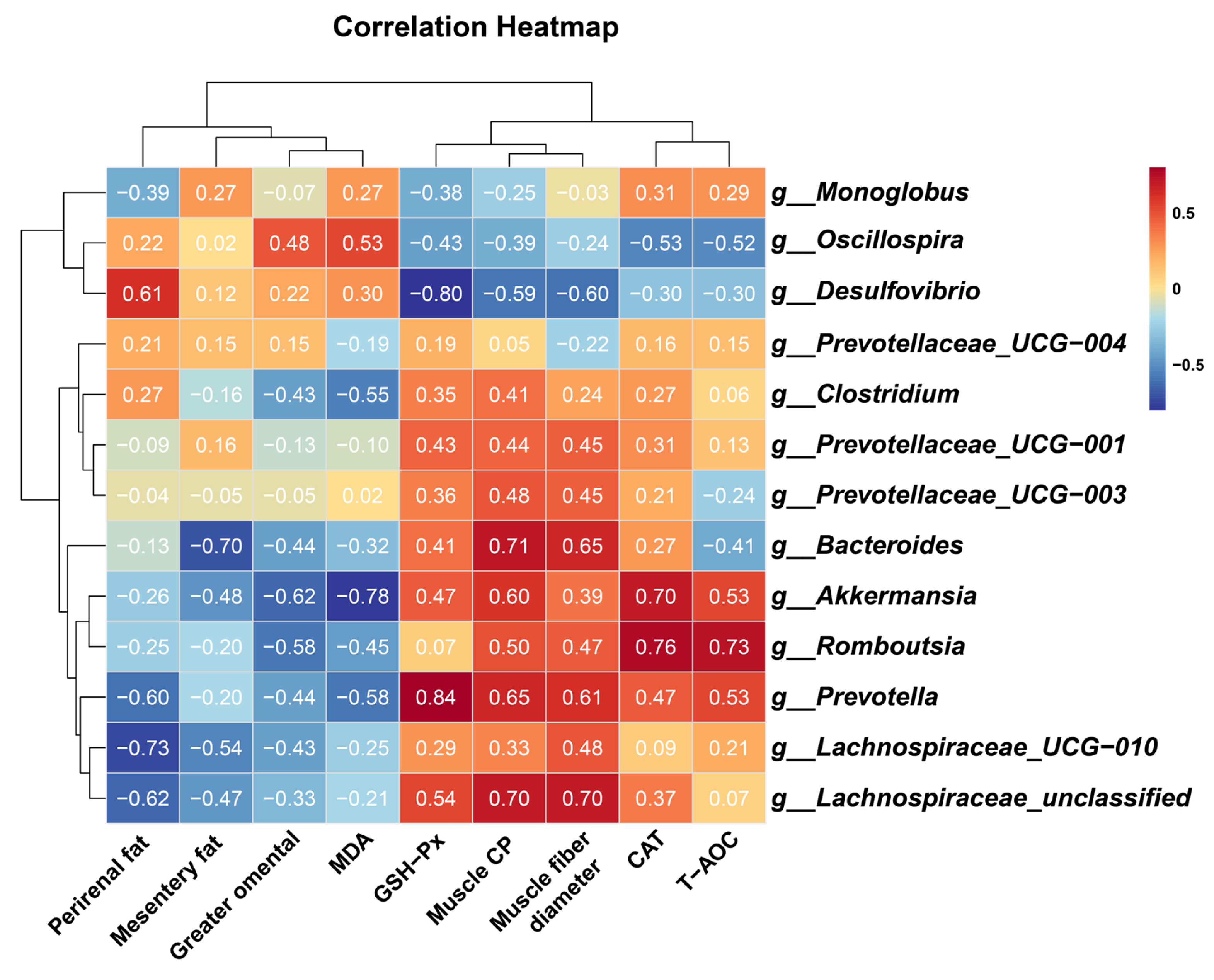
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, Y.; Kong, L.; Li, S.; Nie, L.; Gao, J.; Li, S.; Pan, Y.; Liu, Q.; Wang, Z. Correlation and regression analysis of GH and IGF-1 genes in Liaoning cashmere goats with body size and other production performance. J. Genet. Eng. Biotechnol. 2024, 22, 100440. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Su, Z.; Zhang, W. Melatonin’s role in hair follicle growth and development: A cashmere goat perspective. Int. J. Mol. Sci. 2025, 26, 2844. [Google Scholar] [CrossRef] [PubMed]
- Zagury, Y.; Ianovici, I.; Landau, S.; Lavon, N.; Levenberg, S. Engineered marble-like bovine fat tissue for cultured meat. Commun. Biol. 2022, 5, 927. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Wang, C.; Corazzin, M.; Zhang, T.; Zhai, M.; Zhang, Y.; Zhang, Q.; Zhao, X.; Jin, Y.; Sun, L. Effects of Limosilactobacillus reuteri on gastrointestinal microbiota, muscle fiber formation and meat quality of Sunit sheep. Food Sci. Anim. Prod. 2024, 2, 9240096. [Google Scholar] [CrossRef]
- Lee, S.H.; Joo, S.T.; Ryu, Y.C. Skeletal muscle fiber type and myofibrillar proteins in relation to meat quality. Meat Sci. 2010, 86, 166–170. [Google Scholar] [CrossRef]
- Picard, B.; Gagaoua, M. Muscle fiber properties in cattle and their relationships with meat qualities: An Overview. J. Agric. Food Chem. 2020, 68, 6021–6039. [Google Scholar] [CrossRef]
- Wang, L.; Valencak, T.G.; Shan, T. Fat infiltration in skeletal muscle: Influential triggers and regulatory mechanism. iScience 2024, 27, 109221. [Google Scholar] [CrossRef]
- Ma, W.; Wu, H.; Li, G.; Yan, L.; Wang, L.; Zhao, M.; Guan, S.; Xu, S.; Guo, X.; Liu, F.; et al. Melatonin promotes the growth and development of lambs by increasing growth hormone and testosterone, targeting on apoptosis signaling pathway and intestinal microflora. Front. Endocrinol. 2022, 13, 966120. [Google Scholar] [CrossRef]
- Chen, W.; Tu, Y.; Cai, P.; Wang, L.; Zhou, Y.; Liu, S.; Huang, Y.; Zhang, S.; Gu, X.; Yi, W.; et al. Melatonin supplementation promotes muscle fiber hypertrophy and regulates lipid metabolism of skeletal muscle in weaned piglets. J. Anim. Sci. 2023, 101, skad256. [Google Scholar] [CrossRef]
- Reiter, R.J.; Mayo, J.C.; Tan, D.X.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an antioxidant: Under promises but over delivers. J. Pineal. Res. 2016, 61, 253–278. [Google Scholar] [CrossRef]
- Yao, H.; Xie, Q.; Yang, Y.; Zhou, C.; Zeng, Z.; Zhang, W. Melatonin attenuates sepsis-induced muscle atrophy by regulating the PI3K/Akt signaling pathway. Int. Immunopharmacol. 2025, 144, 113619. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhang, X.; Wang, M.; Zhou, C.; Jiao, J.; Tan, Z. Enhancing metabolic efficiency through optimizing metabolizable protein profile in a time progressive manner with weaned goats as a model: Involvement of gut microbiota. Microbiol. Spectr. 2022, 10, e02545-21. [Google Scholar] [CrossRef] [PubMed]
- Luo, T.; Li, Y.; Zhang, W.; Liu, J.; Shi, H. Rumen and fecal microbiota profiles associated with immunity of young and adult goats. Front. Immunol. 2022, 13, 978402. [Google Scholar] [CrossRef] [PubMed]
- Simon, J.-C.; Marchesi, J.R.; Mougel, C.; Selosse, M.-A. Host-microbiota interactions: From holobiont theory to analysis. Microbiome 2019, 7, 5. [Google Scholar] [CrossRef]
- Zhen, Y.; Xi, Z.; Nasr Shaima, M.; He, F.; Han, M.; Yin, J.; Ge, L.; Chen, Y.; Wang, Y.; Wei, W.; et al. Multi-Omics reveals the impact of exogenous short-chain fatty acid infusion on rumen homeostasis: Insights into crosstalk between the microbiome and the epithelium in a goat model. Microbiol. Spectr. 2023, 11, e05343-22. [Google Scholar] [CrossRef]
- Ma, L.; Lyu, W.; Zeng, T.; Wang, W.; Chen, Q.; Zhao, J.; Zhang, G.; Lu, L.; Yang, H.; Xiao, Y. Duck gut metagenome reveals the microbiome signatures linked to intestinal regional, temporal development, and rearing condition. iMeta 2024, 3, e198. [Google Scholar] [CrossRef]
- Li, T.-T.; Chen, X.; Huo, D.; Arifuzzaman, M.; Qiao, S.; Jin, W.-B.; Shi, H.; Li, X.V.; Iliev, I.D.; Artis, D.; et al. Microbiota metabolism of intestinal amino acids impacts host nutrient homeostasis and physiology. Cell Host Microbe 2024, 32, 661–675.e610. [Google Scholar] [CrossRef]
- Metges, C.C. Contribution of microbial amino acids to amino acid homeostasis of the host. J. Nutr. 2000, 130, 1857s–1864s. [Google Scholar] [CrossRef]
- Yang, C.-H.; Xu, J.-H.; Ren, Q.-C.; Duan, T.; Mo, F.; Zhang, W. Melatonin promotes secondary hair follicle development of early postnatal cashmere goat and improves cashmere quantity and quality by enhancing antioxidant capacity and suppressing apoptosis. J. Pineal. Res. 2019, 67, e12569. [Google Scholar] [CrossRef]
- Duan, T.; Wu, Z.; Zhang, H.; Liu, Y.; Li, Y.; Zhang, W. Effects of melatonin implantation on carcass characteristics, meat quality and tissue levels of melatonin and prolactin in Inner Mongolian cashmere goats. J. Anim. Sci. Biotechnol. 2019, 10, 70. [Google Scholar] [CrossRef]
- Tan, B.; Yin, Y.; Liu, Z.; Li, X.; Xu, H.; Kong, X.; Huang, R.; Tang, W.; Shinzato, I.; Smith, S.B.; et al. Dietary l-arginine supplementation increases muscle gain and reduces body fat mass in growing-finishing pigs. Amino Acids 2009, 37, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Xiao, Y.; Ma, L.; Lyu, W.; Peng, H.; Wang, X.; Ren, Y.; Li, J. Low dose of sucralose alter gut microbiome in mice. Front. Nutr. 2022, 9, 848392. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Han, D.; Su, Z.; He, L.; Zhang, W. Effect of melatonin on the production performance, blood biochemical parameters, nutrient digestibility, and gastrointestinal microbiome of Liaoning cashmere goats. Agriculture 2024, 14, 1983. [Google Scholar] [CrossRef]
- Zheng, Z.; Lyu, W.; Ren, Y.; Li, X.; Zhao, S.; Yang, H.; Xiao, Y. Allobaculum involves in the modulation of intestinal ANGPTLT4 expression in mice treated by high-fat diet. Front. Nutr. 2021, 8, 690138. [Google Scholar] [CrossRef]
- Duan, C.; Xu, J.; Sun, C.; Jia, Z.; Zhang, W. Effects of melatonin implantation on cashmere yield, fibre characteristics, duration of cashmere growth as well as growth and reproductive performance of Inner Mongolian cashmere goats. J. Anim. Sci. Biotechnol. 2015, 6, 22. [Google Scholar] [CrossRef]
- Zambrano, C.; Garitaonaindia, M.T.; Salmerón, D.; Pérez-Sanz, F.; Tchio, C.; Picinato, M.C.; de Medina, F.S.; Luján, J.; Scheer, F.A.J.L.; Saxena, R.; et al. Melatonin decreases human adipose tissue insulin sensitivity. J. Pineal. Res. 2024, 76, e12965. [Google Scholar] [CrossRef]
- Cano-Barquilla, P.; Jiménez-Ortega, V.; Fernández-Mateos, P.; Virto, L.; Maldonado Bautista, E.; Perez-Miguelsanz, J.; Esquifino, A.I. Daily lipolysis gene expression in male rat mesenteric adipose tissue: Obesity and melatonin effects. Int. J. Mol. Sci. 2025, 26, 577. [Google Scholar] [CrossRef]
- Deng, K.; Liu, Z.; Su, Y.; Fan, Y.; Zhang, Y.; Wang, F. Comparison of muscle fiber characteristics and meat quality between newborn and adult Haimen goats. Meat Sci. 2024, 207, 109361. [Google Scholar] [CrossRef]
- Stacchiotti, A.; Favero, G.; Rodella, L.F. Impact of melatonin on skeletal muscle and exercise. Cells 2020, 9, 288. [Google Scholar] [CrossRef]
- Reiter, R.J.; Rosales-Corral, S.; Tan, D.X.; Jou, M.J.; Galano, A.; Xu, B. Melatonin as a mitochondria-targeted antioxidant: One of evolution’s best ideas. Cell. Mol. Life Sci. 2017, 74, 3863–3881. [Google Scholar] [CrossRef]
- Park, J.; Sun, C.W.; Song, S.; Cheng, H.; Im, C.; Shin, T.S.; Kim, G.-D. Longitudinal and transversal intramuscular variation of muscle fiber and meat quality characteristics in bovine M. longissimus thoracis et lumborum. Meat Sci. 2024, 217, 109610. [Google Scholar] [CrossRef] [PubMed]
- Pastsart, U.; De Boever, M.; Claeys, E.; De Smet, S. Effect of muscle and post-mortem rate of pH and temperature fall on antioxidant enzyme activities in beef. Meat Sci. 2013, 93, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Sayed, R.K.A.; Fernández-Ortiz, M.; Diaz-Casado, M.E.; Aranda-Martínez, P.; Fernández-Martínez, J.; Guerra-Librero, A.; Escames, G.; López, L.C.; Alsaadawy, R.M.; Acuña-Castroviejo, D. Lack of NLRP3 inflammasome activation reduces age-dependent sarcopenia and mitochondrial dysfunction, favoring the prophylactic effect of melatonin. J. Gerontol. A 2019, 74, 1699–1708. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.-S.; Yang, Y.; Mou, L.; Xia, X.; Liu, M.; Xu, L.-J.; Liu, R.; Liu, J.-P.; Zhang, H.-Y.; Ao, X.-J.; et al. Melatonin ameliorates age-related sarcopenia via the gut–muscle axis mediated by serum lipopolysaccharide and metabolites. J. Cachexia Sarcopenia Muscle 2025, 16, e13722. [Google Scholar] [CrossRef]
- Ma, L.; Tao, S.; Song, T.; Lyu, W.; Li, Y.; Wang, W.; Shen, Q.; Ni, Y.; Zhu, J.; Zhao, J.; et al. Clostridium butyricum and carbohydrate active enzymes contribute to the reduced fat deposition in pigs. iMeta 2024, 3, e160. [Google Scholar] [CrossRef]
- Xiao, Y.; Huicong, Z.; Jingjing, L.; Tongxing, S.; Wentao, L.; Wen, W.; Zhenyu, W.; and Tao, S. Impact of quorum sensing signaling molecules in gram-negative bacteria on host cells: Current understanding and future perspectives. Gut Microbes 2022, 14, 2039048. [Google Scholar] [CrossRef]
- Wang, D.; Chen, L.; Tang, G.; Yu, J.; Chen, J.; Li, Z.; Cao, Y.; Lei, X.; Deng, L.; Wu, S.; et al. Multi-omics revealed the long-term effect of ruminal keystone bacteria and the microbial metabolome on lactation performance in adult dairy goats. Microbiome 2023, 11, 215. [Google Scholar] [CrossRef]
- Malmuthuge, N.; Liang, G.; Guan, L.L. Regulation of rumen development in neonatal ruminants through microbial metagenomes and host transcriptomes. Genome Biol. 2019, 20, 172. [Google Scholar] [CrossRef]
- Hai, C.; Hao, Z.; Bu, L.; Lei, J.; Liu, X.; Zhao, Y.; Bai, C.; Su, G.; Yang, L.; Li, G. Increased rumen Prevotella enhances BCAA synthesis, leading to synergistically increased skeletal muscle in myostatin-knockout cattle. Commun. Biol. 2024, 7, 1575. [Google Scholar] [CrossRef]
- Dou, L.; Liu, C.; Chen, X.; Yang, Z.; Hu, G.; Zhang, M.; Sun, L.; Su, L.; Zhao, L.; Jin, Y. Supplemental Clostridium butyricum modulates skeletal muscle development and meat quality by shaping the gut microbiota of lambs. Meat Sci. 2023, 204, 109235. [Google Scholar] [CrossRef]
- Zhang, P.; Jiang, G.; Wang, Y.; Yan, E.; He, L.; Guo, J.; Yin, J.; Zhang, X. Maternal consumption of l-malic acid enriched diets improves antioxidant capacity and glucose metabolism in offspring by regulating the gut microbiota. Redox Biol. 2023, 67, 102889. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.-C.; Kao, M.-R.; Saldivar, R.K.; Díaz-Moreno, S.M.; Xing, X.; Furlanetto, V.; Yayo, J.; Divne, C.; Vilaplana, F.; Abbott, D.W.; et al. The Gram-positive bacterium Romboutsia ilealis harbors a polysaccharide synthase that can produce (1,3;1,4)-β-d-glucans. Nat. Commun. 2023, 14, 4526. [Google Scholar] [CrossRef] [PubMed]
- Yao, T.; Wang, C.; Liang, L.; Xiang, X.; Zhou, H.; Zhou, W.; Hou, R.; Wang, T.; He, L.; Bin, S.; et al. Effects of fermented sweet potato residue on nutrient digestibility, meat quality, and intestinal microbes in broilers. Anim. Nutr. 2024, 17, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Tang, H.; Xie, Z.; Yi, H.; Feng, L.; Zhou, P.; Zhang, Y.; Liu, J.; Ao, X.; Zhou, J.; et al. Daily feeding frequency impacts muscle characteristics and fat deposition in finishing pigs associated with alterations in microbiota composition and bile acid profile. Front. Microbiol. 2025, 16, 1510354. [Google Scholar] [CrossRef]
- Wang, F.; Sha, Y.; Liu, X.; He, Y.; Hu, J.; Wang, J.; Li, S.; Shao, P.; Chen, X.; Yang, W.; et al. Study of the interactions between muscle fatty acid composition, meat quality-related genes and the ileum microbiota in Tibetan sheep at different ages. Foods 2024, 13, 679. [Google Scholar] [CrossRef]
- Ioannou, A.; Berkhout, M.D.; Geerlings, S.Y.; Belzer, C. Akkermansia muciniphila: Biology, microbial ecology, host interactions and therapeutic potential. Nat. Rev. Microbiol. 2025, 23, 162–177. [Google Scholar] [CrossRef]
- Cani, P.D.; Depommier, C.; Derrien, M.; Everard, A.; de Vos, W.M. Akkermansia muciniphila: Paradigm for next-generation beneficial microorganisms. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 625–637. [Google Scholar] [CrossRef]
- Lu, S.; Xu, Y.; Song, X.; Li, J.; Jiang, J.; Qin, C.; Wu, K.; Cui, K.; Liu, Y.; Liu, Q.; et al. Multi-omics reveal the effects and regulatory mechanism of dietary neutral detergent fiber supplementation on carcass characteristics, amino acid profiles, and meat quality of finishing pigs. Food Chem. 2024, 445, 138765. [Google Scholar] [CrossRef]
- Kang, L.; Li, X.; Zhao, X.; Liu, T.; Jin, Y.; Duan, Y. Effects of L-arginine supplementation on fat deposition and meat quality in growing lambs: Interactions with gut microbiota and metabolic signalling pathways. Food Chem. 2025, 479, 143677. [Google Scholar] [CrossRef]
- Taglialegna, A. Fat, Desulfovibrio and cancer. Nat. Rev. Microbiol. 2024, 22, 388. [Google Scholar] [CrossRef]
- Dong, Y.; Fanyi, M.; Jingyi, W.; Jingge, W.; Kexin, Z.; Siqi, Q.; Mengfan, L.; Fucheng, W.; Bangmao, W.; Tianyu, L.; et al. Desulfovibrio vulgaris flagellin exacerbates colorectal cancer through activating LRRC19/TRAF6/TAK1 pathway. Gut Microbes 2025, 17, 2446376. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | (%) | Nutrient Levels 2 | |
|---|---|---|---|
| Alfafa | 35.00 | ME, (MJ/Kg) | 9.69 |
| Peanut straw | 35.00 | DM, (%) | 86.99 |
| Corn | 16.00 | EE, (%) | 3.62 |
| Soybean meal | 9.50 | CP, (%) | 14.80 |
| Fermented soybean meal | 3.50 | Ca, (%) | 1.00 |
| Dicalcium phosphate | 0.10 | P, (%) | 0.32 |
| Salt | 0.50 | NDF, (%) | 24.66 |
| Premix 1 | 0.30 | ADF, (%) | 47.38 |
| Total | 100.00 | ASH, (%) | 6.70 |
| Item | C | M | p-Value |
|---|---|---|---|
| Slaughter weight (kg) | 28.73 ± 0.84 | 26.26 ± 0.65 | 0.071 |
| Carcass weight (kg) | 12.40 ± 0.48 | 11.27 ± 0.36 | 0.073 |
| Carcass yield (%) | 43.13 ± 0.73 | 43.05 ± 1.07 | 0.958 |
| Height at withers (cm) | 52.55 ± 0.96 | 50.01 ± 0.80 | 0.106 |
| Length of body (cm) | 56.30 ± 0.48 | 54.83 ± 0.95 | 0.245 |
| Circumference of chest (cm) | 8.98 ± 0.12 | 8.64 ± 0.15 | 0.163 |
| Head weight (kg) | 1.79 ± 0.04 | 1.74 ± 0.02 | 0.215 |
| Hoof weight (kg) | 0.70 ± 0.02 | 0.68 ± 0.02 | 0.158 |
| GR (cm) | 4.82 ± 0.08 | 4.18 ± 0.16 | 0.322 |
| Rib-eye area (cm2) | 20.41 ± 2.35 | 18.33 ± 0.98 | 0.182 |
| Perirenal fat (g) | 470.74 ± 43.56 a | 299.38 ± 49.70 b | 0.049 |
| Greater omental (g) | 946.86 ± 48.52 a | 569.20 ± 58.06 b | 0.002 |
| Mesentery fat (g) | 400.06 ± 41.47 a | 312.58 ± 23.07 b | 0.044 |
| Item | C | M | p-Value | |
|---|---|---|---|---|
| Weight (g) | Heart | 78.04 ± 4.16 | 68.94 ± 1.73 | 0.108 |
| Liver | 467.20 ± 39.23 | 428.38 ± 14.88 | 0.432 | |
| Spleen | 30.62 ± 0.94 | 28.84 ± 2.13 | 0.514 | |
| Lung | 265.74 ± 21.78 | 219.92 ± 13.77 | 0.972 | |
| Kidney | 82.62 ± 2.28 | 70.36 ± 2.31 | 0.100 | |
| Index (%) | Heart | 0.27 ± 0.01 | 0.29 ± 0.01 | 0.139 |
| Liver | 1.62 ± 0.10 | 1.81 ± 0.08 | 0.177 | |
| Spleen | 0.11 ± 0.01 | 0.12 ± 0.01 | 0.172 | |
| Lung | 0.93 ± 0.08 | 0.93 ± 0.07 | 0.972 | |
| Kidney | 0.29 ± 0.01 | 0.24 ± 0.01 | 0.422 | |
| Ovarian (g) | 1.56 ± 0.15 | 1.55 ± 0.14 | 0.997 | |
| Rumen weight (kg) | 3.60 ± 0.47 | 3.16 ± 0.31 | 0.499 | |
| Net rumen (kg) | 1.07 ± 0.12 | 1.05 ± 0.11 | 0.932 | |
| Rumen pH | 6.74 ± 0.26 | 6.79 ± 0.09 | 0.904 | |
| Item | C | M | p-Value | |
|---|---|---|---|---|
| LTL | pH45min | 6.12 ± 0.20 | 6.44 ± 0.23 | 0.076 |
| pH24h | 5.71 ± 0.23 b | 6.26 ± 0.36 a | 0.033 | |
| L* | 37.76 ± 1.66 | 38.07 ± 1.54 | 0.795 | |
| a* | 19.32 ± 1.33 | 17.82 ± 1.43 | 0.164 | |
| b* | 4.62 ± 0.44 a | 3.96 ± 0.34 b | 0.038 | |
| Drip loss (%) | 6.47 ± 2.79 | 2.98 ± 1.86 | 0.071 | |
| Cooking loss (%) | 50.54 ± 4.85 | 43.35 ± 6.68 | 0.129 | |
| Shear force (kgf) | 9.72 ± 3.34 b | 12.88 ± 2.00 a | 0.008 | |
| Moisture content (%) | 66.90 ± 2.43 | 71.54 ± 0.57 | 0.100 | |
| Crude protein content (%) | 55.20 ± 2.44 b | 61.93 ± 0.99 a | 0.034 | |
| Ether extract content (%) | 8.84 ± 1.37 | 7.80 ± 1.15 | 0.555 | |
| GL | pH45min | 6.39 ± 0.25 | 6.61 ± 0.08 | 0.133 |
| pH24h | 5.69 ± 0.29 | 6.01 ± 0.27 | 0.178 | |
| L* | 36.15 ± 1.30 | 36.97 ± 0.98 | 0.509 | |
| a* | 18.88 ± 0.91 | 17.80 ± 1.41 | 0.822 | |
| b* | 4.02 ± 0.44 | 4.05 ± 0.46 | 0.676 | |
| Drip loss (%) | 4.52 ± 2.10 | 3.10 ± 1.94 | 0.351 | |
| Cooking loss (%) | 53.25 ± 4.12 | 50.04 ± 6.97 | 0.451 | |
| Shear force (kgf) | 9.28 ± 4.35 | 12.59 ± 2.10 | 0.208 | |
| Moisture content (%) | 70.75 ± 0.42 | 71.17 ± 0.70 | 0.567 | |
| Crude protein content (%) | 59.65 ± 4.60 | 64.25 ± 2.65 | 0.127 | |
| Ether extract content (%) | 9.46 ± 0.48 | 9.94 ± 1.85 | 0.802 | |
| BF | pH45min | 6.34 ± 0.22 | 6.45 ± 0.14 | 0.339 |
| pH24h | 5.75 ± 0.10 | 6.17 ± 0.43 | 0.093 | |
| L* | 37.76 ± 1.66 | 38.07 ± 1.54 | 0.343 | |
| a* | 19.32 ± 1.33 | 17.82 ± 1.43 | 0.237 | |
| b* | 4.65 ± 0.44 | 3.96 ± 0.34 | 0.930 | |
| Drip loss (%) | 3.42 ± 1.18 | 2.99 ± 1.63 | 0.679 | |
| Cooking loss (%) | 49.61 ± 5.38 | 45.20 ± 8.07 | 0.390 | |
| Shear force (kgf) | 6.60 ± 3.45 | 9.90 ± 0.95 | 0.102 | |
| Moisture content (%) | 69.70 ± 2.62 b | 72.32 ± 0.58 a | 0.003 | |
| Crude protein content (%) | 63.77 ± 4.91 b | 68.68 ± 0.98 a | 0.002 | |
| Ether extract content (%) | 7.77 ± 0.77 | 6.99 ± 0.98 | 0.451 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, D.; Zheng, Z.; Su, Z.; Wang, X.; Ding, S.; Wang, C.; He, L.; Zhang, W. Melatonin Promotes Muscle Growth and Redirects Fat Deposition in Cashmere Goats via Gut Microbiota Modulation and Enhanced Antioxidant Capacity. Antioxidants 2025, 14, 645. https://doi.org/10.3390/antiox14060645
Han D, Zheng Z, Su Z, Wang X, Ding S, Wang C, He L, Zhang W. Melatonin Promotes Muscle Growth and Redirects Fat Deposition in Cashmere Goats via Gut Microbiota Modulation and Enhanced Antioxidant Capacity. Antioxidants. 2025; 14(6):645. https://doi.org/10.3390/antiox14060645
Chicago/Turabian StyleHan, Di, Zibin Zheng, Zhenyu Su, Xianliu Wang, Shiwei Ding, Chunyan Wang, Liwen He, and Wei Zhang. 2025. "Melatonin Promotes Muscle Growth and Redirects Fat Deposition in Cashmere Goats via Gut Microbiota Modulation and Enhanced Antioxidant Capacity" Antioxidants 14, no. 6: 645. https://doi.org/10.3390/antiox14060645
APA StyleHan, D., Zheng, Z., Su, Z., Wang, X., Ding, S., Wang, C., He, L., & Zhang, W. (2025). Melatonin Promotes Muscle Growth and Redirects Fat Deposition in Cashmere Goats via Gut Microbiota Modulation and Enhanced Antioxidant Capacity. Antioxidants, 14(6), 645. https://doi.org/10.3390/antiox14060645